











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Entre los insectos, los formícidos se destacan por las interacciones que establecen con las semillas de diferentes plantas. Entre ellas se incluye la dispersión o mirmecocoria, que es el movimiento de las semillas lejos de la planta madre para asegurar su germinación y supervivencia (Delabie et al., 2003), la remoción que consiste en la relocalización de las semillas (Detrain & Tasse, 2000) y la depredación o granivoría que implica el consumo de las semillas y por lo tanto su destrucción (MacMahon et al., 2000).
Algunas de las semillas que son removidas presentan eleosomas que son cuerpos ricos en lípidos que están adheridos a las diásporas y que atraen a las hormigas (Christianini et al., 2007), sin embargo, muchas de las que son removidas no presentan estas estructuras (Escobar-Ramírez et al., 2012). Una vez removidas, las semillas pueden ser consumidas o almacenadas en los nidos (Jansen et al., 2004), parte de las que son almacenadas se acumulan en cámaras especiales dentro de los nidos o se descartan en sitios específicos, como las pilas de desecho denominadas basureros donde una proporción puede germinar (Retana et al., 2004). La tasa de germinación puede aumentar debido a la escarificación mecánica o química producida por las hormigas al manipularlas (Escobar-Ramírez et al., 2012; Hurtado et al., 2012; Jansen et al., 2004; Ohkawara & Akino, 2005; Salazar-Rojas et al., 2012).
De acuerdo con la teoría del forrajeo óptimo, el tiempo y la energía que cada consumidor invierte en la recolección de alimento son cruciales para maximizar las ganancias energéticas (Pyke, 1980), por esta razón las hormigas utilizan diversas características de las semillas para obtener las mayores ganancias con el menor gasto de energía. En este sentido, la selectividad depende de características que cambian en el tiempo y el espacio, como son la distribución y abundancia de las semillas (Wilby & Shachak, 2000). De esta forma, se ha propuesto que las hormigas serían más selectivas cuando la disponibilidad de sus semillas preferidas es mayor y menos selectivas cuando decrece la disponibilidad de éstas (Crist & MacMahon, 1992; Wilby & Shachak, 2000). También interfieren en la selectividad de las semillas características anatómicas como su forma, tamaño, presencia de apéndices, peso y textura (Escobar-Ramírez et al., 2012; Pulliam & Brand, 1975). Las características nutricionales también resultan ser un factor decisivo ya que se ha encontrado que las hormigas prefieren semillas con un alto contenido nutricional (Kelrick et al., 1986), aunque también pueden remover aquellas con bajo contenido y que pueden ser consumidas en caso de presentarse escasez de alimento (Tschinkel & Kpawich, 2016).
En regiones con estacionalidad marcada, la distribución espacial y temporal de los recursos varía, lo que afecta la actividad de forrajeo de las hormigas y la selectividad de semillas (Pol et al., 2011; Wilby & Shachak, 2000). En algunas regiones se ha encontrado que hay mayor actividad de forrajeo durante la época de lluvia, ya que hay mayor disponibilidad de recursos que pueden ser explotados por varias especies de hormigas (Coelho & Ribeiro, 2006; Marsh, 1985).
En México se sabe que la remoción de semillas se lleva a cabo por hormigas de varios géneros como Forelius, Crematogaster, Dorymyrmex Ectatomma, Pheidole, Monomorium, Solenopsis, Tapinoma, Trachymyrmex y Pogonomyrmex (Cuautle et al., 2005; Munguía-Rosas et al., 2009; Ríos-Casanova et al., 2004), aunque muchos estudios se han enfocado en hormigas estrictamente granívoras como Pogonomyrmex barbatus (Luna et al., 2018; Ríos-Casanova, 2005). Sin embargo, varias especies omnívoras también pueden remover semillas y eventualmente incluirlas en su dieta (Vergara-Torres et al., 2018). Por ejemplo Azteca gnava que remueve semillas de orquídeas (Morales-Linares et al., 2018) o Dorymyrmex goetschi que remueve semillas de cactáceas (Escobar-Ramírez et al., 2012).
La manipulación de semillas por hormigas puede tener un papel clave en la germinación. La escarificación con las mandíbulas y la eliminación del eleosoma por hormigas omnívoras de la familia Dolichoderinae entre las que se encuentran Forelius analis y Dorymyrmex thoracicus, afectan positivamente la germinación de las semillas que remueven (Cuautle et al., 2005; Leal et al., 2007). Además, la aplicación de secreciones de la glándula metapleural puede disminuir la probabilidad de infecciones por patógenos y aumenta la posibilidad de germinación (Ohkawara & Akino, 2005).
El género Dorymyrmex Mayr es uno de los más diversos de la subfamilia Dolichoderinae (Cuezzo & Guerrero, 2012). Aunque se reconoce como un género de hormigas omnívoras que remueve ítems muy diversos, algunas especies de Sudamérica como D. antarcticus, D. insanus, D. minutus, D. tener y D. wolffhuegeli, participan activamente en la remoción de la semilla exótica Carduus nutans (Asteraceae), mientras que D. brunneus y D. thoracicus remueven semillas de diferentes especies de la familia Euphorbiaceae (Leal et al., 2007; Pirk & López de Casenave, 2017), sin embargo en México no se conoce qué semillas podrían estar removiendo las hormigas de este género.
Dentro de la Ciudad de México (CDMX), se encuentra ubicada la Reserva Ecológica del Pedregal de San Ángel (REPSA). La REPSA se reconoce como un ecosistema embebido dentro de una de las ciudades más grandes del mundo y se caracteriza por ser una zona muy heterogénea y compleja (Lot & Cano-Santana, 2009). La REPSA tiene dos temporadas bien diferenciadas, la de lluvias que abarca de junio a octubre, y la seca que comprende de noviembre a mayo (Castillo-Argüero et al., 2004). Se ha visto que la biomasa vegetal y producción de frutos y semillas disponibles en el sitio varía dependiendo de la estación ya que ésta es mayor durante la temporada de lluvias (Cano-Santana, 1994). Como resultado de los estudios que se han realizado en la REPSA para conocer su dinámica y diversidad (Lot & Cano-Santana, 2009), se ha encontrado que en ella habitan al menos 37 especies de hormigas de seis subfamilias entre las que se incluyen granívoras, depredadoras y omnívoras (Ríos-Casanova et al., 2017), sin embargo, aún no se conocen las interacciones de éstos insectos con las plantas ni se tienen datos sobre la remoción de semillas u otros ítems, ni sobre los efectos que la remoción puede tener sobre la germinación.
Dorymyrmex insanus (Hymenoptera: Formicidae) es una especie omnívora que puede remover semillas, restos vegetales y artrópodos (Hung, 1974). Esta hormiga se ha registrado en la zona Núcleo Poniente de la REPSA y es una de las especies más abundantes del sitio, principalmente en la temporada lluviosa, aunque también está activa durante la temporada seca (Jiménez-Vargas, 2017). A pesar de que se tienen antecedentes previos de la remoción de semillas por D. insanus en otros sitios (Carney et al., 2003), no existe información de cómo son los basureros de sus nidos, ni acerca de los ítems que remueve y deposita en éstos o cuál es el porcentaje de germinación de las semillas removidas por esta hormiga en la REPSA.
En este trabajo se evaluó el papel de D. insanus en la remoción de semillas y otros ítems dentro de la REPSA durante la temporada seca y de lluvias, a partir del análisis de los basureros asociados a sus nidos. Por la estacionalidad tan acentuada de la REPSA, esperábamos encontrar una mayor diversidad de semillas removidas por esta especie en la temporada de lluvias, ya que es cuando se encuentran disponibles los propágulos de muchas especies. Asimismo, se esperaba un efecto positivo de D. insanus sobre la germinación de la especie de planta cuyas semillas fueron las más abundantes en los basureros de sus nidos (Tagetes micrantha, ver Resultados), ya que éstas han sido manipuladas por las hormigas, en contraste con las semillas no manipuladas.
Materiales y métodos
Sitio de estudio: Este trabajo se realizó entre diciembre de 2018 y febrero de 2020 en la REPSA, ubicada al suroeste de la Ciudad de México (19°31’N - 99°19’W) (De la Fuente, 2005). El clima es templado subhúmedo (García, 1988), la temperatura media anual es de 15.5 °C y la precipitación promedio anual de 835 mm (Castillo-Argüero et al., 2004). La vegetación predominante es matorral xerófilo con una gran diversidad florística. Se caracteriza por tener pocas especies arbóreas y un gran número de herbáceas donde las asteráceas son las mejor representadas (Castillo-Argüero et al., 2004; Rzedowski, 1954). Los nidos de D. insanus se encuentran principalmente en los senderos de la zona Núcleo Poniente y tienen una distancia de 30 a 50 m entre sí. Los senderos son sitios planos con insolación y evaporación altas donde predominan especies como el pasto Muhlenbergia robusta, la cactácea Opuntia tomentosa y otros arbustos y herbáceas entre las que se encuentran las asteráceas Dahlia coccinea y Tagetes micrantha (Villanueva, 2015).
Remoción de ítems: Una vez que los nidos fueron localizados, se midieron, el diámetro de orificio de entrada, diámetro de la circunferencia que rodea al nido (disco) y altura del montículo, con un vernier digital (Steren HER-411). El basurero, que rodea la entrada del nido, fue dividido en cuatro ejes correspondientes a los puntos cardinales (N, S, E, O); en cada uno de estos puntos se tomó una muestra de material utilizando como medida una cuchara cafetera de 5 ml (Andersen et al., 2000). Estas muestras se recolectaron mensualmente en diez nidos durante 12 meses (dos días por mes de muestreo).
Ya que los basureros de D. insanus están constituidos por tierra, piedras y otros ítems orgánicos, se utilizó la técnica propuesta por Malone (1967), que permite la separación de la materia orgánica por flotación. Para evitar perdida de información por aquellos ítems que podrían no flotar, las muestras se revisaron previamente bajo microscopio estereoscópico. Los ítems encontrados se contaron y fueron asignados a las categorías: a) semillas, b) insectos y c) restos vegetales (raíces, ramas, hojas, partes florales). Se utilizó el término “semilla”, para referirnos a la unidad reproductiva completa más pequeña de una planta removida por las hormigas, aunque en ocasiones se trató de infrutescencias, frutos como los aquenios de las asteráceas o las cariópsides de las poáceas. Todas se determinaron a nivel de familia, género y cuando fue posible, a nivel de especie.
Para conocer si el tamaño de las semillas está relacionado con su abundancia en los basureros, se midieron el largo y el ancho mayor de 20 de cada una de las especies encontradas, cuando se contó con un número menor, se midió el total disponible. Las semillas se midieron a partir de fotografías tomadas con un microscopio estereoscópico marca Motic (modelo SMZ168 con cámara digital M5) y con ayuda del programa Motic Images Plus 3.0.
Pruebas de germinación: Se realizaron pruebas de germinación durante 40 días para examinar los posibles efectos de la remoción por hormigas sobre las semillas de T. micrantha, detectada como la especie más abundante y frecuente en los nidos de D. insanus (ver Resultados). Se recolectaron semillas directamente de los frutos disponibles en las plantas, las cuales fueron almacenadas en bolsas de papel glassine y en condiciones de obscuridad. También se recolectaron semillas de T. micrantha encontradas en los basureros de los nidos de D. insanus. Las muestras se recolectaron de la misma manera que cuando se analizaron los basureros, aunque una vez en el laboratorio, las semillas se separaron manualmente con el uso de un microscopio estereoscópico. En ambos tratamientos, se seleccionaron semillas de tamaño similar las cuales se presionaron con unas pinzas, aquellas que fueron intactas y duras se consideraron viables y se usaron en el experimento, aquellas dañadas y vacías se consideraron inviables y se descartaron.
Las pruebas de germinación se realizaron a temperatura ambiente y luz natural. Para cada tratamiento (semillas manipuladas por hormigas y semillas provenientes de plantas en pie) se utilizaron 20 cajas de Petri, con un diámetro de 60 mm y altura de 15 mm, con una gasa de algodón como sustrato. En esta gasa se colocaron 10 semillas para un total de 200 por tratamiento. La gasa de algodón fue humedecida con 5 ml de agua corriente. Para evitar la pérdida de humedad en las cajas, se añadieron 2 ml de agua a diario. Las cajas se revisaron diariamente para registrar el número de semillas que germinaron, lo cual se consideró con la emergencia de la radícula.
Análisis de datos: Se realizó un listado de las semillas presentes en los basureros de los nidos de D. insanus, así como el mes y la temporada en que fueron registradas. Para determinar si existían diferencias en la abundancia de las semillas entre temporadas, se hizo una prueba U de Mann-Whitney. La diversidad en ambas temporadas se evaluó utilizando el índice de diversidad de Shannon obtenido a partir de la rarefacción de los datos para lo cual se utilizó el programa EstimateS 9.0 y para comparar los valores obtenidos entre temporadas se obtuvieron los intervalos de confianza al 95 % (Colwell, 1997).
Para conocer la relación entre el ancho o largo de las semillas y su abundancia en los nidos se hicieron modelos de regresión para encontrar el mejor ajuste. El mejor modelo fue elegido con base en el valor del criterio Akaike (AIC). Los datos de abundancia de los ítems obtenidos fueron transformados utilizando el logaritmo natural de (x +1), con la finalidad de normalizar los datos. Para conocer el efecto de la manipulación ejercida por las hormigas a las semillas, se realizó una prueba de ANOVA de un factor al porcentaje final de semillas germinadas provenientes de cada tratamiento. Este porcentaje fue transformado previamente mediante la función arcoseno (Hurtado et al., 2012). Para estos análisis se utilizó el programa PAST 3.0 (Hammer et al., 2001).
Resultados
Caracterización de basureros: Las entradas a los nidos se ubican en el centro de los basureros y presentan una forma circular de diámetro 1.3 ± 0.11 mm (media ± E.E.). Estas entradas, se encuentran rodeadas por piedras que forman una estructura cónica donde las hormigas acumulan las semillas y otros ítems formando el basurero. El diámetro del disco de los basureros fue de 37.05 ± 2.49 mm (media ± E.E.), mientras que la altura fue de 7.81 ± 0.61 mm (media ± E.E.).
Remoción de ítems: En los basureros de D. insanus se encontraron semillas de 19 especies (ninguna de las cuales presenta eleosomas), restos de insectos de cuatro órdenes y restos de hojas, raíces y ramas (restos vegetales) (Tabla 1). Las semillas representaron el 50.55 % de los ítems removidos seguido por los restos vegetales (49.09 %) e insectos (0.35 %). Entre los insectos se encontraron restos de organismos de los órdenes Coleoptera, Hemiptera, Hymenoptera y Orthoptera. La mayor cantidad de restos de Hymenoptera corresponden a cabezas de la hormiga Solenopsis geminata.
Tabla 1 Semillas por familia y otros ítems presentes en los basureros de los nidos de Dorymyrmex insanus durante dos temporadas en la REPSA / Table 1. Seeds by botanical family and other items found in Dorymyrmex insanus nest middens during two seasons at REPSA.
| 2018 | 2019 | 2020 | ||||||||||
| Dic | Mar | Abr | Jun | Jul | Ago | Sep | Oct | Nov | Dic | Ene | Feb | |
| Semillas | Temporada seca | Temporada de lluvias | Temporada seca | |||||||||
| Asteraceae | ||||||||||||
| Bidens aurea | 3.8 | 0.4 | 0.4 | 1.5 | - | - | - | 1.1 | 2 | 0.7 | 0.6 | 2.2 |
| Dyssodia papposa | - | - | - | - | - | - | - | 4.7 | 0.3 | 11.5 | 0.9 | 0.6 |
| Heterosperma pinnatum | 0.5 | 0.8 | - | 0.2 | 0.5 | 0.3 | - | - | - | 0.7 | - | 0.6 |
| Tagetes micrantha | 82 | 90.1 | 74.1 | 25.3 | 9.1 | 9.5 | 3.6 | 48.3 | 59 | 49.5 | 35.9 | 478 |
| Convolvulaceae | ||||||||||||
| Ipomoea purpurea | - | - | - | 0.5 | 0.2 | 0.9 | 0.3 | - | - | - | - | - |
| Crassulaceae | ||||||||||||
| Echeveria gibbiflora | - | - | 0.4 | - | - | - | - | - | - | - | - | - |
| Cyperaceae | ||||||||||||
| Bulbostylis capillaris | - | - | 0.4 | - | 0.3 | - | 0.7 | - | - | - | - | - |
| Fabaceae | ||||||||||||
| Crotalaria pumila | 1 | - | - | 3.7 | 1.2 | 0.1 | 1.8 | 0.8 | 0.5 | 7.5 | 4.9 | 16.1 |
| Onagraceae | ||||||||||||
| Onagraceae sp. 1 | 9.9 | 0.4 | 6.1 | 4.8 | 0.3 | 0.5 | - | 0.8 | 11.2 | 1 | 2.3 | 1.1 |
| Oxalidaceae | ||||||||||||
| Oxalis corniculata | - | - | 0.4 | - | 0.2 | 0.1 | - | - | - | - | - | - |
| Poaceae | ||||||||||||
| Panicum sp. | - | - | - | 0.2 | - | - | - | - | - | - | - | - |
| Paspalum sp. | - | - | - | 0.3 | - | - | - | - | - | - | - | - |
| Poaceae sp.1 | 1 | - | 0.4 | - | - | - | - | - | - | - | - | - |
| Poaceae sp. 2 | 1.9 | 7.1 | 1.2 | - | - | - | - | - | - | - | - | - |
| Poaceae sp. 3 | - | - | - | 0.2 | - | - | - | - | - | - | - | - |
| Portulacaceae | ||||||||||||
| Portulaca pilosa | - | - | - | - | - | 2.4 | - | - | - | - | - | - |
| Solanaceae | ||||||||||||
| Solanum sp. | - | - | - | 0.2 | - | - | - | - | - | - | - | - |
| Verbenaceae | ||||||||||||
| Verbena carolina | - | 1.2 | 16.2 | 3.4 | 8.9 | 8.3 | 11 | 11 | 15 | - | - | - |
| No identificada | ||||||||||||
| Sp. 1 | - | - | 0.4 | - | - | - | - | - | - | - | - | - |
| Otros ítems | ||||||||||||
| Coleoptera | - | - | - | - | 0.2 | 0.2 | 0.3 | - | - | - | - | - |
| Hemiptera | - | - | - | - | 0.2 | - | - | - | - | - | - | |
| Hymenoptera | - | - | - | 0.6 | 0.7 | - | - | - | 0.2 | 0.4 | - | 1.1 |
| Orthoptera | - | - | - | - | - | 0.1 | - | - | - | - | - | 30.6 |
| Restos vegetales | - | - | - | 59.2 | 78.1 | 77.5 | 82.4 | 33.1 | 11.8 | 28.7 | 55.5 | - |
Valores expresados como porcentaje del total. / Values expressed as percentage of the total.
La familia Poaceae presentó el mayor número de especies (5), seguido por Asteraceae (4) (Tabla 1). El número de semillas removidas en la temporada seca fue significativamente mayor en comparación con las removidas en la temporada de lluvias (Z = -2.078, P = 0.037), mientras que el número de restos vegetales fue mayor en la temporada de lluvias (Z = -3.743, P < 0.001), en tanto que no hubo diferencia entre la abundancia de insectos presentes en ambas temporadas (Z = -1.805, P = 0.071). Las semillas de Tagetes micrantha fueron las más abundantes ya que representaron el 67.28 % del total de semillas registradas, seguido por Verbena carolina con 15.04 % (Fig. 1).
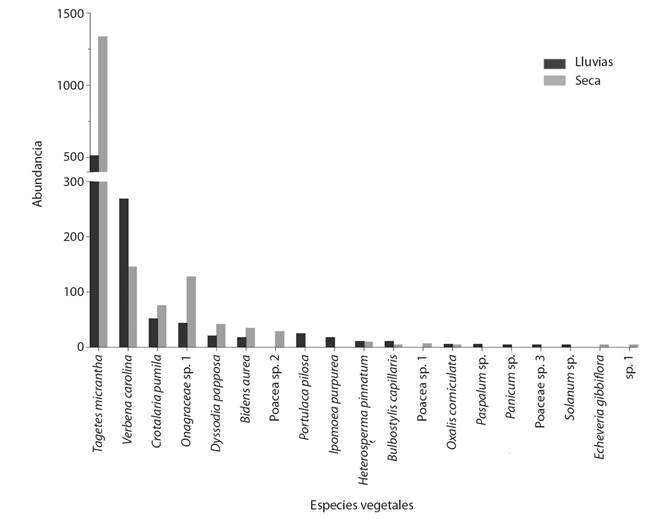
Fig. 1 Abundancia total de las semillas encontradas en los basureros de los nidos de Dorymyrmex insanus en la REPSA. / Fig. 1. Total abundance of seeds found in Dorymyrmex insanus nest middens at REPSA.
La diversidad de especies removidas fue mayor en la temporada de lluvias (H’ = 1.38 ± 0.031, índice de diversidad ± intervalo de confianza al 95 %) que en la temporada seca (H’ = 0.99 ± 0.025). Los índices son diferentes ya que sus intervalos de confianza no se traslapan.
Con respecto al tamaño de las semillas halladas en los basureros de D. insanus, la relación entre la longitud del largo mayor y la abundancia fue descrita con una regresión polinomial de tercer grado dado que presentó el mejor ajuste de AIC (39.91) la cual fue significativa (R² = 0.42, F = 3.51, P = 0.04, Fig. 2). El ancho de las semillas y su abundancia en los nidos no tuvieron ninguna relación (R² = 0.06, F = 1.095, P = 0.90).
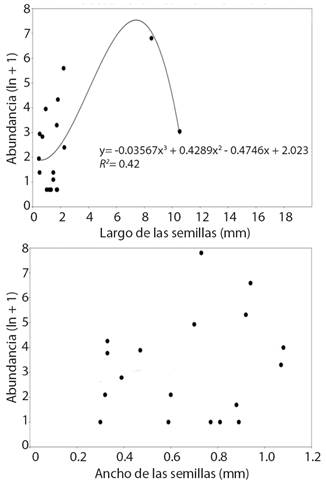
Fig. 2 Relación entre el largo o ancho de las semillas y su abundancia en basureros de los nidos de Dorymyrmex insanus en la REPSA. / Fig. 2. Relationship between the abundance and the length or width of the seeds found in the nest middens of Dorymyrmex insanus at REPSA.
Germinación de semillas: La proporción de semillas de T. micrantha germinadas fue mayor para las manipuladas por hormigas (0.47 ± 0.039) (media ± E.E.) que para aquellas provenientes de las plantas (0.27 ± 0.031, F = 16.16, g.l.= 39, P = 0.0002, Fig. 3).
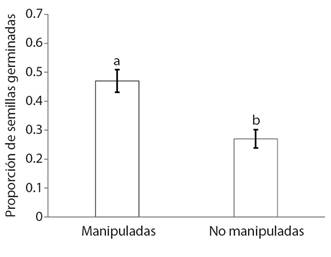
Fig. 3 Proporción de semillas de Tagetes micrantha germinadas manipuladas y no manipuladas por Dorymyrmex insanus. Medias (± E.E.) con letras distintas son significativamente diferentes (P < 0.05). / Fig. 3. Proportion of germinated Tagetes micrantha seeds manipulated and not manipulated by Dorymyrmex insanus. Means (± S.E.) with different letters are significantly different (P < 0.05).
Discusión
En este estudio se registró una gran variedad de ítems en los basureros de D. insanus, entre los que se encuentran: insectos, restos vegetales y semillas de 19 especies de plantas. La diversidad de semillas en estos basureros fue afectada por la temporalidad, pues, aunque en la temporada de lluvias se obtuvo la mayor diversidad, en la temporada seca los basureros tuvieron una abundancia mayor de semillas. Además, se encontró que la manipulación por D. insanus afecta positivamente la germinación de Tagetes micrantha.
En la REPSA se ha documentado una riqueza taxonómica de hasta 377 plantas vasculares (Castillo-Argüero et al., 2009), que son dispersadas principalmente por viento (César-García, 2002), sin embargo, las semillas de estas plantas pueden ser removidas por las hormigas luego de haber caído al suelo. Muchas especies de plantas presentes en la REPSA florecen y fructifican durante la temporada de lluvias (Castillo-Argüero et al., 2009), lo que puede estar explicando la mayor diversidad de semillas en los nidos de D. insanus durante esa época. Sin embargo, muchas especies de pastos y asteráceas producen sus propágulos al final de la temporada de lluvias por lo que sus semillas están disponibles al terminar esta temporada. Además, las plantas de estas especies producen una gran abundancia de semillas por lo que hay una gran cantidad disponible para las hormigas en la temporada seca (César-García, 2002). Esta gran disponibilidad es lo que puede explicar por qué se registró la mayor abundancia de semillas en los nidos de D. insanus, durante la temporada seca.
Estos hallazgos estarían de acuerdo con las predicciones de la teoría de forrajeo óptimo pues durante las lluvias habría poca disponibilidad de las semillas preferidas por las hormigas por lo que la dieta de éstas se ampliaría y por ello encontramos una diversidad más alta. Por el contrario, en la temporada seca hay una gran abundancia de semillas de asteráceas y pastos que son preferidas por estas hormigas y, por lo tanto, aunque la diversidad de semillas encontrada en los basureros en esta época es menor, hay una abundancia muy alta de algunas especies como por ejemplo T. micrantha. Sin embargo, para poder afirmar que D. insanus cumple con estas predicciones, es necesario que, de las semillas encontradas en los basureros de D. insanus, se cuantifique su disponibilidad y patrón de distribución en la REPSA porque esto puede estar afectando la forma en la que las hormigas las seleccionan (Pol et al., 2011; Wilby & Shachak, 2000).
Aunque D. insanus se considera una especie omnívora, en este estudio se encontró que más del 50 % de los ítems removidos por esta hormiga son semillas, por lo que podría considerarse que ejerce una función similar a la de muchas especies granívoras. Para otras especies como Dorymyrmex tener y Dorymyrmex antarcticus ya se ha reportado que son importantes removedoras de semillas en la Patagonia argentina (Ortiz et al., 2021). Debido a que el género Dorymyrmex suele ser considerado un grupo omnívoro y generalista, existen pocos estudios en los que se ha analizado su interacción con semillas, no obstante, los datos disponibles sugieren que este género podría tener un papel relevante en la remoción, por ello consideramos fundamental que se realicen estudios que incluyan experimentos donde se controle el número de semillas ofrecidas a esta especie de hormiga y a otras que habitan en la REPSA.
En este trabajo encontramos una gran abundancia de semillas de T. micrantha en los basureros de D. insanus, lo cual sugiere que esta hormiga remueve activamente las semillas de esta especie. De todas las que lleva a sus nidos, esta especie es la que presenta uno de los tamaños más grandes (8.51 ± 0.2 mm). De acuerdo con Pirk (2007) ante la oferta de semillas de diferentes tamaños, las más grandes suelen ser preferidas por las hormigas ya que ofrecen mayores beneficios en términos de biomasa y nutrientes, lo que podría explicar en parte los resultados obtenidos. Además, aunque encontramos que las semillas más grandes removidas por D. insanus son las de Bidens aurea (10.53 ± 0.52 mm), éstas son escasas en los basureros por lo que es muy probable que D. insanus tenga un límite en el tamaño de las semillas que remueve.
Otras especies de Dorymyrmex como D. brunneus son capaces de remover semillas de Cnidoscolus quercifolius (Euphorbiaceae) que tienen una longitud promedio de 13.5 mm (Leal et al., 2007), la cual es mayor a la longitud promedio de T. micrantha (8.51 mm), a pesar de que los tamaños de ambas hormigas son similares (Cuezzo & Guerrero, 2012). Un factor importante en la selectividad de semillas y su posterior remoción es su disponibilidad (Wilby & Shachak, 2000). Es probable que en la REPSA haya semillas más grandes que potencialmente podrían ser utilizadas por D. insanus sin embargo, debido a su baja disponibilidad, no fueron abundantes ni frecuentes como otras más pequeñas. La disponibilidad de semillas para D. insanus en la REPSA es un aspecto que tendrá que ser explorado en estudios futuros.
Las semillas de las gramíneas presentan bajas concentraciones de metabolitos secundarios en comparación con las de otras familias de plantas (Díaz, 1996). Esto podría explicar el número de especies de gramíneas removidas por D. insanus. Además, la calidad nutricional podría ser un factor que tuviera un efecto sobre la selectividad. La concentración de carbohidratos y ácidos grasos está asociada positivamente con la selectividad de las hormigas (Boulay et al., 2006; Kelrick et al., 1986). Por ejemplo, las semillas de Tagetes minuta contienen ácidos grasos como palmítico, linoleico y linolénico (Rezaei et al., 2018). Aunque en este estudio no se evaluaron características químicas de los componentes de las semillas de T. micrantha es posible que presente alguno de estos ácidos grasos lo cual podría explicar su alta frecuencia en los nidos de D. insanus, por lo que es necesario analizar los compuestos químicos de esta especie.
Varias especies del género Dorymyrmex remueven ítems que no consumen (e. g. ramas, hojas, partes florales y piedras) los cuales son colocados alrededor de la entrada del nido. La función de estos objetos ha sido asociada con marcadores que funcionan para el reconocimiento de nidos (Lenoir et al., 2009) y con la regulación de la temperatura (Smith & Tschinkel, 2007). No obstante, se desconoce qué otras funciones podrían tener la remoción de estos ítems no comestibles y si este comportamiento podría tener algún papel adaptativo (Cuautle et al., 2005).
En la REPSA habitan otras hormigas que podrían estar compitiendo con D. insanus por recursos o espacios para anidar, como por ejemplo Monomorium minimum, Nylanderia bruessi, Pheidole sp., y Solenopsis geminata que son especies dominantes en la zona Núcleo Poniente de la REPSA (Jiménez-Vargas, 2017). Debido a que no está clara cuál es la interacción que existe entre las hormigas presentes en el sitio de estudio, se requiere de trabajos en los que se analicen las interacciones entre estas especies.
En los nidos de D. insanus encontramos restos de Solenopsis geminata, lo cual coincide con los hallazgos de otros autores que han identificado a D. insanus como una fuerte competidora de S. geminata y Solenopsis invicta principalmente por sitios de anidación y forrajeo (Nickerson et al., 1975). Las observaciones indican que las obreras de D. insanus llevan los cuerpos a sus nidos, donde son desmembrados y las cabezas son colocadas en los basureros (Hung, 1974; Nickerson et al., 1975).
En este estudio, se encontró experimentalmente que las semillas de T. micrantha manipuladas por D. insanus presentan mayor capacidad germinativa comparadas con aquéllas que provenían directamente de las plantas en pie. Ese resultado sugiere que dicha manipulación podría mejorar las capacidades germinativas de las semillas debido a que probablemente las escarifica mediante procesos físicos, aunque esto requiere ser corroborado. No obstante, este manejo de las semillas por hormigas puede influir en la germinación a través de factores mecánicos y químicos (Leal et al., 2007; Ohkawara & Akino, 2005). En otros estudios se encontró que las semillas de Turnera ulmifolia (Passifloraceae) que fueron recolectadas en los nidos de la hormiga Forelius analis presentaron hasta cuatro veces mayor germinación que las que no provenían de los nidos (Cuautle et al., 2005; Salazar-Rojas et al., 2012). Otras especies como Odontomachus sp., Pachycondyla sp. y S. geminata pueden generar incisiones con las mandíbulas en semillas de Calathea microcephala (Marantaceae) durante los eventos de remoción favoreciendo su germinación (Horvitz, 1981). La escarificación permite que se rompa la latencia además de facilitar la entrada de agua y oxígeno, lo que aumenta la tasa de germinación (Traveset & Verdú, 2002).
Los resultados obtenidos sugieren que Dorymyrmex insanus juega un papel importante en la remoción de semillas en la REPSA. Los basureros de D. insanus reflejan de forma parcial el número de ítems que pueden remover estas hormigas. Aunque las observaciones en campo nos permitieron detectar a algunas obreras removiendo ítems como piedras pequeñas y ramas, además de algunas semillas, otros ítems podrían estar presentes en los basureros debido a factores como el azar o agentes de dispersión mecánica como el viento. Por ello consideramos indispensable continuar analizando la remoción que lleva a cabo esta hormiga, por ejemplo, retirando directamente los ítems que estas hormigas transportan.
La remoción realizada por D. insanus puede tener el potencial de aumentar la germinación de semillas, particularmente en el caso de T. micrantha. Con la finalidad de comprender los patrones en el destino de las semillas luego de ser removidas y descartadas se requieren de estudios a largo plazo en los que se compare la germinación o el reclutamiento de plantas en condiciones de campo. Además, se requiere de estudios adicionales para obtener información sobre el contenido nutricional y la composición química de las semillas removidas. Asimismo, aunque se analizaron todos los nidos disponibles de D. insanus se recomienda realizar búsquedas más intensivas que permitan complementar la información obtenida. Finalmente, los datos sobre la remoción de ítems por D. insanus representan una base sobre la cual, se podrán realizar investigaciones futuras sobre su comportamiento, ecología y estrategias de forrajeo.
Ethical statement: the authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.

