











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Los clipeastéridos (Echinoidea: Clypeasteroida) son erizos irregulares caracterizados por patrones corporales aplanados, siendo las galletas de mar el grupo con origen más reciente en su pasado geológico durante el Eoceno (55 M.a.) (Ghiold & Hoffman, 1986; Smith & Kroh, 2013). El género ClypeasterLamarck, 1801 está compuesto por 109 especies (Kroh & Mooi, 2020). Algunas de las características morfológicas que estableció Durham (1955) para definir a las especies del género son: presencia de una testa de tamaños medianos a grandes (80 a 200 mm de diámetro) en la cual, la altura máxima puede estar en posición generalmente central (Clypeaster europacificusClark, 1914) o ser completamente aplanada [Clypeaster humilis (Leske, 1778)]; la superficie ventral puede ser cóncava [Clypeaster rosaceus (Linnaeus, 1758)] o plana (C. europacificus); el margen de la testa puede ser redondeado, aplanado o grueso y tanto el sistema apical como el peristoma se localizan en una posición central de la testa.
Los clipeastéridos son organismos ampliamente distribuidos en los mares tropicales (Durham, 1955). En el litoral mexicano se tienen reportadas 30 especies pertenecientes al orden Clypeasteroida L. Agassiz, 1835. La familia Clypeasteridae L. Agassiz, 1835 está representada por 10 especies todas incluidas dentro del género Clypeaster, de las cuales sólo cuatro se distribuyen en el Pacífico mexicano: C. europacificusClark, 1914; Clypeaster ochrusClark, 1914; Clypeaster rotundus (A. Agassiz, 1863); y Clypeaster subdepressus (Gray, 1825) (Solís-Marín, Caballero-Ochoa, Frontana-Uribe, Laguarda-Figueras & Durán-González, 2017).
Clypeaster europacificus es una especie de galleta de mar que se ha reportado en rangos batimétricos de 0 a 402 m (Clark, 1914; Ziesenhenne, 1937; Maluf, 1988; Solís-Marín et al., 2013), sin embargo, en los registros correspondientes a la franja litoral del Pacífico mexicano, no existen registros de esta galleta de mar que superen los 200 m de profundidad (Conejeros-Vargas, Solís-Marín, & Laguarda-Figueras, 2017). Geográficamente C. europacificus tiene un rango latitudinal amplio a lo largo del Pacífico americano, siendo reportada desde Cabo San Lucas, México, hasta Perú (Maluf, 1988; Clark, 1914; Brusca, 1980; Solís-Marín et al., 2013). El Golfo de California es una cuenca marginal situada al noreste del Pacífico mexicano, se encuentra delimitado al occidente por la Península de Baja California y al oriente por las costas de los estados de Sonora y Sinaloa; se ubica entre los 23° y 32° de latitud Norte y entre los 107° y 115° de longitud Oeste. Esta cuenca se considera un mar semicerrado con poco más de 1 200 km de longitud y una anchura promedio de 150 km (Alvarez-Arellano & Molina-Cruz, 1986).
A pesar de que existen diversos trabajos que han tratado de identificar la relación de la morfología de la testa en erizos de mar con procesos de mineralización en las primeras etapas del desarrollo (Killian & Wilt, 2008; Martino, Chiarelli, Roccheri, Matranga, & Byrne, 2019) o bien durante las modificaciones que presenta la testa como resultado del crecimiento; el cual, puede evidenciarse ya sea al incrementar el tamaño o número de placas o bien, con la aparición de estructuras como lúnulas en las regiones interambulacrales en algunas galletas de mar fenómeno que modifica la distribución y tamaño de las placas (Seilacher, 1979; Swisher & Lin, 2019), aún son pocos los trabajos que describen la respuesta de los organismos durante el crecimiento y sus implicaciones al momento de identificar organismos.
El endoesqueleto o testa de las galletas de mar está conformado por la fusión de las placas ambulacrales e interambulacrales. Durante el crecimiento se ha identificado que el número de placas puede variar en algunas especies; en ejemplares de Echinarachnius parma (Lamarck, 1816) se ha reportado que el número de placas varia de 38 (en organismos recién metamorfoseados) a más de mil, considerando las placas dorsales, ventrales y de las áreas ambulacrales e interambulacrales en organismos adultos (Zachos, 2015). Particularmente, para C. europacificus no existen estudios que analicen la variación morfológica a lo largo de su crecimiento. Tener conocimiento de estas variaciones podría ayudar a seleccionar mejor los caracteres de importancia taxonómica e identificar a partir de qué talla dichos caracteres toman relevancia para la identificación específica en clipeastéridos y en la sistemática de este grupo.
En el presente trabajo se describen morfológicamente y se añaden comentarios relacionados con cambios y variaciones evidentes en la testa de C. europacificus; así como, una caracterización de dichos cambios mediante un análisis morfométrico, con el objetivo de conocer aspectos de su crecimiento relativo en estructuras de importancia taxonómica como la testa y algunas otras estructuras de importancia taxonómica.
Materiales y métodos
Se examinaron las características morfológicas de la testa de 1 857 especímenes identificados como Clypeaster europacificus que fueron recolectados dentro de los límites correspondientes al Golfo de California, México (Fig. 1) y que se encuentran resguardados en 48 lotes (1 846 especímenes) en la Colección Nacional de Equinodermos (CNE) “Dra. María Elena Caso Muñoz”, ICML-UNAM. Adicionalemente se examinó el material de la serie tipo depositado en el National Museum of Natural History Smithsonian Institution, Washington, D.C., U.S.A. (NMNH) (5 lotes con 7 especímenes) y en el Museum of Comparative Zoology, Universidad de Harvard, U.S.A. (MCZ) (2 lotes con 4 especímenes) (Anexo 1).
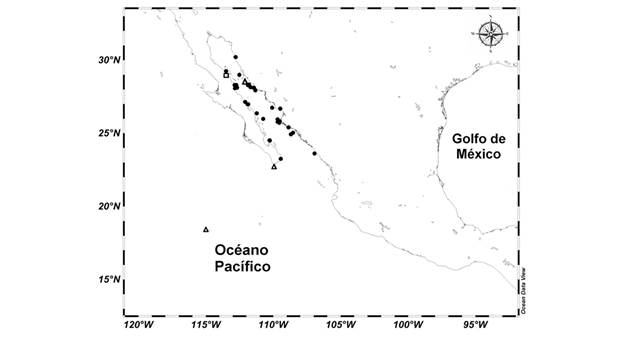
Fig. 1. Fig. 1 Registros del material examinado de C. europacificus. El cuadrado señala la localización geográfica de los especímenes utilizados en el análisis de crecimiento relativo [ICML-UNAM 4.27.33] y los triángulos el material tipo con localidad en México. Records of the material examined from C. europacificus. The square indicates the geographic location of the specimens used in the relative growth analysis [ICML-UNAM 4.27.33] and the triangles the type material with locality in Mexico.
Antes de iniciar los análisis, se corroboró la identidad taxonómica de los especímenes comparando los caracteres morfológicos externos de la testa con ayuda de un microscopio estereoscópico Olympus SZX7 en el Laboratorio de Sistemática y Ecología de Equinodermos (LSEE), ICML-UNAM y comparándolos con las descripciones contenidas en los trabajos de Clark (1914); Clark (1940; Clark (1948), Grant & Hertlein (1938), Mortensen (1948) (como Clypeaster [Alexandria] europacificus) y Caso (1961); Caso (1980).
Análisis morfológico. Los principales caracteres incluidos en el análisis descriptivo fueron: tamaño de los ejemplares, el cual fue estimado por el diámetro de la testa (D) medido desde el borde de la testa sobre el radio IV hasta el borde de la testa en el radio II sobre la superficie dorsal, longitud, cobertura y forma de los petaloides, forma del cuerpo (posición de la altura máxima de la testa), ubicación del peristoma y periprocto.
Para el análisis morfológico de microestructuras, se extrajeron espinas primarias, espinas secundarias y pedicelarios de la superficie dorsal y ventral de los organismos. El término microestructura(s) será empleado a partir de este momento en el texto para referirnos a estructuras de carbonato de calcio que están articuladas sobre la testa como espinas primarias, espinas secundarias y pedicelarios, los cuales pueden tener dimensiones superiores a 1 mm. Para eliminar el resto de tejido orgánico de las microestructuras, estos fueron sumergidos en una solución de hipoclorito de sodio [NaClO 5-6.5 %] sobre un portaobjetos excavado. Después de verificar que el tejido orgánico se había disuelto y que las microestructuras se habían precipitado sobre el portaobjetos, la solución de NaClO fue extraída con una pipeta y las microestructuras fueron enjuagadas con agua destilada. A continuación, se extrajo el excedente de agua destilada y se realizaron una serie de enjuagues con etanol a concentración creciente (70, 80 y 95 %) posteriormente, se añadió etanol absoluto y se dejó secar la muestra. Por último, las microestructuras fueron trasladadas a una cinta de carbono conductora de doble recubrimiento sobre una base metálica. Las muestras se recubrieron con oro 2,5 kV en el ionizador Polaron E3000 durante 3 minutos y posteriormente se fotografiaron usando un microscopio electrónico de barrido (MEB y SEM por sus siglas in inglés) Hitachi S-2460N, en el Laboratorio de Microscopía y Fotografía de la Biodiversidad II del Instituto de Biología (IB), UNAM.
Análisis de crecimiento relativo. A pesar de que algunos autores consideran que para comprender el crecimiento en organismos como los erizos de mar se debe identificar el crecimiento primero en las placas individuales de la testa, Zachos (2015) menciona que existen dos métodos para poder identificar el modelo de crecimiento individual en las placas de la testa. La primera es utilizar las bandas de crecimiento o bien comparar placas homólogas a través de una serie ontogenética de individuos donde se registre el crecimiento. Para C. europacificus se utilizó el criterio de homología de las placas de la superficie dorsal, considerando que durante el crecimiento las placas de la testa que están en la parte central se conservan y que las placas que se agregan quedan en los bordes de la testa, las proporciones entre las distancias lineales de cualquier estructura morfológica y el borde de la testa debe reflejar un cambio producto del crecimiento.
Dentro del material incluido en este trabajo se seleccionó un lote (ICML-UNAM 4.27.33) con especímenes que tuvieran la testa completa y sin marcas de regeneración, se analizaron 32 especímenes en un rango de tamaño de 1.2 a 10.1 cm de radio mayor (Fig. 4) y con un peso seco de 0.37 a 73.80 gr con material procedente de la bahía Santa Inés, Baja California Sur, México que fueron recolectados a 64.2 m de profundidad (Fig. 1).
A cada individuo seleccionado se le tomaron seis medidas: radio mayor (R), radio menor (r), altura de la testa (h), la longitud del petaloide anterior impar (LP), área de la testa (A) y el peso seco (P). Para obtener los valores de las distancias lineales (R, r, h, LP y A) sobre la testa, los ejemplares fueron fotografiados con una cámara digital Sony Alpha 57 y procesados con el software ImageJ escalando las fotografías (106 pixeles = 1 cm). Las medidas tomadas con este programa fueron las siguientes: R, desde el centro del sistema petalar al borde mayor de la testa sobre sobre la fusión de las placas ambulacrales del ambulacro II; r, desde el centro del sistema petalar hasta el borde posterior de la testa sobre la fusión de las placas interambulacrales 5 (Fig. 2A); LP, desde el borde de la madreporita hasta la última placa ambulacral con poros en el petaloide anterior impar (Fig. 2A, 2B); h, desde el borde de la membrana del peristoma en la superficie ventral hasta el punto central del sistema apical en la superficie dorsal; el área (A) se obtuvieron delimitando el contorno de la testa y utilizando las funciones correspondientes del programa. El peso seco (P) se midió con una balanza analítica VE-210; los valores de las mediciones se realizaron en el LSEE, ICML, UNAM y se procesaron en el programa Microsoft Excel.
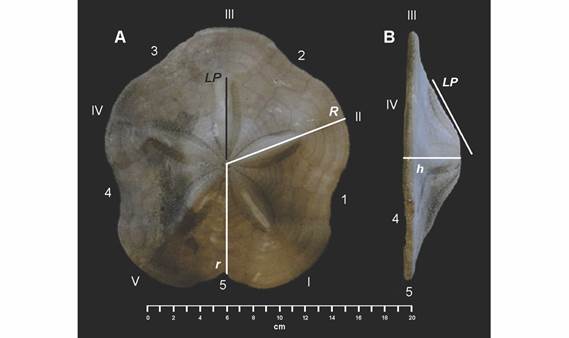
Fig. 2 Fig. 2 Clypeaster europacificus (ICML-UNAM 4.27.33). A. Vista dorsal (R = 10 cm). B. Vista lateral señalando las medidas tomadas: longitud del petaloide anterior impar (LP); radio mayor (R); radio menor (r) y altura (h). Áreas ambulacrales en números romanos (I-V); áreas interambulacrales en números arábigos (1-5). Clypeaster europacificus (ICML-UNAM 4.27.33). A. Dorsal view (R = 10 cm). B. Side view indicating the measurements taken: length of the odd anterior petaloid (LP); major radius (R); minor radius (r) and height (h). Ambulacral areas in Roman numerals (I-V); interambulacral areas in Arabic numerals (1-5).
Las relaciones morfométricas se determinaron tomando R como la variable independiente (debido a que consideramos que esta distancia refleja mejor la talla de los organismos, los cuales en etapas tempranas de su desarrollo tienen una forma casi circular [Fig. 3D y 3E]) y las variables restantes (r, LP, h, A y P) como dependientes. Las variables se ajustaron a la ecuación logarítmica de esta función, log Y = b (log X) + log a la cual, muestra la ecuación de una línea recta donde la pendiente b representa el parámetro de crecimiento (Huxley y Teissier, 1936). En las relaciones entre dos medidas lineales (R vs. r, R vs. LP y R vs. h), el valor de b se comparó contra el valor 1. Si b es mayor que 1 (alometría positiva), indica un aumento mayor de Y en relación con X; si b es menor que 1 (alometría negativa), implica un menor crecimiento de Y en relación con X; si b = 1 (isometría) ambas variables crecen al mismo ritmo (Gould, 1966; Huxley, 1924; 1932). Para verificar si b difería o no significativamente de 1, se aplicó la prueba
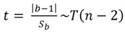 ,
,en donde
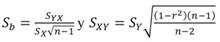 ,
,con S * igual a la desviación estándar de X o Y, r 2 el coeficiente de determinación y n el número de datos (Zar 1999). De manera análoga, en las relaciones con el área y peso (R vs. A y R vs. P), el valor de b se comparó a valores de 2 y 3 respectivamente, para determinar el tipo de crecimiento. Todos los análisis se hicieron a un nivel de significancia de α = 0.05.
Resultados
Sistemática
Orden Clypeasteroida Agassiz, 1835
Familia Clypeasteridae Agassiz, 1835
Género ClypeasterLamarck, 1801
Clypeaster europacificusClark, 1914
Clypeaster europacificusClark, 1914: 27-29; Clark, 1940: 349; Clark, 1948: 294-295; Grant & Hertlein, 1938: 45-46; Caso, 1961: 306; Caso, 1980: 5-10.
Clypeaster (Alexandria) europacificusMortensen, 1948: 53-54.
Los especímenes examinados poseen un diámetro de 2.4 a 18.4 cm, cuerpo comprimido dorsoventralmente. La región central de la testa correspondiente al sistema apical es convexa ya que la elevación puede ser de pocos milímetros (en especímenes con radio (R) ≤ 3 cm; Fig. 3D, 3E) a un par de centímetros (especímenes con R ≥ 3.1 cm; Fig. 3A, B, C). Dependiendo del tamaño de los ejemplares el borde de la testa puede ser circular (R ≤ 3 cm; Fig. 3D, E; Fig. 4A) o presentan prolongaciones de la testa en las áreas correspondientes a las placas ambulacrales dando un aspecto pentagonal al perímetro del cuerpo (Fig. 3A, B, C; Fig. 4B). Generalmente, en los ejemplares preservados en seco, todo el cuerpo presenta una coloración verdosa olivo o ligeramente café, sin importar el tamaño o la localidad. El Sistema apical es comprimido, las placas genitales están fusionadas en una sola placa genital central con forma de estrella con perforaciones irregularmente dispuestas; los poros genitales están dispuestos en las puntas de la estrella, las cuales corresponden con cada una de las cinco áreas interambulacrales.

Fig. 3 Fig. 3 Clypeaster europacificus (ICML-UNAM 4.27.33). Especímenes de diferentes tallas: A, B y C = R ≤ 3 cm. D y E = R ≥ 3.1 cm. 1. Vista dorsal. 2. Vista ventral. 3. Vista lateral. Clypeaster europacificus (ICML-UNAM 4.27.33). Specimens of different sizes: A, B and C = R ≤ 3 cm. D and E = R ≥ 3.1 cm. 1. Dorsal view. 2. Ventral view. 3. Lateral view.

Fig. 4 Fig. 4 Vista dorsal de especímenes de C. europacificus (ICML-UNAM 4.27.33). A. Especímenes con borde circular (R ≤ 3 cm). B. Especímenes con borde pentagonal (R ≥ 3.1 cm). Dorsal view of specimens of C. europacificus (ICML-UNAM 4.27.33). A. Specimens with circular edge (R ≤ 3 cm). B. Specimens with pentagonal edge (R ≥ 3.1 cm).
Placas oculares pequeñas, reducidas sólo a una pequeña placa con el borde cubierto por tubérculos miliares muy pequeños y una perforación ocular grande en el centro; cada placa ocular corresponde a una sección de las áreas ambulacrales. La membrana del periprocto se localiza en el borde de la testa; la membrana está completamente cubierta por pequeñas placas irregularmente dispuestas con los bordes irregulares y de diferentes tamaños, las placas proximales a la abertura anal, la cual está en la posición central en la membrana, tienen pequeñas proyecciones con forma de espinas que se dirigen hacia la abertura anal protegiéndola; el resto de las placas están cubiertas totalmente por tubérculos y pequeños pedicelarios oficéfalos. Peristoma dispuesto en posición central sobre la superficie oral, se localiza en el centro de la intersección de los surcos alimenticios; la boca se localiza en una cavidad cóncava muy pronunciada, pero con una cobertura reducida; la entrada de esta cavidad está completamente rodeada por pedicelarios tridentados de diferentes tamaños y espinas primarias y secundarias largas; la porción de la membrana del peristoma está muy reducida. En el centro de la boca sobresalen cinco pequeños dientes con los extremos libres romos. Las áreas interambulacrales tienen una cobertura casi rectangular sobre la testa. Las placas ambulacrales que están por fuera del sistema petalar tienen una cobertura triangular sobre la testa, siendo más estrecha en la región proximal al borde del petaloide y más ancha en la región distal (Fig. 5). En ejemplares pequeños (R ≤ 3 cm) el número de placas ambulacrales desde el borde del petaloide varía de tres a cuatro pares y nueve o diez pares de placas interambulacrales (Fig. 5A), mientras que en ejemplares de tamaño mediano a grandes (R ≥ 3.1 cm) presentan cinco, máximo seis pares de placas y 11 pares de placas interambulacrales (Fig. 5B). Las placas ambulacrales pueden ser de dos tipos: 1) con perforaciones o sin perforaciones. Las placas ambulacrales con perforaciones son las que conforman a los petaloides. Cada placa ambulacral tiene dos pares de poros ambulacrales, cada poro está dispuesto en una hilera curva que va desde el borde de la madreporita hasta la quinta placa ambulacral sobre la superficie aboral (en sentido del borde de la testa al sistema apical), las áreas poríferas están en el extremo proximal de la placa con respecto a las placas interambulacrales; la hilera de poros interna (hilera entre el área interporífera y los poros distales), siempre presenta un pequeño poro circular, mientras que en la hilera distal al área interporífera los poros se van ovalando conforme se alejan de la madreporita y aumentan su tamaño gradualmente. 2) Las placas ambulacrales que no tienen perforaciones son rectangulares y más grandes que las placas interambulacrales. En la superficie oral, las placas ambulacrales son muy anchas, casi del doble que las placas interambulacrales; el área que corresponde a la unión de las placas ambulacrales está ligeramente deprimida y cubierta por espinas primarias. Esta depresión conforma los surcos alimenticios, los cuales: son casi rectos y van desde el borde de la testa hasta el centro de ésta donde se dispone la abertura oral. En la superficie dorsal las placas interambulacrales corren desde el borde de la madreporita (a un costado de la perforación genital) hasta el borde de la testa, en la superficie oral las placas son más cuadrangulares que las ambulacrales y sólo cubren hasta el nivel de la tercera placa ambulacral proximales a la abertura oral, antes de esto, las placas interambulacrales están ausentes. Los tubérculos primarios son pequeños, la areola que los rodea es circular y muy profundidad; a los márgenes de cada tubérculo primario se disponen pequeños tubérculos secundarios, esta condición se repite irregularmente en toda la superficie de la testa, tanto en placas ambulacrales como en interambulacrales.
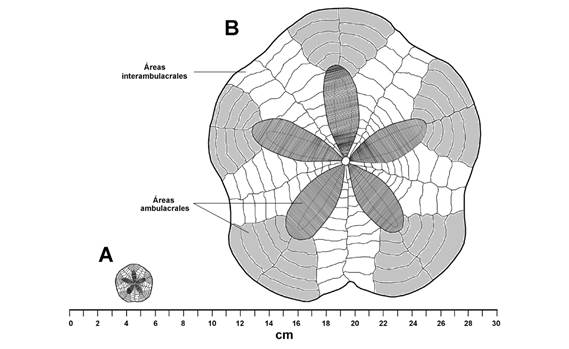
Fig. 5 Fig. 5 Clypeaster europacificus (ICML-UNAM 4.27.33). A. Ejemplar más pequeño R = 1.2 cm. B. Ejemplar más grande R = 9.2 cm. Clypeaster europacificus (ICML-UNAM 4.27.33). A. Smallest specimen R = 1.2 cm. B. Largest specimen R = 10.1 cm.
El borde del cuerpo está rodeado por espinas primarias largas (3-4 mm) con los bordes acerrados y la punta ligeramente ensanchada (Fig. 6A). Las espinas de la superficie dorsal son pequeñas y compactas (300-400 µm) con los bordes redondeados; en algunas espinas la región media es más delgada que la región terminal (Fig. 6B, 6C) dan un aspecto aterciopelado al cuerpo. Las espinas dispuestas en la sección del sistema apical, entre los petaloides son largas, con ornamentaciones en la periferia en forma de escalones que corren a lo largo de la espina, son las espinas más largas de la superficie dorsal (5-6 mm).
Espinas primarias de la superficie oral largas (3-4 mm) y delgadas (Fig. 6D), en general las espinas son ligeramente agudas y están un poco curvadas sobre la testa del organismo; las espinas primarias más proximales que rodean la boca son más curvadas que las espinas distales. Sobre la superficie de las placas interambulacrales las espinas son pequeñas (≈ 4 mm) y más robustas. Espinas secundarias delgadas sobre toda la testa, con la base más estrecha que la región terminal, rodeada por pequeñas hileras de ornamentaciones, de casi la mitad del tamaño (2-3 mm) de las espinas primarias. Presentan dos tipos de pedicelarios: tridentados y oficéfalos trivalvados. Los pedicelarios tridentados pueden tener diferentes tamaños, en la región de las placas ambulacrales que rodea la abertura oral el tallo es corto (200-250 µm), con la región terminal ligeramente abultada y con una mayor densidad del estereoma en comparación con el resto del tallo (Fig. 6E), las valvas son largas casi del mismo tamaño que el tallo y el cuello flexible del pedicelario (200-300 µm) (Fig. 6E, F). En la región cercana a los petaloides en la superficie aboral, también se disponen pequeños pedicelarios tridentados (200 µm) (Fig. 6G, H, I). Los pedicelarios tridentados se caracterizan por tener valvas con borde aserrado, un diente terminal prominente y a lo largo de toda la valva tienen pequeñas perforaciones irregulares con diferente forma, disposición y tamaño (Fig. 6F, H, I). En las placas del borde de la testa, se disponen pequeños pedicelarios oficéfalos trivalvados con el borde aserrado aún en la unión de las valvas, incluso en la sección apical curvada de estas (Fig. 6J); con los tallos largos (200-350 µm) y los cuellos muy reducidos (Fig. 6K, M); las valvas presentan un estrechamiento en su parte media, entre la región distal y la región de articulación, con el estereoma muy compacto y con pequeños dientes sobre la apófisis, la parte distal de la valva presenta de cuatro a ocho pequeñas perforaciones de formas y tamaños irregulares y con todo el borde completamente aserrado (Fig. 6L, N); este tipo de pedicelarios también se dispone en otras áreas del cuerpo como la superficie dorsal aunque son de menor tamaño, con el tallo de (40-60 µm) y valvas de 20 a 50 µm (Fig. 6M, N).
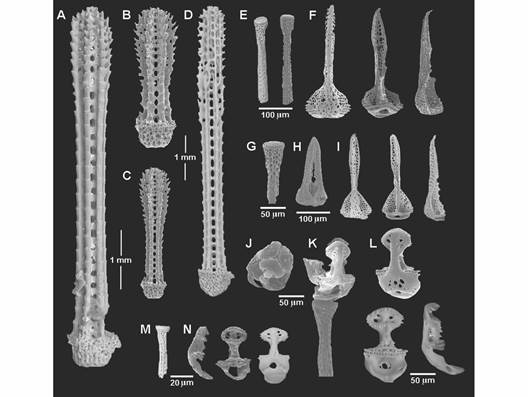
Fig. 6 Fig. 6 Clypeaster europacificus. Imágenes de MEB de las microestructuras. A. Espina primaria del borde de la testa. B-C. Espinas primarias de la superficie dorsal. D. Espina primarias de la superficie ventral. E-I. Pedicelarios tridentados. E, G. Tallos. F, I. Valvas. H. Cabeza con valvas cerradas. J-N. Pedicelarios oficéfalos. J. Vista apical de la cabeza con valvas cerradas. K. Vista lateral de la cabeza con valvas abiertas y tallo. L, N. Valvas. M. Tallo. Clypeaster europacificus. SEM images of the microestructures. A. primary spine of the test border. B-C. Primary spines from the dorsal surface. D. Primary spine from the ventral surface. E-I. Tridentate pedicellariae. E, G. Stems. F, I. Valves. H. Head with closed valves. J-N. Ophicephalous pedicellariae. J. Apical view of the head with closed valves. K. Lateral view of the head with open valves and stem. L, N. Valves. M. Stem.
Análisis de tamaño (crecimiento relativo)
El análisis de las relaciones alométricas para variables lineales indicó que las relaciones R vs. r (Fig. 7A) y R vs. h (Fig. 7C) tuvieron un crecimiento de tipo alométrico negativo (t-test, p < 0.05), en tanto que para la relación R vs. LP (Fig. 7B), el crecimiento fue de tipo alométrico positivo (t-test, p < 0.05). Para las relaciones R vs. A (Fig. 7D) y R vs. P (Fig. 7E), la relación fue de tipo alométrico negativo (t-test, p < 0.05) (Tabla 1).
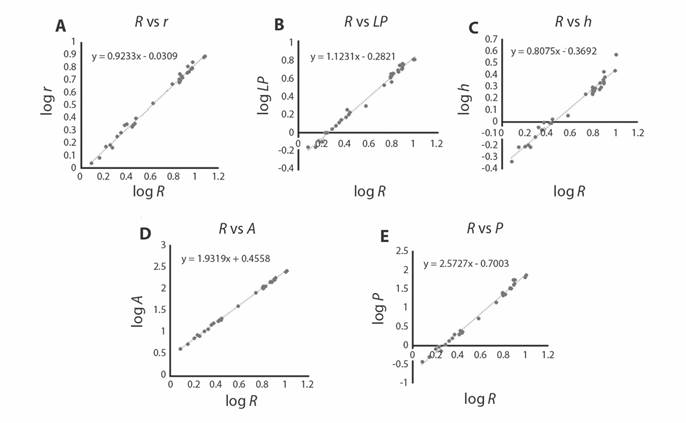
Fig. 7 Fig. 7 Regresiones lineales entre el radio mayor (R) (variable independiente) contra: A. Radio menor (r). B. Longitud del petaloide anterior impar (LP). C. Altura (h). D. Área (A). E. Peso seco (P) (variables dependientes). Linear regressions between the major radius (R) (independent variable) against: A. Minor radius (r). B. Length of the odd anterior petaloid (LP). C. Height (h). D. Area (A). E. Dry weight (P) (dependent variables).
Discusión
Clark (1914) utilizó la forma de la testa de C. europacificus para identificar a los organismos de esta especie, la forma convexa de la superficie dorsal de estos especímenes (descrita también en el material de la serie tipo) causada por la elevación de la zona central del sistema apical y del sistema petalar, está relacionada con la cavidad donde se alberga la mandíbula (Fig. 3A-3, 3B-3, 3C-3, 3D-3 y 3E-3). Durante el análisis de los ejemplares depositados en la CNE, ICML-UNAM, logramos identificar que esta característica se mantiene desde los ejemplares más pequeños analizados (R = 1.2 cm) hasta las tallas máximas evaluadas (R = 10.1 cm). Sin embargo, la diferencia en la proporción entre la altura y longitud de la testa es más evidente en los ejemplares de tallas que superan un R ≥ 3.1 cm (Fig. 3A, 3B y 3C; Fig. 4B).
En el análisis de datos se identificó la relación entre la altura de la testa y el tamaño de los organismos reflejando un crecimiento de tipo alométrico negativo (Tabla 1, Fig. 7C). Esto aunado a la variación intraespecífica y/o de la plasticidad morfológica que está relacionada con la disponibilidad de alimento detectado para otras especies (Escarcega-Quiroga, Granados-Barba, González-Gándara, Epherra, Marín-Hernández, & Crespi-Abril, 2019). Esto significa que, a pesar de la variación intraespecífica y/o de la plasticidad morfológica que está relacionada con la disponibilidad de alimento, la relación en la altura de la testa con respecto al incremento de talla (Fig. 7C) refleja un crecimiento de tipo alométrico negativo. Es probable que esta relación (h de la testa vs. crecimiento) sea la causa de porque la forma de la testa en una vista lateral sea un caracter informativo filogenéticamente (Mihaljević, Jerjen, & Smith, 2011), y que éste se haya utilizado ampliamente para identificar organismos de tallas grandes a nivel específico.
Tabla 1 Table 1 Relaciones alométricas entre pares de variables para C. europacificus recolectada en bahía Santa Inés, Baja California Sur, México Allometric relationships between pairs of variables for C. europacificus collected in Santa Ines bay, Baja California Sur, Mexico
| Variable independiente | Variable dependiente | Parámetro de crecimiento (b) | Alometría |
| R | r | 0.92 | - |
| R | LP | 1.12 | + |
| R | h | 0.81 | - |
| R | A | 1.93 | - |
| R | P | 2.57 | - |
Variable independiente: radio mayor (R). Variables dependientes: longitud del petaloide anterior impar (LP), radio menor (r), altura (h), área (A) y peso seco (P).
Independent variable: major radius (R). Dependent variables: length of the odd anterior petaloid (LP), minor radius (r), height (h), area (A) and dry weight (P).
Actualmente para clasificar clipeastéridos los caracteres más utilizados para la identificación taxonómica de especímenes son, la forma del cuerpo, así como el tamaño de los petaloides o las proporciones de cobertura sobre la testa (Mooi, 1989). La forma pentagonal del cuerpo en C. europacificus es una de las características más distintivas que poseen los organismos de tallas medianas a grandes (R ≥ 3.1 cm). En estos organismos las placas correspondientes a las áreas ambulacrales son de mayor tamaño en comparación a las placas interambulacrales, lo que le da un aspecto ligeramente pentagonal (con los bordes curvados) (Fig. 3A, B y C; Fig. 4B). Por otro lado, en ejemplares de la misma localidad que en tallas menores (R ≤ 3 cm) la forma del cuerpo es completamente circular (Fig. 3D y E; Fig. 4A), un aspecto que comparte con otras especies e incluso otros grupos de clipeastéridos.
Al igual que en otras galletas de mar, los especímenes de C. europacificus tienen etapas durante el desarrollo en la que se asemejan a la morfología de otros clipeastéridos, es por esto que durante la descripción de Clypeaster europacificusMortensen (1948) mencionó la semejanza morfológica con especímenes de C. euclastusClark, 1941; pero a diferencia de C. europacificus, C. euclastus siempre conserva la forma circular de la testa sin importar la talla de los individuos. Además, la característica más notable para diferenciar entre especies es la forma de los petaloides, los cuales son mucho más abiertos distalmente en C. euclastus, mientras que en C. europacificus están más cerrados (Fig. 3A; Fig. 1A, B, C, D y E), y por la posición inframarginal del periprocto en C. europacificus y ventral en C. euclastus (Francisco & Pauls, 2008: 218, Fig. 1; USNM 1151419).
La especie más parecida a ejemplares medianos o grandes de Clypeaster europacificus es C. ravenelii (Agassiz, 1869) por la forma pentagonal de la testa. C. ravenelli se diferencia de C. europacificus por poseer un borde abultado de las placas marginales de la superficie dorsal (Mihaljević et al., 2011: 26, Fig. 2G) a diferencia del borde delgado presente en C. europacificus (Fig. 3A, B, C, D y E). Por lo anterior, es importante mencionar que la forma de la testa es un caracter morfológico que por sí sólo carece de fuerza al momento de realizar una identificación taxonómica. Sin embargo, existen caracteres de la testa que se conservan desde las primeras etapas del desarrollo como la forma y proporción de los petaloides y la posición del periprocto en la superficie de la testa. El cambio en la forma de los organismos se relaciona con el hábitat y las condiciones de este, por lo que podríamos hipotetizar que, en el caso de los especímenes pequeños de C. europacificus después de alcanzar cierta talla (R ≥ 3.1 cm), los erizos migran a ambientes con otras condiciones del sustrato en las que se ven favorecidos al incrementar de tamaño para llevar a cabo funciones básicas como la alimentación y locomoción, como se ha demostrado para otros equinoideos (Seilacher, 1979).
Con respecto a las microestructuras calcáreas que se evaluaron (espinas primarias, secundarias y pedicelarios), es difícil poder detectar descriptivamente una correlación con el crecimiento del organismo y la forma o tamaño de estas estructuras por diversos motivos. En el caso de las espinas primarias y secundarias, al ser estructuras que están asociadas a funciones de defensa (Moureaux et al., 2010), alimentación y locomoción (Fu-Shiang, 1969) es común encontrar evidencias de regeneración (Vinnikova & Drozdov, 2011) ya sea por extracción completa o fragmentación de las espinas, lo que dificulta establecer los límites morfológicos que están ligados con el crecimiento y a mecanismos regenerativos. Algo muy parecido ocurre con los pedicelarios tridentados y oficéfalos (Coppard & Campbell, 2006; Coppard, Kroh, & Smith, 2010), los cuales tienen asociados funciones de limpieza y defensa del organismo y durante estas actividades es que sufren daño o bien son sustituidos por estructuras nuevas, lo que dificulta su delimitación morfológica y correlación con el crecimiento de los especímenes (Dubois & Ameye, 2001).
Crecimiento relativo. Las relaciones alométricas efectuadas indican la presencia de cambios en la forma corporal de C. europacificus durante su crecimiento (Fig. 7). Con respecto a los parámetros lineales, la alometría negativa en la relación R vs. r denota que durante el crecimiento la distancia entre el borde del sistema apical y la testa (sobre el ambulacro II) aumenta a un ritmo mayor en comparación al ritmo en el que cambia el interambulacro 5. Lo anterior se ve reflejado morfológicamente al momento de comparar el espécimen más pequeño evaluado (R = 1.213 cm y r = 1.095 cm) con una forma del cuerpo circular (Fig. 5A) donde la diferencia de la longitud del ambulacro II y del interambulacro 5 es muy pequeña (0.118 cm), mientras que en el espécimen más grandes (R = 10.19 cm y r = 7.77 cm) la longitud del ambulacro II es 2.41 cm más grande que la longitud del interambulacro 5, medidas de un espécimen con forma pentagonal (Fig. 5B). Esta misma relación se demostró en el trabajo de Swisher y Lin (2019) para Arachnoides placenta durante su crecimiento. Es importante señalar que el cambio en la forma del borde de la testa presente en C. europacificus durante su crecimiento, es un fenómeno que no se comparte en todas las especies de clipeastéridos, p. e. Zachos (2015) reportó que en el análisis que realizó para Echinarachnius parma la testa mostró un crecimiento casi simétrico (isométrico), en donde el aumento de tamaño en las placas desde los organismos post-metamorfosis (con 38 placas) hasta los organismos adultos (con más de mil placas) se conserva la proporción entre las secciones de la testa, con la forma de la misma durante el crecimiento de los organismos. Para ejemplares pequeños de C. europacificus el número de placas para la superficie dorsal fue de 40 placas ambulacrales (sin contar las incluidas en el sistema petalar) y 100 placas interambulacrales, mientras que en ejemplares de tallas medianas a grandes, los especímenes presentan 60 placas ambulacrales y 110 placas interambulacrales.
La alometría positiva en la relación R vs. LP (Fig. 7B) y negativa (Tabla 1) en la relación R vs. h (Fig. 7C) indican que no todas las variables crecen al mismo ritmo, es decir, R crece a un menor ritmo que LP, pero mayor al de h. Por otro lado, la alometría negativa resultante de las relaciones R vs. A (Fig. 7D) y R vs. P (Fig. 7E) (Tabla 1) indican que, con el crecimiento, el área y el volumen de la galleta crecen menos con relación al radio mayor, esto significa que tanto el área como el volumen de la galleta, son menores a la de un círculo y una esfera de radio R. No existen antecedentes acerca del valor del parámetro b de crecimiento para C. europacificus; sin embargo, los valores aquí proporcionados (Tabla 1) ayudan a enriquecer los criterios empleados por la taxonomía para identificar a las especies, o bien, para identificar diferencias morfológicas entre poblaciones de la misma especie.
Durante el crecimiento en Clypeaster europacificus los ejemplares menores a 3 cm de radio presentan en promedio 140 placas en la superficie dorsal y a partir de los 3.1 cm de radio 170 placas. Este incremento en el número de placas altera las proporciones de la testa en los estudios iniciales del crecimiento a diferencia de los estadios finales cuando los especímenes tienen una forma pentagonal. En especímenes pequeños de C. europacificus, la forma circular de la testa se asemeja mucho a la de C. euclastus y al adquirir una forma pentagonal en tallas medianas o grandes, se asemeja morfológicamente a C. ravenelli. El crecimiento de las placas ambulacrales e interambulacrales que produce un incremento en el área de la testa y el peso del organismo tienen una relación alométrica negativa. C. europacificus es una especie de galleta de mar que apartir de los 3 cm de radio comienza a cambiar la forma general del cuerpo de un borde circular a uno pentagonal.
A pesar de que en el presente trabajo se realizó un esfuerzo para identificar los cambios morfológicos durante el crecimiento de C. europacificus, aún faltan resolver algunas interrogantes que podrían ser evaluadas en trabajos posteriores, como identificar la relación entre las dimensiones de la linterna de Aristóteles y la altura de testa, o bien, identificar si existen variaciones morfológicas a lo largo de toda su distribución latitudinal y de manera experimental, evaluar si la disponibilidad de alimento y las condiciones asociadas con el sedimento (granulometría y profundidad dentro del sedimento) tienen un efecto sobre la morfología de la testa.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

