











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
La mayoría de las cuencas de cabecera (1-3 orden) del mundo reciben desde los bosques aledaños, grandes cantidades de material foliar o leñoso que forman la materia orgánica particulada gruesa (MOPG). Este material constituye la principal fuente de energía para las redes tróficas en dichos ecosistemas ya que los procesos autotróficos son limitados por la poca incidencia de luz en el cauce (Hynes, 1975; Rosas et al., 2020; Vannote et al., 1980; Wallace et al., 1997). Sin embargo, el aporte y almacenamiento de MOPG se ve alterado por la deforestación asociada a sistemas agropecuarios (Allan, 2004; Chará et al., 2007).
La MOPG ingresa al afluente por dos vías principales: De manera vertical desde el dosel de los árboles que cubren la fuente de agua y de forma lateral arrastrada desde las zonas aledañas a los arroyos (Elosegi & Pozo, 2005). La primera vía está determinada principalmente por la fenología, calidad, abundancia y diversidad de la vegetación ribereña; la segunda, por la inclinación del terreno que determina la cantidad y periodicidad de entrada del material (Wantzen et al., 2008). Las entradas pueden ser continuas o no durante todo el año y los picos dependen del clima de la región; en algunos casos se dan en épocas de lluvia y en otra en épocas secas (Afoso et al., 2000; Colón-Gaud et al., 2008; Gonçalves et al., 2006).
Cuando la hojarasca ingresa al afluente, es transportada a lo largo del cauce por acción del agua y una parte empieza a ser transformada en las pozas o sitios que poseen estructuras retentivas como rocas grandes y acumulaciones de ramas y troncos, mientras que otra parte se exporta aguas abajo (Abelho, 2001; Tank et al., 2010). Por lo tanto, el transporte y retención de la materia MOPG depende de las características físicas propias (tamaño y densidad), de parámetros hidráulicos como la velocidad y turbulencia del flujo, de las características geomorfológicas de las quebradas tales como la geometría del canal, la presencia de rocas y madera, y de características biológicas como la presencia de plantas acuáticas (Allan et al., 2021; Hoover et al., 2010; Horvat, 2003). La retención del material por un periodo prolongado permite el procesamiento por parte de organismos detritívoros especializados, lo que contribuye a la construcción de las redes alimenticias del ecosistema lótico y al flujo de energía a través de estas (Jones, 1997; Lamberti & Gregory, 2006).
Adicionalmente, la magnitud y la dinámica de la MOPG que ingresa a las quebradas de cabecera depende del tipo e intensidad de las actividades humanas que se desarrollan en la cuenca y particularmente en la zona ribereña (Delong & Brusven, 1994; Quinn et al., 1997; Renöfält & Nilsson, 2008). Por ejemplo, la eliminación total o parcial de la vegetación ribereña por actividades ganaderas modifican la cantidad, la calidad y la fenología de las entradas de materia orgánica (Scarsbrook et al., 2001).
En Latinoamérica, la ganadería bovina ha sido una actividad histórica (> 500 años) y hoy día predomina en varias zonas típicamente agrícolas. En algunos países puede ocupar entre el 40 y 80 % del área total, lo que ha generado impactos importantes no sólo en el suelo sino también en los ambientes acuáticos (FAO, 2008; Murgueitio & Ibrahim, 2009). Un ejemplo de esto es la región del Eje Cafetero de Colombia, donde el cambio de usos del suelo ha deteriorado las cuencas de cabecera y la zona ribereña (Chará et al., 2007; Chará-Serna et al., 2015; Giraldo et al., 2014).
La eliminación de los bosques protectores de las zonas ribereñas es el detonante para los impactos negativos sobre la calidad del agua, los macroinvertebrados acuáticos, algunos aspectos físicos de los riachuelos, entre otros (Chará et al., 2007; Giraldo et al., 2014; Johnson & Host, 2010; Machado-Silva et al., 2022; Scanlon et al., 2007). Sin embargo, a pesar de la importancia ecológica y social de dichos ecosistemas en Colombia y el norte de Suramérica, es escasa la información sobre el efecto que la actividad ganadera ejerce en la dinámica de la MOPG.
Dado que la eliminación de los bosques ribereños y su reemplazo por pastizales puede alterar la dinámica de la MOPG, el objetivo de este estudio fue evaluar las diferencias en las entradas, el almacenamiento y la exportación de dicho material entre quebradas con bosques y quebradas con pastizales en su zona ribereña, ubicadas en un paisaje ganadero del Eje Cafetero de Colombia.
Materiales y métodos
Área de estudio: La región del Eje Cafetero se localiza en el centro occidente de Colombia en la región Andina, comprende los departamentos del Quindío, Risaralda, Caldas, Valle del Cauca (Norte) y Tolima (Noroccidente), y cubre un área de 28 563 km2. La región presenta un régimen bimodal de lluvias, con dos periodos de mayor precipitación al año (marzo-mayo, octubre-noviembre), la cual depende del gradiente altitudinal en el cual se encuentra. La precipitación promedio anual en la región es de 1 950 mm (Uribe-Gómez, 2009).
En la región predominan pasturas para la producción de ganadería bovina, buena parte de las cuales se establecieron en zonas donde previamente hubo cultivos de café (Coffea arabiga) (1 200 y 1 800 m.s.n.m). El cambio de uso de suelo de café a pasturas se dio principalmente en los años 90 debido a que los productores abandonaron su producción como consecuencia de la inestabilidad económica por los bajos precios internacionales (Sadeghian et al., 1999).
La presente investigación fue realizada en quebradas de cabecera de primer y segundo orden ubicadas en una zona de influencia ganadera del Eje Cafetero de Colombia, departamentos de Caldas (Municipios de Villamaría y Manizales), Quindío (Salento y Filandia) y Risaralda (Santa Rosa de Cabal) (Fig. 1). Se seleccionaron cinco microcuencas que poseen bosques nativos en la zona ribereña (denominadas Bosques) y en las cuales no se desarrollan actividades agropecuarias en un área aledaña aproximada de 20 m de ancho a cada lado del cauce. Para comparación, se escogieron cuatro quebradas desprotegidas cubiertas por gramíneas en su zona ribereña (denominadas Pastizal), las cuales son usadas para pastoreo de bovinos. Todos los sitios de estudio están localizados en alturas entre los 1 800 y 2 400 m.s.n.m.
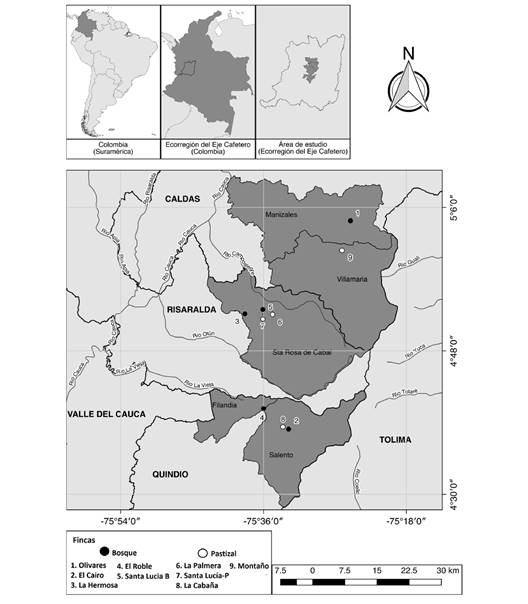
Fig. 1 Ubicación de nueve quebradas evaluadas en el Eje Cafetero, Colombia. / Fig. 1. Location of nine streams under study in the Colombian Coffee-growing Region.
La composición florística de los bosques ribereños de las microcuencas, pertenece a la zona de vida bosque húmedo montano bajo (bh-MB), según la clasificación de Holdridge (1947). La intervención antrópica en estos sitios se ha dado de manera selectiva relacionada con la tala de árboles de maderas apreciadas en la región como especies de la familia Lauraceae. Debido a la proximidad de la ganadería y pastoreo en la zona, se ha modificado la cobertura y estructura típica de la zona de vida a la que pertenece, evidenciándose la presencia de especies pioneras en áreas con regeneración natural como Heliocarpus americanus (Malvaceae), Cecropia telealba (Urticaceae) y Baccharis sp. (Asteraceae), entre otras.
En el caso de las quebradas clasificadas como pastizal, dos de los sitios estudiados presentan en la zona ribereña plantas de hábito herbáceo y arbustivo representado principalmente en familias como Melastomataceae y Fabaceae. Los sitios sin vegetación leñosa en la zona ribereña presentan gramíneas exóticas como pasto estrella (Cynodon nlemfuensis) y pasto Kikuyo (Cenchrus clandestinus).
Entradas de MOPG: Con base en la metodología propuesta por Pozo et al. (2009) y Rodríguez-Barrios et al. (2008), en cada una de las quebradas de estudio se seleccionó un tramo de 100 m en el cual se midieron los aportes verticales de hojarasca utilizando 10 canastas de caída de 0.5 x 0.5 m en el centro del cauce. Para los aportes laterales se instalaron 10 trampas de 0.5 x 0.5 m en la margen derecha e izquierda de la fuente de agua, dispuestas cada 10 m, para un total de 30 canastas por quebrada.
En las quebradas con pastizal en la zona ribereña sólo fueron instaladas canastas laterales, ya que en dicha zona los escasos arbustos no alcanzaban a tener un aporte vertical. Las canastas fueron construidas con marcos de PVC de ½ pulgada y mallas de Nylon con poro de 1 mm. La colecta de la hojarasca se realizó cada 15 días, durante el periodo comprendido entre febrero de 2016 a marzo de 2017.
MOPG acumulada en el lecho: Usando un cuadrante de 0.30 x 0.30 m, se tomaron 10 muestras de hojarasca aleatoriamente a lo largo del tramo de 100 m seleccionado para cada quebrada, lo cual permitió estimar la biomasa de la MOPG acumulada en el cauce.
Exportación de la MOPG: Cada dos meses, en cada quebrada, se dispusieron redes de deriva (al inicio del tramo que corresponde a 0 m, a los 50 m y al final del tramo a los 100 m), las cuales se ubicaron en cada sección cubriendo el cauce correspondiente para evitar pérdida del material. Las redes se mantuvieron por un periodo de 24 horas en cada muestreo. En la entrada de cada red, se midió la profundidad del agua y la velocidad de la corriente, datos que fueron usados para el cálculo de la densidad de la deriva de acuerdo con la siguiente ecuación (Rodríguez-Barrios, 2011):
D = (g)/(T.V.A)
donde: D= Densidad de deriva de MOPG (gramos de MOPG m-3); g= gramos de MOPG; T= Tiempo de exposición de la red (segundos); V= Velocidad de la corriente en la boca de la red (m.s-1); A= Área sumergida de la red (m2).
Para los tres casos (entradas, acumulación y exportación), las muestras recolectadas fueron almacenadas en el campo en bolsas plásticas debidamente rotuladas, luego se transportaron al laboratorio donde se secaron inicialmente a temperatura ambiente y posteriormente en un horno a 60 °C por un periodo de 48 horas, para determinar su peso seco mediante el uso de una balanza analítica con precisión de 0.0001 g. En el laboratorio se separó el material en tres categorías (hojas, material reproductivo, madera), para determinar los aportes de cada ítem.
Características físicas de los sitios de estudio: En el tramo de 100 m seleccionado en cada quebrada, se midió la cobertura del dosel cada 10 metros (donde se encontraban ubicadas las canastas de colecta de MOPG de caída vertical), mediante el uso de un densiómetro esférico cóncavo (Forest Densiometers Model-C). La pendiente del talud y del tramo se determinó usando una estación total Topcon GTS 235 en los sitios de pastizal y con un nivel de precisión Topcon AT-B 4 en los sitios con bosques. La velocidad de la corriente fue medida cada mes usando un velocímetro (VALEPORT 801, Flat), además se tomaron datos del ancho del cauce y profundidad de agua, que sirvieron para el cálculo del caudal.
Análisis de datos: Los datos de las entradas verticales y laterales, material acumulado, exportación de la MOPG y tipo de material que ingresó a las fuentes de agua fueron analizados inicialmente de manera descriptiva.
Posteriormente, las entradas totales, tipo de entrada (vertical, lateral), acumulado y exportación de la MOPG fueron comparados entre tipos de cobertura ribereña (bosque y pastizal) y meses de estudio usando un análisis de varianza de dos vías (ANOVA). Cuando fue necesario el ajuste a la normalidad, se efectuaron transformaciones log (x + 1).
Las características físicas de las quebradas se compararon entre tipos de cobertura de la ribera mediante una prueba t (Tabla 1). Los análisis fueron realizados con el programa estadístico R (R Core Team, 2018) versión 3.5.2.
Tabla 1 Características físicas de las quebradas de estudio en el Eje Cafetero de Colombia (Valores promedio, rangos). / Table 1. Physical characteristics of the streams studied in the Colombian Coffee- growing Region (Average values, ranges)
| Variable | Zona ribereña | ||
| Bosque n=5 | Pastizal n=4 | ||
| Cobertura del dosel (%) | 93.7 (80.7-98.7)a | 24.4 (0-42.2)b | |
| Pendiente del talud (%) | 85.3 (70-115.6)a | 47.6 (34.8-57.8)b | |
| Pendiente del tramo (%) | 11.2 (7.2-16.5) a | 4.8 (2.9-6.7)b | |
| Ancho del cauce (m) | 1.38 (1.03-1.85) | 0.89 (0.67-1.09) | |
| Profundidad (m) | 0.47 (0.06-1.15) | 0.11 (0.06-0.20) | |
| Caudal (L/s) | 20.2 (7.7-35.8) | 17.7 (5.8-28.3) | |
Letras distintas indican diferencias significativas. / Different letters indicated significant differences.
Resultados
Entradas de la MOPG: El ingreso promedio anual de la MOPG en las quebradas con bosques ribereños fue de 915 (± 183.7) g m-2, valor siete veces mayor que el observado en aquellas que presentan pastizales que fue de 125.4 (± 109.5) g m-2. En las quebradas con bosques, la entrada vertical promedio fue 400.3 (± 177) g m-2 y la lateral fue 514.6 (± 194.3) g m-2. Los ingresos de MOPG fueron significativamente diferentes entre los dos tipos de quebradas (bosques y pastizales) (ANOVA, F 1,5975 = 870.8, P < 0.0001) y entre el tipo de entrada (lateral y vertical) (ANOVA; F 1,5975 = 48.2, P < 0.0001). El ingreso diario promedio en las quebradas con bosques y pastizal en su zona ribereña fue de 3.5 (± 0.69) g m-2 d-1 y 0.47 (± 0.28) g m-² d-1, respectivamente (Fig. 2).
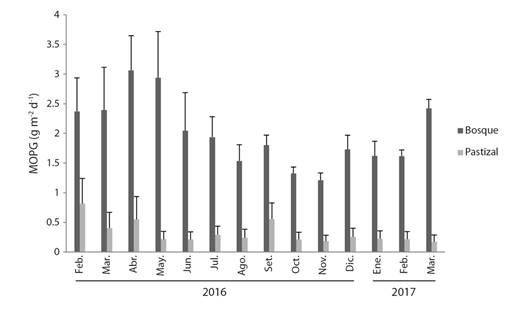
Fig. 2 Valores diarios de MOPG (promedio, EE) que ingresa a las quebradas con bosques y pastizales en la zona ribereña, en el Eje Cafetero, Colombia. / Fig. 2. Daily values of CPOM (mean, SE) that entered the streams with forests and grasslands in the riparian zone, in Colombian Coffee-growing Region.
Los meses con mayor ingreso diario promedio de MOPG fueron febrero, marzo, abril, mayo de 2016 y marzo de 2017, lo cuales fueron significativamente diferentes a los meses de octubre y noviembre (ANOVA; F 13,5963 = 10.2, P < 0.0001).
En cuanto al tipo de material que ingresó a las fuentes de agua, las hojas obtuvieron mayores valores en los dos tipos de cobertura evaluados con 2.4 g m-2 d-1 (bosques) y 0.33 g m-2 d-1 (pastizales), lo que representa el 76 y 92 % del total de las entradas (Fig. 3). La madera presentó el segundo valor más alto en quebradas con bosques (1 g m-2 d-1, 17 %), mientras que, en los pastizales, el material reproductivo y la madera tuvieron valores bajos (Fig. 3).
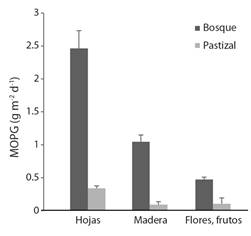
Fig. 3 Valores diarios (promedio, EE) de diferentes componentes de la MOPG que ingresa a las quebradas con bosques y pastizales en la zona ribereña en el Eje Cafetero, Colombia. / Fig. 3. Daily values (mean, SE) of different components of the CPOM that entered in streams with forests and grasslands in the riparian zone (Colombian Coffee-growing Region).
Materia MOPG acumulada: La MOPG acumulada en el lecho fue significativamente mayor en quebradas con presencia de bosques en la zona ribereña 732 (± 168.4) g m-2, que en quebradas con pastizales 205.5 (± 81.7) g m-2 (ANOVA; F 1,227 = 87.02, P < 0.0001). En junio se presentó un mayor valor de MOPG acumulada en el lecho en quebradas con bosques ribereños y en octubre en quebradas con pastizales (Fig. 4). Para las tres fechas, el material acumulado en las quebradas con bosque fue alrededor de tres veces mayor que en las quebradas con pastizal.
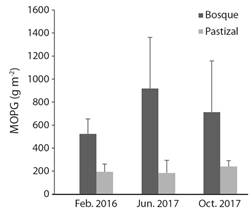
Fig. 4 Valores de MOPG acumulada en el lecho de quebradas con bosques y pastizales en su zona ribereña, en el Eje Cafetero, Colombia (promedio, E). / Fig. 4. Values of CPOM accumulated in the channel of streams with forests and grasslands in its riparian zone, located in the Coffee-growing Region of Colombia (mean, SE).
Exportación de la MOPG: La exportación total promedio de MOPG en los ocho periodos de muestreo fue de 334 g en las quebradas con bosques y 128 g en las quebradas con pastizal. La exportación promedio diaria en las quebradas con bosque y con pastizal fue de 1.27 (± 0.64) y 0.70 (± 0.27) g m-3 d-1 respectivamente, aunque estas diferencias no fueron significativas (ANOVA; F 1,451 = 2.68, P > 0.05). La cantidad de MOPG promedio diaria exportada en los meses de muestreo se presenta en la fig. 5. En las quebradas con bosques ribereños el mes con mayor exportación promedio fue noviembre de 2015 con 0.45 (± 0.081) g m-3 d-1 y el menor febrero de 2016 con 0.03 (± 0.02) g m-3 d-1. En cuanto a las quebradas con pastizales, noviembre de 2016 fue el mes con mayor exportación promedio 0.16 (± 0.09) g m-3 d-1 y febrero de 2016 el de menor cantidad 0.04 (± 0.02) g m-3 d-1.
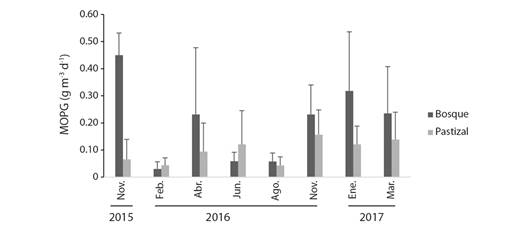
Fig. 5 Variación temporal de la exportación de MOPG (promedio, EE) en quebradas con bosques y pastizales en su zona ribereña, en el Eje Cafetero, Colombia. / Fig. 5. Temporal variation of CPOM export (mean, SE) in streams with forests and pastures in the riparian zone, located in the Coffee-growing Region of Colombia.
Discusión
Los resultados de este estudio apoyan la hipótesis de que la deforestación de la zona ribereña debido al manejo ganadero, tiene un efecto negativo sobre la transferencia y almacenamiento de la MOPG en quebradas de cabecera de la zona andina central de Colombia. La magnitud de los ingresos totales de MOPG desde los bosques ribereños fue significativa en el ámbito de los estudios disponibles. Estos valores coinciden con otras investigaciones realizadas en Colombia (Rodríguez-Barrios et al., 2008), pero difieren en otros casos de trabajos a nivel nacional y mundial; por ejemplo: 1 050 g m-2/año (Larned, 2000), 460-600 g m-2/año (Benfield et al., 2000), 1 134 g m-2/año (Afoso et al., 2000), 1 406-2 812 g m-2/año (Chará et al., 2005), 820 g m-2/año (Wantzen & Wagner, 2006), 1 137 g m-2/año (Colón-Gaud et al., 2008).
El mayor porcentaje de entrada de MOPG se dio por vía lateral con 56.2 % y el restante 43.7 % por vía vertical. Además, la mayor contribución relativa de material que ingresó a las fuentes de agua estuvo dado por hojas (76.7 %) y en menor proporción por flores y frutos (6.8 %). Patrones similares se han registrado en otros estudios, como los realizados por Benson & Pearson (2020), Chará et al. (2005), Colón-Gaud et al. (2008), Iroumé et al. (2020), Molinero (2019), Rodríguez-Barrios (2011) y Tonin et al. (2017). Cabe resaltar, que en varios de estos estudios no se ha incluido la contribución de madera de pequeñas ramas, lo cual constituye un elemento de importancia en la formación de estructuras físicas en el cauce, al crear barreras que permiten la retención de hojarasca foliar, creación de pozas y otro tipo de microhábitats ocupados para los organismos acuáticos (Bisson et al., 2017; Diez et al., 2000; Morris et al., 2007; Tank et al., 1993).
Aunque los aportes totales (lateral + vertical) de MOPG desde la vegetación aledaña se produjeron durante todo el año, hubo mayor ingreso en épocas secas en la región (febrero, marzo, abril). Esto es consistente con lo encontrado por Colón-Gaud et al. (2008), Wantzen et al. (2008), Wright & Cornejo (1990) and Zalamea & González (2008). Mientras en zonas templadas los picos de entrada de hojarasca se dan en otoño mediados por la disminución de la temperatura del aire, en el Trópico los mayores aportes se dan en época seca, básicamente por las bajas precipitaciones y mayor radiación solar lo que genera estrés hídrico, además de la fenología de la vegetación. Sin embargo, esto puede depender del bioma evaluado, que determina el tipo de vegetación en las regiones (Tonin et al., 2017).
Existen pocos estudios sobre el aporte de MOPG en sitios deforestados, lo cual es llamativo considerando la presión sobre los bosques de la zona ribereña en todo el mundo. Algunos estudios han encontrado ingresos desde 2.8 a 264.8 g m-2/año en quebradas con alta e intermedia actividad agropecuaria y entre 68 y 276.4 g m-2/año de aporte de pastos en zonas de actividad ganadera (Hagen et al., 2010; Menninger & Palmer, 2007). Los ingresos en sitios impactados se relacionan mayormente con la presencia de parches de árboles que se encuentran en la zona ribereña tal como sucedió con el presente estudio, donde a pesar de la simplificación de la composición de la vegetación adyacente a las quebradas inmersas en pastizales, en algunos casos presentaron vegetación secundaria conformada por melastomatáceas y piperáceas. La distribución dispersa de árboles (“parches” o grupos), además de pastos y hierbas, tiene un efecto significativo sobre el ingreso MOPG a los sistemas lóticos (Larned, 2000; Menninger & Palmer, 2007). Sin embargo, es necesario nuevos estudios específicos del tema, que permita determinar cómo influye esta condición en la dinámica de la MOPG.
La reducción de las entradas verticales y laterales de MOPG desde la zona ribereña tiene implicaciones importantes en el funcionamiento del ecosistema lótico, especialmente para la comunidad de macroinvertebrados que usan dicho recurso como alimentación y refugio (Kobayashi & Kagaya, 2004). La deforestación, sin duda, ejerce presión negativa sobre la cantidad y calidad de hojarasca que ingresa a los canales e induce a cambios en los procesos energéticos del sistema (Allan et al., 2021; Sweeney et al., 2004).
En este estudio la magnitud promedio de la MOPG acumulada en el lecho, fue cerca de cuatro veces superior en las quebradas con bosques que con pastizales (Fig. 6). Sin embargo, en términos generales, las quebradas mostraron valores de almacenamiento menores que los reportado en otros estudios similares (Mathuriau et al., 2008; Rodríguez-Barrios, 2011). La retención de las partículas que ingresan a las fuentes de agua tiene que ver con la cantidad de sustratos rocosos gruesos como bloques y cantos rodados presentes, pero además con el tamaño de la partícula y los restos de madera (Cordova et al., 2008; Miller, 2013).
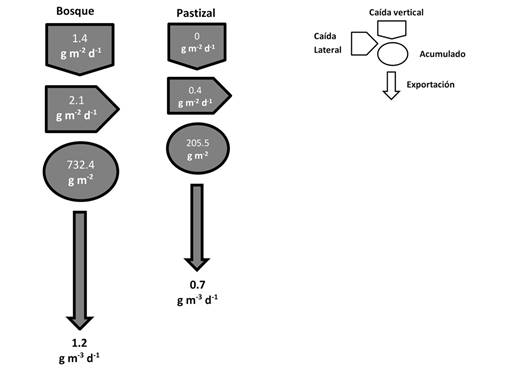
Fig. 6 Modelo conceptual de balance de masas simplificado de la materia orgánica particulada gruesa en microcuencas de zonas ganaderas del Eje Cafetero, Colombia. / Fig. 6. Simplified conceptual model of mass balance of coarse particulate organic matter in micro-basins of cattle areas (Coffee-growing Region of Colombia).
Además de la disminución de la cantidad de MOPG que ingresa a las fuentes de agua, la eliminación de los bosques de la zona ribereña, reduce el porcentaje de la sombra en el canal e incrementan la temperatura del agua (Bourque & Pomeroy, 2001; Hagen et al., 2010), como se ha visto en este estudio donde los sitios sin bosques en la zona ribereña presentaron en promedio 21 °C de temperatura, mientras en el grupo de sitios protegidos fue de 18 °C. Por otro lado, la deforestación de la vegetación leñosa al lado de los cauces genera erosión de los bancos y con esto, el incremento de sedimentos finos en el cauce (Bilby & Bisson, 1998; Herbst et al., 2012).
En general, las microcuencas estudiadas tienen una naturaleza exportadora de la MOPG, los valores para esta variable están cercanos a las entradas (Fig. 6). La MOPG puede entrar a los riachuelos en grandes cantidades dentro de periodos cortos de tiempo y esporádicamente en eventos atmosféricos extremos (Liu et al., 2018). Además, las variaciones temporales y espaciales están asociadas con los patrones de lluvia, el flujo del agua y con el nivel de madurez de la vegetación en la cuenca (Heartsill-Scalley et al., 2012). En el caso particular de las quebradas en estudio, los altos pulsos de entrada de MOPG a las quebradas, pueden estar dados por la fenología de los árboles y la pendiente del terreno, que permite un aporte durante todo el año del material a nivel de la cuenca y por ende una exportación constante a los ríos. Además, las quebradas como las estudiadas experimentan eventos torrenciales que permiten el arrastre del material en periodo de minutos a pocas horas.
Se esperaba que la disminución de la exportación de MOPG en arroyos con pastizales en la zona ribera, fuese proporcional a la disminución de las entradas. Sin embargo, a pesar de que las entradas diarias en las quebradas con pastizal equivalen aproximadamente al 10 % de lo que entra a las quebradas con bosques ribereños (principalmente por la ruta lateral), la exportación es mayor. Esto puede tener relación con las pocas estructuras retentivas que posee este tipo de ambientes (menor cantidad piedras, acumulaciones de ramas y troncos, etc.). Además, durante algunos periodos de creciente las quebradas con pasto exportaron más material, posiblemente por una menor retención y liberación de reservorios relativamente inactivos. En otros momentos de alto caudal exportaron menos posiblemente por el agotamiento de dichos reservorios. En general, la condición exportadora de las cuencas en estudio tiene implicaciones importantes para ríos de mayor tamaño por la cantidad de carbono que se moviliza a través del transporte del material (Bunte et al., 2016; Cuevas-Lara et al., 2021; Turowski et al., 2016).
De acuerdo con el presente estudio, la destrucción de los bosques ribereños en esta región genera una reducción de aportes de MOPG de 7 896 kg/ha/año. Esto es importante por la gran extensión que alcanzan las zonas pecuarias dominadas por pastos limpios en la zona Andina de Colombia que equivalen a 10 915 099 ha, segundo lugar en extensión en el país, después de la Orinoquía (IAVH, 2018). De manera inversa, este estudio permite suponer cuál sería el aporte de MOPG si los actuales pastizales que comprometen la zona ribereña entran en un proceso de sucesión secundaria o restauración ecológica. Según Sánchez-Cuervo et al. (2012), entre el 2000 y 2010, hubo una tendencia de recuperación de bosques en la zona Andina de Colombia, lo cual brinda la oportunidad de expandir las áreas protegidas actuales y promover la conectividad del hábitat.
Aunque es necesario seguir explorando sobre los efectos que tienen las actividades agropecuarias en ecosistemas acuáticos de cabecera, los resultados de este estudio permiten comprender la influencia de dichas actividades sobre el balance de masas de la MOPG (Fig. 6) y de esta forma diseñar el manejo y la gestión de las cuencas en paisajes tropicales ganaderos de Sur América.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

