











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Las selvas albergan más de la mitad de la biodiversidad del planeta y proveen importantes bienes y servicios (Balvanera, 2012; Barlow et al., 2018), de estas una porción importante se desarrolla en ambientes kársticos entre los que destacan los ubicados en el sureste de China, Centroamérica y la Península de Yucatán en México (Geekiyanage et al., 2019).
Al sur de la Península de Yucatán en la región de Calakmul, se ubican las selvas que forman junto con la reserva privada de Rio Bravo en Belice y la Reserva de la Biosfera Maya en Guatemala el área de selva más grande de Mesoamérica (Ellis et al., 2017). Estas selvas albergan seis tipos de vegetación y 23 asociaciones, entre las que destacan las selvas perennifolias alta y mediana, la selva mediana subperennifolia (dominante en la región), las selvas bajas subperennifolia, subcaducifolia y caducifolia (Martínez & Galindo Leal, 2002). Estos tipos de vegetación están asociados a gradientes topográficos, de dinámica de nutrientes y condiciones edáficas que se caracterizan por la presencia de terrenos kársticos, con suelos someros y abundantes afloramientos de calcio que reducen la disponibilidad de agua (Wendt, 1993; Martínez & Galindo Leal, 2002; Bautista & Palacio-Aponte, 2011; Chiquini-Heredia et al., 2017; Estrada-Medina et al., 2019). Estos afloramientos de calcio pueden ser colonizados por especies de la familia Sapotaceae, siendo la selva baja y el bajo mixto los tipos de vegetación que se han reportado en estos sitios (Martínez & Galindo Leal, 2002).
La diversidad y estructura de diferentes tipos de vegetación en Calakmul han sido estudiadas por distintos autores (Vester et al., 2007; Zamora-Crescencio et al., 2012; García-Licona et al., 2014; Báez-Vargas et al., 2017; Chiquini-Heredia et al., 2017; Esparza-Olguín & Martínez-Romero, 2018; Esparza-Olguín et al., 2019). Sin embargo, hasta ahora no ha sido descrita y analizada la diversidad, estructura y composición de selvas que se desarrollan en afloramientos de calcio conocidos localmente como yesales.
Los suelos kársticos en los yesales poseen baja capacidad de retención hídrica, dado el alto grado de infiltración del agua al subsuelo, lo que dificulta la absorción de agua por parte de las plantas y acentúa su sensibilidad a sequías estacionales (García-Gil et al., 2002; Martínez & Galindo-Leal, 2002; Estrada-Medina et al., 2019; Geekiyanage et al., 2019). Además, debido a una baja fijación de fósforo (P) y nitrógeno (N) (Bautista & Palacio-Aponte, 2011; Liu et al., 2018; Estrada-Medina et al., 2019; Geekiyanage et al., 2019), constituyen ambientes restrictivos para el desarrollo de las plantas (Du et al., 2011; Fenton et al., 2017; Pan et al., 2018; Geekiyanage et al., 2019). Las especies vegetales que se desarrollan sobre este tipo de afloramientos pueden ser especialistas con adaptaciones para hacer frente a las limitaciones de nutrientes en estos suelos, o generalistas que toleran las limitaciones asociadas a estos afloramientos (Geekiyanage et al., 2019).
En este estudio se analizan la composición, diversidad y estructura de la vegetación arbórea que se desarrolla en los yesales y las condiciones edáficas de los mismos, comparando estos atributos con los que se presentan en la vegetación secundaria (VS) circundante. La hipótesis planteada es que la composición, diversidad y estructura serán distintas en la vegetación arbórea de los yesales respecto a las de VS circundante. Se espera que los yesales presenten una menor riqueza y diversidad, así como valores promedio de altura y diámetro más bajos debido a las condiciones del suelo.
Materiales y métodos
Área de estudio: El trabajo se realizó en el ejido Once de Mayo (18°5’28.11” N & 89°27’40.82” W), ubicado en el sureste del municipio de Calakmul y de la Reserva de la Biosfera de Calakmul. El clima es cálido subhúmedo Aw2(x´) (i´) g, con alta humedad, con régimen de lluvias en verano. La temperatura media anual es de 26 ºC, mientras que la precipitación anual fluctúa entre 1 100 y 1 500 mm. Los suelos son litosoles con pequeñas áreas de rendzinas (García-Gil et al., 2002; Bautista et al., 2011).
Diseño de muestreo: En junio de 2016, se realizaron recorridos por el ejido para delimitar el polígono del afloramiento de calcio con un área total de 14.5 ha. Posteriormente, se determinó el tipo de vegetación que rodea al afloramiento empleando el método semi-cuantitativo, que permite caracterizar la cobertura vegetal mediante rasgos de fisonomía, estructura física y composición florística (Jardel-Peláez, 2015; Aguilar-Duarte et al., 2019). A partir de la información recabada en la vegetación circundante, fue posible observar que la composición y estructura de esta correspondía a la de vegetación secundaria derivada de selva subperennifolia. Por otro lado, se documentó mediante entrevistas que la vegetación presente en el afloramiento de calcio no ha sido usada desde hace más de 65 años; mientras que el área de VS aledaña fue usada para pastoreo de ganado ovino y la siembra de chile en la década de los noventa y principio de los años 2000, actualmente se extrae leña del área. Posteriormente, se generó un mapa del polígono del afloramiento y la vegetación secundaria (VS) circundante para ubicar al azar 17 unidades de muestreo (14 en yesales y 3 en VS); así como los puntos para la toma de muestras de suelo en cada unidad.
Las unidades de muestreo fueron parcelas circulares de 1 000 m² (17.84 m de radio), que se subdividieron en ocho secciones para facilitar el censo de cada individuo arbóreo presente.
Características edáficas: El muestreo de suelos se realizó con base en la Norma Oficial Mexicana (NOM-021-RECNAT-2000), que establece las especificaciones de fertilidad, salinidad y clasificación de suelos; así como su estudio, muestreo y análisis (Secretaría de Medio Ambiente y Recursos Naturales, 2002). En cada una de las 17 parcelas se tomaron cinco muestras de suelo; una en el centro y cuatro en los límites con dirección a los puntos cardinales (norte, este, sur y oeste) para la obtención de una muestra compuesta por parcela. Para cada muestra se delimitó un área de 0.3 x 0.3 m la cual se excavo con ayuda de una pala pequeña hasta los 0.3 m de profundidad. Con las cinco muestras de cada parcela se prepararon muestras compuestas homogéneas de 1.5 kg en tinas de plástico con capacidad para 30 kg, evitando la contaminación entre muestras. Las muestras se enviaron a un laboratorio de suelos (Fertilab) para su análisis fisicoquímico estimando: pH, conductividad eléctrica (CE-salinidad), % de carbonatos de calcio (CaCO3), materia orgánica (MO), contenido de fósforo (P) y nitrógeno (N).
Composición florística, diversidad de especies y estructura de la vegetación arbórea: Cada individuo arbóreo ubicado en las parcelas se etiquetó e identificó, tomando los datos de nombre común y nombre científico. La identificación taxonómica se realizó con la ayuda de dos parataxónomos, apoyados con el listado de Martínez et al., (2001). La nomenclatura se corroboró en la página especializada http://www.worldfloraonline.org (World Flora Online, 2021). Se construyó el listado florístico con los datos de especie, género y familia botánica. Para analizar la similitud entre los yesales y VS, se hizo un análisis de coordenadas principales (PCoA), mediante el índice de Bray y Curtis (1957) empleando el software PAST 4.06b (Hammer et al., s.f.).
La diversidad se estimó mediante los índices de diversidad verdadera de orden q = 0 (riqueza), q = 1 (Shannon) y q = 2 (Simpson), mediante la expresión general qD = (S ∑ i = 1 pqi) 1/(1-q), donde, qD es la diversidad verdadera, pi es la abundancia relativa (abundancia proporcional) de la iésima especie, S es el número de especies, y q es el orden de la diversidad. Considerando la necesidad de estandarizar el tamaño y la completitud de la muestra para poder comparar las condiciones analizadas, se emplearon los métodos de rarefacción y extrapolación propuestos por Chao et al., (2014).
Además, se midió la altura total y el diámetro a la altura de 1.3 m (diámetro normal) o en individuos menores a 1.30 m de altura el diámetro en la base del tronco (diámetro al cuello). Con estos datos se obtuvieron el diámetro y la altura promedio por parcela y condición (yesales y VS).
La comparación de los valores de cada una de las variables de las condiciones edáficas entre los yesales y VS, se hizo mediante un análisis de varianza (ANOVA). Para determinar homogeneidad de varianza, previo al ANOVA, se realizó una prueba de Levene en todas las variables. La CE (salinidad) no cumplió con este requisito por lo que fue necesario transformarla aplicando log10. Por otro lado, la comparación de las variables de estructura entre los yesales y la VS se realizó con una prueba de Kruskal-Wallis.
Para examinar diferencias en la composición de especies entre los yesales y la VS se realizó un análisis de similitud (ANOSIM), utilizando la distancia del índice de Bray Curtis (Clark, 1993; Magurran, 2004). Este análisis no paramétrico estima una medida absoluta de distancia entre los grupos (R), con valores entre 0 y 1, la similitud es mayor entre las muestras cuanto más se acerca a 0. El nivel de significancia se calculó mediante la prueba de Bonferroni. La proporción que aporta cada especie a la diferencia entre yesales y VS se estimó mediante el porcentaje de disimilitud entre ellas, empleando la subrutina SIMPER (Clark, 1993). Estos análisis se realizaron con el programa PAST 4.06b (Hammer et al., s.f.).
Para determinar posibles relaciones entre las variables edáficas y el diámetro promedio, la altura promedio y la diversidad verdadera (1D), se realizaron análisis de regresión linear mediante el software Statistica 13.0.
Resultados
Características edáficas: Al comparar cada una de las variables edáficas entre los yesales y VS (Tabla 1) se observó que la única variable que no presentó diferencias significativas fue el pH, que tuvo valores que oscilaron entre 7.5 y 8 (moderadamente alcalino). Los suelos en los yesales presentaron bajas concentraciones de MO, P disponible y N, con CE-salinidad de media a alta y altos porcentajes de CaCO3. En el caso de las variables N (F = 99.20, P = 0.00017), P (F = 205.00, P = 0.00017) y MO (F = 99.18, P = 0.0002), los yesales presentaron valores significativamente menores que los encontrados en la VS, teniendo en contraste valores significativamente mayores de porcentaje de CaCO3 (F = 20.93, P = 0.0005) y conductividad eléctrica (CE-salinidad, F = 35.25, P = 0.00019).
Composición florística, diversidad de especies y estructura de la vegetación arbórea: En total se registraron 7 025 individuos, 6 443 en los yesales y 594 en la VS circundantes. En los yesales se encontraron 54 especies pertenecientes a 25 familias y en VS 62 especies y 27 familias (Apéndice digital). En los yesales las familias con mayor número de especies fueron: Fabaceae (8), Rubiaceae (5), Euphorbiaceae (4), Polygonaceae (4) y Sapotaceae (4), aportando el 46.3 % del total de especies. En la VS circundante las familias que destacaron fueron: Fabaceae (13), Polygonaceae (5), Sapotaceae (4) y con 3 especies Apocynaceae, Myrtaceae, y Rubiaceae, lo que representa el 50 % de las especies totales.
Las especies más abundantes en los yesales fueron: Cameraria latifolia L. (1917), Bonellia flammea (Millsp. ex Mez) B.Ståhl & Källersjö (677), Metopium brownei (Jacq.) Urb. (523), Diospyros bumelioides Standl. (497) y Eugenia karwinskyana O.Berg (462). En la VS las especies que destacaron fueron: Hampea trilobata Standl. (74), Croton icche Lundell (45), M. brownei (40), Sideroxylon salicifolium (L.) Lam. (25) y Eugenia ibarrae Lundell (17). Entre los yesales y la VS se comparten 27 especies, siendo las más abundantes C. latifolia, M. brownei, Gymnopodium floribundum Rolfe y Haematoxylum campechianum L. (Apéndice digital).
El ANOSIM mostró que la composición de especies entre yesales y VS fue significativamente diferente (R = 0.9904, P = 0.0016). Según el análisis SIMPER se presentó una disimilitud del 90.96 % entre yesales y VS; las especies que contribuyen a estas diferencias se presentan en la Tabla 2. Estos resultados son congruentes con el PCoA, en el que las dos primeras coordenadas representaron el 55.7 % de la variabilidad (33.87 y 17.87 % respectivamente), que permitió reconocer que las parcelas de VS son diferentes de las de parcelas de yesales, es decir presentan una composición de especies distinta (Fig. 1), con condiciones edáficas significativamente diferentes (Tabla 1).
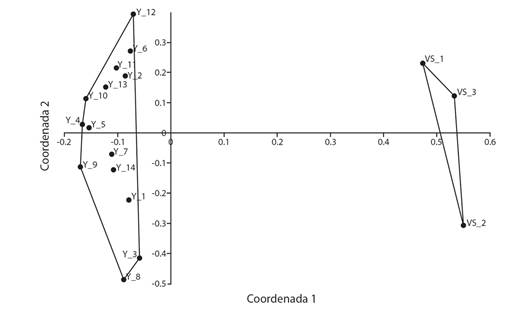
Fig. 1 Análisis de coordenada principales (PCoA) entre parcelas estudiadas en yesales y vegetación secundaria en Once de Mayo, Calakmul, Campeche. Fig. 1. Principal Coordinates Analysis (PCoA) between plots of yesales and secondary vegetation in Once de Mayo, Calakmul, Campeche.
TABLA 1 Valores promedio de los diferentes parámetros edáficos y estructurales evaluados en yesales y vegetación secundaria en Once de Mayo, Calakmul, Campeche. Table 1. Average values of the different edaphic and structural parameters evaluated in yesales and secondary vegetation in Once de Mayo, Calakmul, Campeche
| Condición | MO (%) | pH | CaCO3 (%) | CE | P (ppm) | N (%) | Altura (m) | Diámetro (cm) |
| Yesales | 0.47(± 0.11 ) | 7.68 ± (0.05) | 39.66 (± 2.03) | 4.04 (±0.42) | 2.33 (± 0.19) | 0.09 (± 0.02) | 1.05 (± 0.07) | 2.87 (± 0.09) |
| VS | 2.97(± 0.15) | 7.60 ± (0.06) | 17.93 (± 3.77) | 0.66 (±0.06) | 10.33 (± 0.88) | 0.59 (± 0.03) | 7.04 (± 0.27) | 8.47 (± 1.09) |
Los datos se presentan como valores promedio ± el error estándar (entre paréntesis). MO = materia orgánica, CE = conductividad eléctrica, P = fósforo, N = nitrógeno.
TABLA 2 Especies que contribuyen con los mayores porcentajes de disimilitud encontradas en yesales y vegetación secundaria en Once de Mayo, Calakmul, Campeche. Table 2. Species that contribute the highest percentages of dissimilarity found in yesales and secondary vegetation in Once de Mayo, Calakmul, Campeche
| Especie | Pd | % Contr. | % Contr. A |
| Cameraria latifolia L. | 18.96 | 20.8 | 20.84 |
| Bonellia flammea (Millsp. ex Mez) B.Ståhl & Källersjö | 8.21 | 9.0 | 29.87 |
| Metopium brownei (Jacq.) Urb. | 4.66 | 5.1 | 34.99 |
| Diospyros bumelioides Standl. | 4.42 | 4.9 | 39.86 |
| Eugenia karwinskyana O.Berg | 4.32 | 4.8 | 44.62 |
| Gymnopodium floribundum Rolfe | 4.29 | 4.7 | 49.33 |
| Haematoxylum campechianum L. | 3.89 | 4.2 | 53.62 |
| Hampea trilobata Standl. | 3.74 | 4.1 | 57.73 |
Promedios de disimilitud (Pd); porcentaje de contribución (% Contr.); porcentaje de contribución acumulado (% Contr. A).
En las curvas de diversidad con rarefacción y extrapolación por muestras, se observa que los valores de diversidad verdadera evaluados (riqueza, Shannon y Simpson) son significativamente menores en los yesales respecto de la VS (Fig. 2A). En relación con la completitud del inventario se observa que en el caso de los yesales está fue de 88.5 %; mientras en la VS fue de 52.9 % para las tres unidades muestreadas y de 79.5 % respecto a la extrapolación realizada para 14 unidades de muestreo (Fig. 2B). En cuanto a los valores de altura y diámetro, se encontraron diferencias significativas entre yesales y VS, siendo significativamente menor en ambos casos para los yesales (Tabla 1).
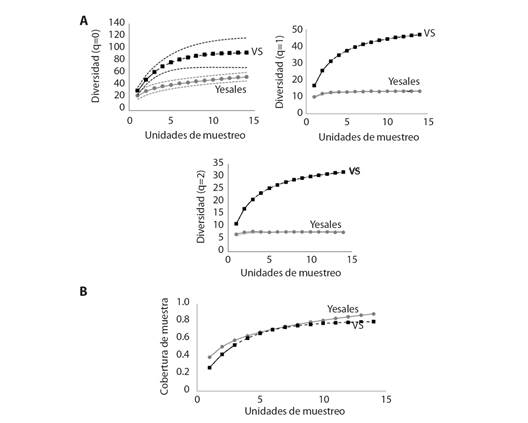
Fig. 2 A. Curvas de rarefacción (líneas continuas) y extrapolación (líneas punteadas) basadas en el tamaño de muestra de la diversidad de especies arbóreas presentes en yesales y vegetación secundaria (VS). B. Curva de completitud del inventario para las muestras con rarefacción (líneas continuas) y con extrapolación (líneas punteadas) de especies arbóreas presentes en yesales y vegetación secundaria (VS) en Once de Mayo, Calakmul, Campeche. Fig 2. A. Rarefaction curves (solid lines) and extrapolation (dotted lines) based on the sample size of the diversity of tree species present in yesales and secondary vegetation (VS). B. Inventory completeness curve for samples with rarefaction (solid lines) and with extrapolation (dotted lines) of tree species present in yesales and secondary vegetation (VS) in Once de Mayo, Calakmul, Campeche.
Al analizar la posible relación entre las diferencias en los valores de altura promedio, diámetro promedio y diversidad respecto de las condiciones edáficas presentes en yesales y VS, se encontró que en todos los casos se presentan relaciones significativas (Fig. 3, Fig. 4, Fig. 5). Los valores de diversidad (1D), altura y diámetro aumentan conforme se presentan mayores cantidades de N, P y MO; mientras que menores porcentajes de CaCO3 y la disminución en los valores de CE (salinidad) se relacionan con un aumento en la diversidad, la altura y el diámetro.
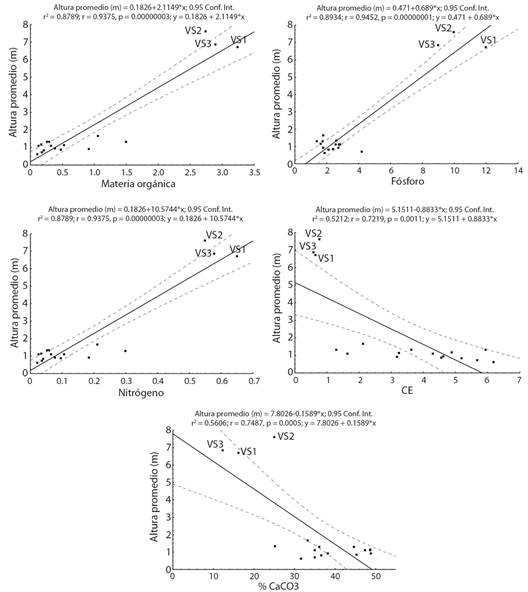
Fig. 3 Relación entre altura promedio de especies leñosas y las condiciones edáficas presentes en yesales y vegetación secundaria en Once de Mayo, Calakmul, México. Fig. 3. Relationship between average height of woody species and the edaphic conditions present in yesales and secondary vegetation in Once de Mayo, Calakmul, Mexico.
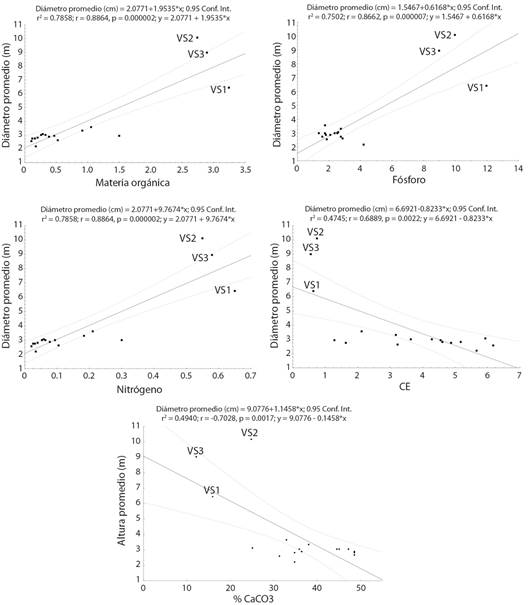
Fig. 4 Relación entre diámetro promedio de especies leñosas y las condiciones edáficas presentes en yesales y vegetación secundaria en Once de Mayo, Calakmul, México. Fig. 4. Relationship between average diameter of woody species and the edaphic conditions present in yesales and secondary vegetation in Once de Mayo, Calakmul, Mexico.
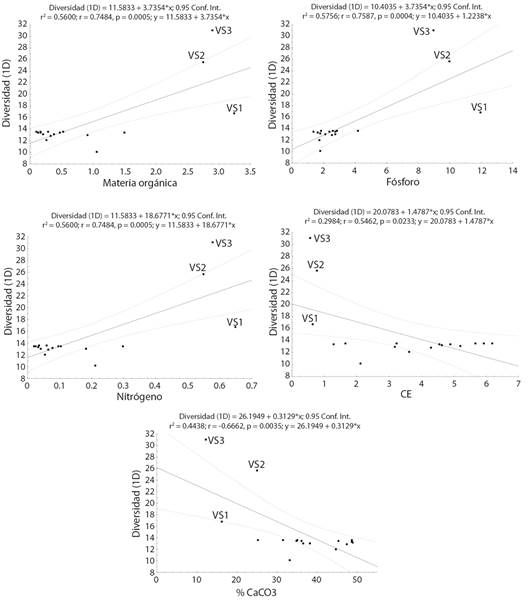
Fig. 5 Relación entre diversidad (1D) de especies leñosas y las condiciones edáficas presentes en yesales y vegetación secundaria en Once de Mayo, Calakmul, México. Fig. 5. Relationship between diversity (1D) of woody species and the edaphic conditions present in yesales and secondary vegetation in Once de Mayo, Calakmul, Mexico.
Discusión
Diversos autores han reportado bajas cantidades de MO, P y N en suelos kársticos, como es el caso de las condiciones edáficas estimadas para yesales en este estudio (García et al., 2005; Jiang et al., 2006; Zhang et al., 2006; Du et al., 2011; Liu et al., 2018; Geekiyanage et al., 2019; Wang et al., 2019). Estas condiciones edáficas están asociadas a la alta concentración de CaCO3 que afecta la estabilización de la MO lo que genera una baja disponibilidad de N y P, pues ambos elementos se unen al Ca, lo que dificulta que queden libres para ser absorbidos por las plantas (Hu et al., 2012; Pan et al., 2016). Además, los nutrientes disueltos se lixivian fácilmente en los porosos suelos kársticos lo que limita aún más la disponibilidad de N y P (Zhang et al., 2011; Pan et al., 2016; Geekiyanage et al., 2019; Wang et al., 2019). Los valores encontrados para yesales y VS, respecto a estos tres elementos de la fertilidad del suelo, concuerdan con estudios que han señalado valores particularmente bajos para el P y el N en áreas abiertas o con vegetación secundaria en fases tempranas que aumentan conforme avanza la sucesión (Jiang et al., 2006; Zhu et al., 2012; Aryal et al., 2015).
En contraste al efecto de los altos porcentajes de CaCO3 en la MO, la CE, es decir la salinidad es afectada positivamente (Umer et al., 2020), lo que se refleja en valores altos como los encontrados en los yesales, que concuerdan con lo reportado por Chang et al., (2018) para el área con suelos kársticos en China.
La composición de especies en los yesales es semejante a la que se ha reportado para selvas subperennifolias en Calakmul, donde destacan especies de las familias Fabaceae, Rubiaceae y Sapotaceae (Díaz-Gallegos et al., 2002; Lawrence et al., 2004; Vester et al., 2007; Zamora-Crescencio et al., 2012; García-Licona et al., 2014; Esparza-Olguín & Martínez-Romero, 2018; Esparza-Olguín et al., 2019). Estas familias se encuentran también entre las diez más frecuentes y con mayor número de especies en la península de Yucatán (Ibarra-Manríquez et al., 1995; Carnevali et al., 2010; Pérez-Sarabia et al., 2017). En cuanto a las especies más abundantes en los yesales, todas han sido reportadas para selvas bajas y medianas en Calakmul y otras áreas de la península de Yucatán (Ibarra-Manríquez et al., 1995; Carnevali et al., 2010; Ochoa-Gaona et al., 2018).
La diversidad estimada para los yesales fue menor a la reportada en otros estudios para las selvas subperennifolias de Calakmul; en contraste el valor estimado para la VS es semejante a los estimados en otros estudios (Díaz-Gallegos et al., 2002; Báez-Vargas et al., 2017; Esparza-Olguín & Martínez-Romero, 2018; Esparza-Olguín et al., 2019). A pesar de la diferencia en el tamaño de la muestra entre yesales y VS, los resultados de los análisis de rarefacción y extrapolación permiten sugerir que la diferencia en la composición de especies y la diversidad entre yesales y VS, puede explicarse por las condiciones edáficas contrastantes a las que están asociadas, lo que quedó de manifiesto al analizar la relación entre los valores de diversidad y los correspondientes a MO, N y P. En este sentido, diversos autores han reportado que la distribución de las especies arbóreas en áreas tropicales kársticas están fuertemente relacionadas con las condiciones edáficas (Pérez-García et al., 2009; Guo et al., 2017; Wang et al., 2019); presentando especies que no necesariamente muestran adaptaciones específicas para este tipo de suelo pero que toleran las limitaciones químicas o físicas asociadas o bien que desarrollan relaciones con la microbiota del suelo para poder tolerar dichas limitaciones (Zhu et al., 2012; Lu et al., 2014; Pan et al., 2018; Geekiyanage et al., 2019; Wang et al., 2019).
Si bien la composición de especies en los yesales es semejante a la que se ha reportado para selvas subperennifolias en Calakmul y la península de Yucatán, sus características de la estructura horizontal (diámetro) y vertical (altura) son menores a las reportadas para estas selvas en la región a pesar de tratarse de especies arbóreas con individuos que ya son reproductivos. Los valores de diámetro y de altura de la vegetación estimados para yesales en este estudio son menores a las reportadas para áreas con selva baja subperennifolia e incluso para vegetación secundaria en fase temprana, es decir, con hasta cinco años de recuperación en la región de Calakmul (Díaz-Gallegos et al., 2002; Lawrence et al., 2004; Vester et al., 2007; García-Licona et al., 2014). Por ejemplo, Lawrence et al., (2004) documentaron que, en vegetación secundaria en fase temprana, los diámetros dominantes oscilan entre 1 y 5 cm; mientras Vester et al., (2007) reportan, para vegetación secundaria con entre 2 y 5 años en recuperación, un diámetro promedio de hasta 8.4 cm y una altura de 7.2 m. La relación significativa entre los valores promedio de altura y diámetro en los yesales respecto a la cantidad de MO, P y N disponibles, así como al porcentaje de CaCO3 y la CE, indican que las bajas tallas, tanto en altura como en diámetro, de los individuos arbóreos en los yesales pueden explicarse a partir de la poca disponibilidad de nutrientes (García et al., 2005; Jiang et al., 2006; Zhang et al., 2006; Du et al., 2011; Liu et al., 2018; Geekiyanage et al., 2019), la alta concentración de CaCO3 que limita aún más la disponibilidad de P y N (Hu et al., 2012; Pan et al., 2016), y los altos valores de CE que afectan la absorción de nutrientes por parte de las plantas (Chang et al., 2018).
En conclusión, la vegetación presente en los yesales tiene una composición semejante a la presente en selvas subperennifolias de Calakmul. Sin embargo, como se esperaba las características estructurales horizontal (diámetros) y vertical (altura) son menores incluso a las que se presentan en vegetación secundaria en etapas tempranas. Lo anterior, a pesar de presentar individuos reproductivos de las especies más abundantes y de tratarse de vegetación que no ha sido usada en al menos 65 años. Estas tallas pequeñas en los individuos arbóreos están relacionadas con el alto porcentaje de CaCO3 y los altos valores de CE que condicionan en parte la poca disponibilidad de MO, N y P afectando el crecimiento de los árboles. Los resultados de este estudio contribuyen a sustentar la idea de que condiciones edáficas precarias afectan negativamente la diversidad y estructura (horizontal y vertical) de la vegetación arbórea en suelos kársticos.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

