











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Los huracanes o ciclones tropicales son masas de aire convectivo que pueden alcanzar diámetros de hasta 600 km, con vientos oscilatorios entre los 119 y los 300 km/h (Meléndez, 2015). Estos fenómenos se forman en las cuencas del Atlántico norte y se desplazan hacia el oeste, e impactan en la mayoría de los casos contra el litoral sur de Estados Unidos y México, con tendencia en los últimos años a desplazarse hacia Centroamérica (Collins & Roache, 2017). Por esta razón se prevé un aumento en la incidencia e intensidad de huracanes debido al incremento en la temperatura de la superficie marítima por el cambio climático global (Sobel et al., 2016; Walsh et al., 2016; Wang, Liu, & Foltz, 2017). Además, Allen, Tippett y Sobel (2015); Collins et al. (2016); y Murakami et al. (2017) indicaron que valores extremos en temperatura, humedad y velocidad del viento producidos por el fenómeno del niño contribuyen con la formación de tormentas tropicales y huracanes. Asimismo, estudios recientes demuestran que estos cambios abruptos en las condiciones climáticas son los principales factores que influyen en la contracción de ecosistemas (Gabler et al., 2017; Feher et al., 2017; Osland et al., 2017).
La temporada de huracanes 2015-2016 cerró la actividad en noviembre del 2016 con el huracán Otto, históricamente fue de los pocos huracanes que lograron desplazarse tan al sur del continente y cruzar hasta las cuencas del pacífico oriental, atravesó tierra en la región fronteriza entre Costa Rica y Nicaragua (Collins & Roache, 2017); convirtiéndose así en el primer registro oficial de impacto directo de un huracán en el territorio costarricense (Brown, 2017). Sin embargo, Lizano y Fernández (1996) indicaron que es posible que este hecho haya sucedido en el pasado, en el siglo XIX. Además, se tienen registros de impacto indirecto por los huracanes Kattie, Gilbert, César y Mitch (Díaz, 2004).
En la zona norte de Costa Rica, el huracán Otto causó grandes impactos en bosques primarios y secundarios, con reducciones de hasta el 30 % en los valores de área basal y del 35 % del número de individuos registrados por hectárea, aunque los índices de diversidad no presentaron diferencias significativas con los valores medidos antes del evento (Araya-Orozco, 2017). Se podría esperar que los principales daños en estos bosques sean coincidentes con otros casos de la región, como México y Puerto Rico, donde las mayores perturbaciones corresponden al vuelco, descope y quiebre de individuos (Zimmerman et al., 1994; Navarro-Martínez, Durán-García, & Martínez-Gonzáles, 2012). Algunos autores como Macamo, Massuanganhe, Nicolau, Bandeira, y Adams (2016) consideran que estos eventos pueden crear condiciones extremas producto de la caída de árboles dentro del bosque, los cuales son sustituidos por sus mismos rebrotes o por regeneración de diferentes especies a lo largo del tiempo.
Las perturbaciones naturales y antropogénicas, como los huracanes y aprovechamientos forestales, juegan un papel importante en los cambios del entorno físico y la disponibilidad de recursos, ya que modifican poblaciones bióticas cruciales en el mantenimiento de procesos ecológicos y evolutivos de alta complejidad (Hogan, Zimmerman, Thompson, Nytch, & Uriarte, 2016). En los bosques en general, las tormentas y huracanes ocasionan cambios agudos en estructura reflejados a largo plazo (Long, Giri, Primavera, & Trivedi, 2016). Los patrones de producción de flores y semillas se ven fuertemente influenciados por las perturbaciones de los huracanes y las variaciones climáticas locales producidos por las oscilaciones globales que se generan a miles de kilómetros de distancia (Chang-Yang, Sun, Tsai, Lu, & Hsieh, 2015; Zimmerman, Hogan, Nytch, & Bithorn, 2018). Estudios realizados por Fernandes, Rollinson, Kearney, Dietze y Fagherazzi (2018) evidenciaron supresiones episódicas en el crecimiento radial de los árboles hasta por cuatro años después de pasado un evento extremo de tormenta, luego de este período el crecimiento comenzó a estabilizarse de nuevo.
El monitoreo de ecosistemas perturbados se ha convertido en un desafío para la ciencia, por lo que se han implementado técnicas de muestreos ecológicos y de teledetección para determinar los cambios en estructura e índices de vegetación asociados a las variaciones climáticas y los efectos de los fuertes vientos y lluvias (Jones, 2015; Ibharim, Mustapha, Lihan, & Mazlan, 2015; Duke et al., 2017). Hogan et al. (2016) publicaron que, además del efecto de los huracanes, existe un efecto agregado por las intervenciones antropogénicas que darán cambios en la resiliencia y secuencia de recuperación en los ecosistemas que ya están supuestamente adaptados a este tipo de disturbios.
A pesar de que se han realizado múltiples estudios en bosques afectados por huracanes en países como México, Belice, Puerto Rico y Guatemala (Zimmerman et al., 1994; Navarro-Martínez et al., 2012); para Costa Rica la información que existe se relaciona con fenómenos indirectos. Sin embargo, Otto es el primer registro oficial de impacto directo de un huracán en territorio nacional y se pronostica un aumento en la probabilidad de este tipo de eventos en el país (Sobel et al., 2016; Walsh et al., 2016; Wang et al., 2017); por lo que es necesario implementar nuevos trabajos que permitan ampliar el conocimiento acerca de los efectos de estos fenómenos en bosques tropicales y sus mecanismos de respuesta hacia la recuperación de la cobertura forestal. Consecuentemente, se vuelve necesario identificar la vulnerabilidad de los ecosistemas por las pérdidas que se generan a diferentes escalas, producto de los intensos eventos pluviales y los fuertes vientos (Queiroz et al., 2017). Este estudio tiene como objetivo general determinar el comportamiento y estado de la recuperación en un bosque secundario afectado directamente por el huracán Otto en Boca Tapada, Pital, San Carlos, Costa Rica. Como objetivos específicos (i) determinar la composición florística y estado de la masa remanente luego del paso del huracán; (ii) estimar los cambios en la cobertura forestal a causa de los efectos del huracán y (iii) comparar el estado actual del bosque secundario con el de un bosque natural aledaño que también fue afectado por el paso del huracán Otto en la Zona Norte de Costa Rica.
Materiales y métodos
Área de estudio: El estudio fue realizado entre enero y setiembre del 2019 en Boca Tapada de Pital, San Carlos, provincia de Alajuela, Costa Rica. Específicamente en un bosque secundario afectado por el huracán Otto en noviembre del 2016, propiedad del hotel Laguna del Lagarto Lodge y que se ubica en las coordenadas (10º41’27.924’’ N & 84º10’5.448’’ W). Este bosque tiene una edad aproximada de 25 años. El uso anterior del suelo fue la ganadería y se le realizó un Plan General de Manejo en el 2016 con un posterior aprovechamiento forestal meses previos al paso del huracán, donde se cortaron y extrajeron 24 árboles distribuidos a lo largo de las 21.67 ha que conforman el área de estudio.
Este bosque se encuentra ubicado según la clasificación de zonas de vida de Holdridge (1967) en la zona de vida bosque muy húmedo tropical (bmh-T). La precipitación media oscila entre los 3 000 y los 4 000 mm anuales, con una temperatura media de 27 °C y una topografía variable que alcanza hasta el 60 % de pendiente (Tapia, 2011). La estación seca tiene una duración de 2 meses en el período comprendido entre marzo y mayo (Alpizar, 2014). El tipo de suelo es clasificado dentro del gran grupo Tropohumult, y en el orden ultisol, con terrenos de origen volcánico y aluvial sedimentario (Mata, Rosales, Vásquez, & Sandoval, 2013).
Muestreo: Se realizó un muestreo sistemático, donde se estableció un carril madre o principal y perpendicular a este se crearon siete carriles secundarios paralelos, con un distanciamiento de 50 m entre cada uno de ellos. Dentro de los carriles secundarios se establecieron un total de 10 unidades de muestreo (UM), con dimensiones de 20 x 50 m, distribuidas de forma sistemática con una intensidad de muestreo del 4.6 %. Cada una de las UM fue dividida en 10 subparcelas de 100 m2.
Variables estudiadas: Dentro de las UM se muestrearon todos los individuos con diámetros iguales o superiores a los 5 cm a una altura de 1.30 m (d), se les identificó la familia y especie taxonómica, y se les midieron las variables altura total, forma de copa y se detalló la condición clasificándolos como, individuos en buen estado, a los que no presentaron un daño evidente por el paso del huracán, descopados, a los que presentaban evidencias de haber perdido la totalidad de su copa e inclinados, los que poseían inclinación de su fuste con respecto a su eje vertical original a causa de los fuertes vientos. Adicional a esto, dentro de la UM se determinaron las especies de los individuos volcados que presentaron rebrotes, y se consideraron como rebrotes aquellos tallos emergentes desde la base de individuos dañados y que poseían una altura superior a 1.30 metros, con un d mayor o igual a un centímetro; y como individuos volcados aquellos que perdieron completamente su verticalidad hasta desraizarse.
Se determinó el grado de recuperación de la copa luego de que los individuos fueron descopados. Para esto, se utilizó como base la metodología de Dawkins (1958), la cual establece las siguientes categorías:
• Perfecta, circulo perfecto, copas densas, simétricas, desarrolladas.
• Buena, más o menos simétricas, circulo irregular, algunas ramas ausentes.
• Tolerable, media copa, asimétrica, rala.
• Pobre, menos de media copa, muy asimétrica, pocas ramas con muerte regresiva extensa.
• Muy pobre, descopado, una o pocas ramas.
Cambio de cobertura: Se realizó un sobrevuelo a 100 m de altura con un vehículo aéreo no tripulado modelo DJI Phantom 4 Pro, con el cual se tomaron fotografías del área total del estudio con un traslape del 80 % y se creó una fotografía aérea ortorectificada. Con la ayuda del ortomosaico y el levantamiento topográfico obtenido durante el muestreo se identificaron diferentes áreas, las que presentaban evidencia de haber sido afectadas por el aprovechamiento forestal previo o por el paso del huracán Otto, caminos principales, lagunas, humedales y las zonas que conservaron la cobertura del bosque secundario en su condición inicial.
Con la información del plan general de manejo realizado en el bosque e informes de corta suministrados por la Comisión de Desarrollo Forestal de San Carlos (CODEFORSA) se ubicaron los árboles extraídos y se proyectaron sobre el ortomosaico. Luego se georreferenciaron las pistas de arrastre y se estimó el área afectada por la corta y extracción de los árboles. Con base en la georreferenciación de los árboles cortados, se estimó la proyección del área de impacto generada por la caída y se le sumó el área afectada por la creación de pistas de arrastre. Además, se partió de la premisa de que la condición inicial del bosque secundario mantenía el balance entre la caída normal de árboles, con claros pequeños que son rápidamente colonizados.
Se creó un archivo vectorial en formato shape para cada una de las condiciones del sitio de estudio, condición inicial, luego del aprovechamiento y condición actual con efectos del paso del huracán Otto. Finalmente, se utilizó el sistema de información geográfica y los archivos vectoriales debidamente clasificados se calculó el área para cada una de las coberturas en las diferentes condiciones evaluadas y con esto se determinó la diferencia de área para cada categoría al recibir las distintas perturbaciones.
Comparación con un bosque primario: La información recolectada en el muestreo fue comparada con información de un muestreo realizado en el 2017 en un bosque primario ubicado aproximadamente a 1 000 m del área de estudio; que sufrió el paso de un tornado local en el 2011 y posteriormente el impacto del huracán Otto en el 2016. En ambos casos hubo grandes daños a la masa forestal producto de los fuertes vientos, similares a los producidos en el bosque secundario del estudio.
En el bosque primario se realizó, al igual que en el bosque secundario, un muestreo sistemático, con la apertura de un transecto principal y otros secundarios perpendiculares cada 50 m, con un área total muestreada de tres hectáreas distribuidas en 10 parcelas de medición de 20 X 150 m y la medición de variables dasométricas en los individuos con diámetros (d) superiores a los 10 cm. Cabe resaltar, que debido a que el diámetro mínimo de medición fue diferente entre el bosque primario y el secundario, se omitieron en el análisis los individuos con d inferiores a los 10 cm, para homogeneizar los datos de ambos bosques y poder compararlos.
Se construyó la curva de acumulación de especies por área para cada uno de los bosques muestreados, además en ambos casos se calculó el estimador Chao 1 propuesto por Chao (1984), el cual está basado en la presencia de especies raras y es representado por la siguiente fórmula Sest = Sobs + F2 / 2G. Donde Sest corresponde al número de especies a estimar, Sobs es el número de especies observadas, F es el número de especies representadas por un solo individuo dentro de la muestra y G es el número de especies representadas por exactamente dos individuos.
El análisis de variables dasométricas, las presentaciones de cuadros resumen y gráficos se realizaron con el software Microsoft® Excel 2013. El procesamiento de las imágenes aéreas se llevó a cabo en el software Agisoft PhotoScan 1.4.5. La digitalización de los archivos vectoriales y la estimación en los cambios de área de cobertura se realizaron con el sistema de información geográfica Qgis 3.4.8., mientras que para la estimación de las curvas de acumulación de especies se utilizó el software EstimateS 9.1.0.
Adicional, se seleccionaron 99 subparcelas de 100 m2 que fueron establecidas en el muestreo del bosque secundario y se compararon con 99 subparcelas de 100 m2 del bosque primario para probar la hipótesis de la existencia de diferencias significativas en el número de individuos, área basal y número de especies encontradas entre los dos tipos de bosque. Debido a que los datos, en el software estadístico InFostat (versión 2019; Di Rienzo et al., 2016) no cumplieron con los supuestos de normalidad de residuos con la prueba Shapiro-Wilks, ni con la prueba de homocedasticidad de residuos absolutos, se aplicó estadística no paramétrica, con la prueba de Wilcoxon (Mann-Whitney U), con una confianza del 95 % (α = 0.05).
Resultados
Estado actual del bosque: En el muestreo del bosque secundario se registró un promedio de 1 052 individuos por hectárea con d superior a los 5 cm, de los cuales 731 individuos no presentan evidencia de afectación por el huracán, 214 individuos fueron descopados, mientras que 128 quedaron inclinados, lo que representa que un 69.5 % de los individuos se encuentra en buen estado, un 20.3 % presentó descope y un 12.17 % estaban inclinados. Un 2 % de los individuos mostraron ambas condiciones de daño, descope e inclinación. En cuanto al área basal, se determinó que el bosque posee un valor medio de 20.54 m2/ha, del cual un 21 % (4.32 m2/ha) es aportada por individuos descopados y un 7.16 % (1.47 m2/ha) por individuos inclinados.
Se registraron un total de 171 especies distribuidas en 51 familias taxonómicas, Fabaceae fue la que más riqueza aportó con un total de 24 especies, seguido por Sapotaceae con 15 especies, Lauraceae con 11, Rubiaceae con 8, Annonaceae, Euphorbiaceae y Myrtaceae con 7 especies cada una, y Moraceae con 6 especies; entre estas 8 familias mencionadas el 51 % del total de las especies encontradas, mientras que 23 de las familias restantes solo contribuyen con una especie cada una, lo que equivale al 13.4 % del total de familias.
En la Fig. 1 se muestra la cantidad de especies por familia que presentaron individuos descopados por causa del paso del huracán Otto. En total se contabilizaron individuos dañados pertenecientes a 35 familias y un total de 85 especies, donde la familia que más especies afectadas presentó fue Fabaceae con 12 especies, seguido por Sapotaceae con 5 y Salicaceae, Myristicaceae, Malvaceae y Burseraceae con 4 especies cada una. Sin embargo, cabe mencionar que la especie que más individuos descopados presentó fue Couma macrocarpa de la familia Euphorbiaceae, seguido por Laetia procera (Salicaceae), Vochysia ferruginea (Vochysiaceae), Casearia arborea (Salicaceae) y Virola koschnyii (Myristicaceae); estas suman entre todas más de un 25 % del total de los individuos dañados por descope.
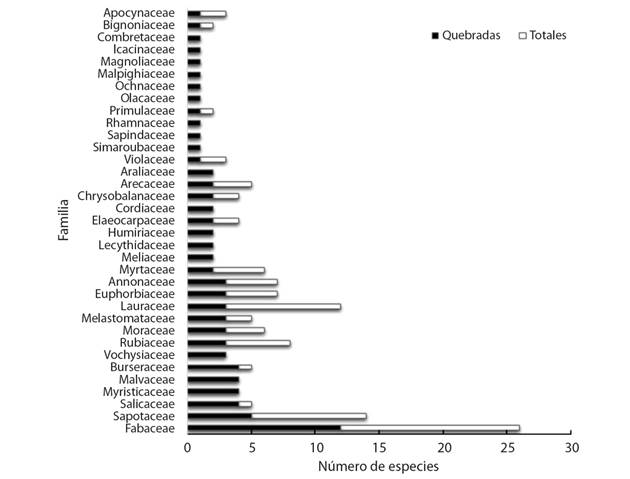
Fig. 1 Cantidad de especies dañadas por descope de acuerdo a cada familia taxonómica en un bosque secundario de Boca Tapada, Pital, San Carlos, Costa Rica. / Fig. 1. Number of species damaged by tree top loss according to each taxonomic family in a secondary forest of Boca Tapada, Pital, San Carlos, Costa Rica.
En la Fig. 2 se muestra el total de especies por familia taxonómica cuyos individuos presentaron evidencia de haberse inclinado como consecuencia del paso del huracán Otto. Se registraron un total de 29 familias afectadas, donde nuevamente la que presentó más especies dañadas fue Fabaceae con 7 especies, seguido por Rubiaceae con 5, Melastomataceae, Annonaceae y Sapotaceae con 4 especies cada una. La especie que mayor cantidad de individuos inclinados presentó fue Couma macrocarpa, seguida por Casearia arborea, Qualea polychroma y Ferdinandusa panamensis; lo que representa un 25 % del total de los individuos inclinados.
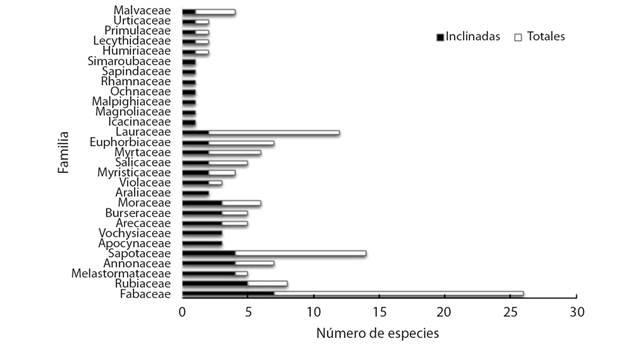
Fig. 2 Cantidad de especies inclinadas por el paso del huracán Otto de acuerdo a cada familia taxonómica en un bosque secundario de Boca Tapada, Pital, San Carlos, Costa Rica. / Fig. 2. Number of species inclined by the passage of Hurricane Otto according to each taxonomic family in a secondary forest of Boca Tapada, Pital, San Carlos, Costa Rica.
Recuperación del bosque: Se identificaron las especies cuyos individuos volcados produjeron rebrotes, las cuales se detallan en la Tabla 1. Se contabilizaron un total de 136 individuos por hectárea con rebrote, distribuidos en 36 especies y 20 familias; la familia Fabaceae fue la que más especies con capacidad de rebrote registró con 6 especies, seguido por Euphorbiaceae, Lauraceae, Sapotaceae y Vochysiaceae con 3 especies cada una. Entre estas cinco familias se acumuló el 50 % de las especies registradas. La especie con mayor número de individuos rebrotados fue Inga cooclensis, seguido por Cupania scrobiculata, Alchornea latifolia y Cespedecia spathulata; entre ellas acumulan un 37.5 % del total de individuos con rebrote.
Tabla 1 Familias taxonómicas y especies registradas con presencia de rebrotes en un bosque secundario de Boca Tapada, Pital, San Carlos, Costa Rica. / TABLE 1. Taxonomic families and registered species with resprouting in a secondary forest of Boca Tapada, Pital, San Carlos, Costa Rica
| Familia | Especie |
| Araliaceae | Dendropanax arboreus |
| Burseraceae | Protium glabrum |
| Chrysobalanaceae | Hirtella media |
| Combretaceae | Terminalia amazonia |
| Elaeocarpaceae | Sloanea tuerckheimii |
| Euphorbiaceae | Alchornea latifolia |
| Croton schiedeanus | |
| Mabea occidentalis | |
| Fabaceae | Dialium guianense |
| Dussia macroprophyllata | |
| Enterolobium schomburgkii | |
| Inga cocleensis | |
| Inga thibaudiana | |
| Vatairea erythrocarpa | |
| Hernandiaceae | Hernandia didymantha |
| Humiriaceae | Vantanea occidentalis |
| Hypericaceae | Vismia macrophylla |
| Lauraceae | Licaria sarapiquensis |
| Nectandra umbrosa | |
| Ocotea dendrodaphne | |
| Malvaceae | Apeiba membranacea |
| Myristicaceae | Virola multiflora |
| Virola sebifera | |
| Ochnaceae | Cespedesia spathulata |
| Olacaceae | Minquartia guianensis |
| Rubiaceae | Ferdinandusa panamensis |
| Warszewiczia coccinea | |
| Salicaceae | Casearia arborea |
| Laetia procera | |
| Sapindaceae | Cupania scrobiculata |
| Sapotaceae | Elaeoluma glabrescens |
| Pouteria campechiana | |
| Sideroxylum contrerasii | |
| Vochysiaceae | Qualea polychroma |
| Vochysia alleni | |
| Vochysia ferruginea |
En la Fig. 3 se puede observar la curva de distribución diamétrica de los individuos por hectárea que fueron dañados por descope, así como las curvas poblacionales de acuerdo al grado de recuperación de los mismos luego de que fueran dañados por el paso del huracán Otto. El grado de recuperación se estimó basándose en la forma de copa.
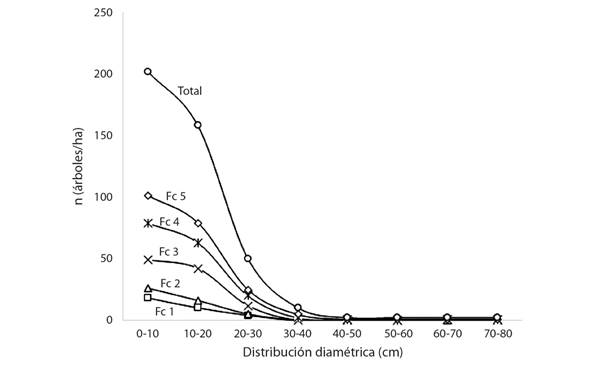
Fig. 3 Individuos por distribución diamétrica en cada una de las clases de formas de copa (Fc) en un bosque secundario de Boca Tapada, Pital, San Carlos, Costa Rica. / Fig. 3. Individuals by diametric distribution in each of the classes of tree top (Fc) in a secondary forest of Boca Tapada, Pital, San Carlos, Costa Rica.
En la curva de individuos totales se muestra con claridad que la mayor cantidad de individuos dañados se encuentra en la categoría de 10 a 20 centímetros de d y que esta cantidad disminuye gradualmente conforme aumenta la categoría diamétrica, lo que indica que individuos de diámetros inferiores tienden a quebrarse más. A partir de la clase de 40 a 50 cm de d hasta la clase máxima se presentó solo un individuo dañado por clase. Además, si se observan las curvas de distribución de acuerdo a la forma de copa, se puede notar que todos los individuos que logran reponer su copa de la forma 3 o tolerable hasta la forma de copa 1 o copa perfecta, pertenecen a clases diamétricas inferiores a los 30 cm, mientras que los individuos con valores de diámetro superiores a esto poseen copas clasificadas en la forma 4 y 5, lo que corresponde a copas pobres y muy pobres respectivamente. Ninguno de los individuos con estos diámetros superiores logró recuperar su copa hasta llegar a los valores de forma de copa óptimos. Este comportamiento sugiere que los individuos más jóvenes son los que poseen una mayor capacidad de recuperación luego de ser descopados por el viento.
Cambio de la cobertura forestal: Con la ayuda del levantamiento forestal realizado, los datos del inventario previo al paso del huracán y los informes de corta del aprovechamiento forestal, se clasificaron las áreas para cada una de las coberturas en los tres diferentes tiempos evaluados (Fig. 4). El área correspondiente a caminos principales, lagunas y humedales se mantuvo fijo durante el tiempo de estudio. El bosque secundario en su condición inicial poseía un área aproximada de 20.74 ha, luego de la ejecución del aprovechamiento forestal producto de las labores de corta y extracción de los árboles esa área se redujo a 19.15 ha lo que representa una reducción del 7.63 % de la cobertura del bosque secundario, para posteriormente reducirse a 10.68 ha, con un porcentaje de daño del 44.25 % que corresponde a un total de 8.48 ha de bosque afectadas por el paso del huracán Otto. La reducción del área de bosque secundario desde el momento inicial hasta el paso del huracán fue del 59.1 % equivalente a una afectación de 12.3 ha de la cobertura forestal inicial.
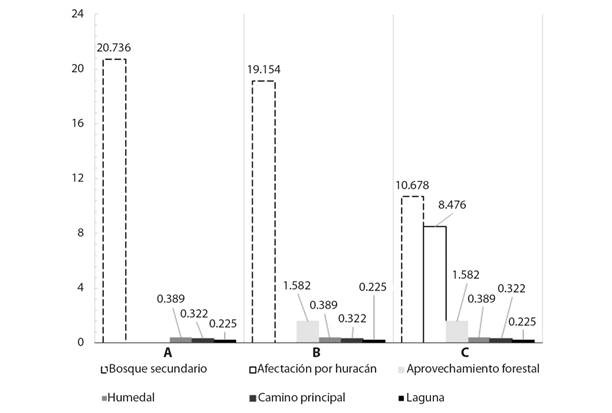
Fig. 4 Hectáreas de cobertura en el sitio de estudio. A. Condición inicial. B. Luego de la ejecución de un aprovechamiento forestal. C. Condición actual luego del paso del huracán Otto en un bosque secundario de Boca Tapada, Pital, San Carlos, Costa Rica. / Fig. 4. Hectares of coverage at the study site. A. Initial condition. B. After the execution of a forest logging. C. Current condition after the Hurricane Otto in a secondary forest of Boca Tapada, Pital, San Carlos, Costa Rica.
Comparación entre tipos de bosque: En la Fig. 5 se presentan las curvas de acumulación de especies reales obtenidas en los muestreos y las calculadas basándose en el estimador no paramétrico Chao 1, el cual se basa en la abundancia de individuos dentro de cada especie y en la frecuencia con que estos aparecen conforme se aumenta el tamaño del área de muestreo. En este caso, se puede observar que la curva del bosque secundario (BS) sugiere la presencia de una mayor cantidad de especies con respecto al bosque primario (BP), tanto para los valores reales obtenidos en los muestreos como para los calculados con el estimador Chao 1. Además, en las curvas estimadas se puede visualizar como estas se intersecan una vez alcanzados los 8 000 m2 de área, de este punto en adelante la curva de BP se vuelve superior a la del BS.
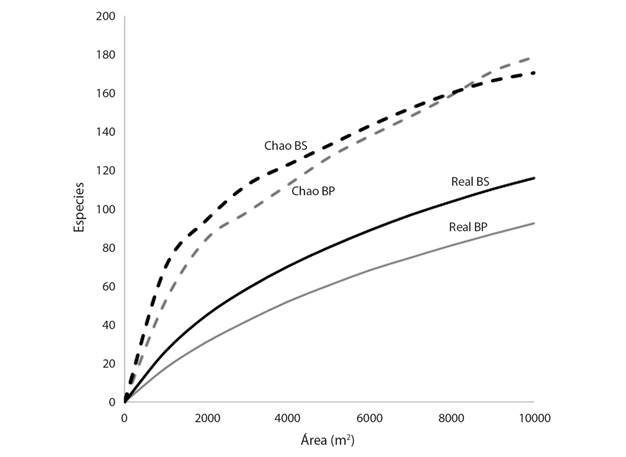
Fig. 5 Especies acumuladas por área, observadas y estimadas para un bosque secundario (BS) y un bosque primario (BP) en Boca Tapada, Pital, San Carlos, Costa Rica. / Fig. 5. Cumulative species by area observed and estimated for a secondary forest (BS) and a primary forest (BP) in Boca Tapada, Pital, San Carlos, Costa Rica.
Al realizar la prueba de igualdad de varianzas de Wilcoxon (Mann-Whitney U) se determinó que existe diferencia significativa entre la cantidad de individuos (P = 0.0011; α = 0.05), y entre la cantidad de especies encontradas en subunidades de muestreo de 100 m2 (P = 0.0004; α = 0.05). En ambos casos el bosque secundario posee los valores medios más altos. En el caso del área basal, no se muestran diferencias significativas entre los dos tipos de bosque comparados (P = 0.6951; α = 0.05).
Discusión
Los mayores efectos observados por el paso del huracán Otto están dados, se da en las alteraciones en la abundancia y arquitectura de los individuos arbóreos (Bellingham, Tanner, & Healey, 1995; De Gouvenain & Silander, 2003; Vandecar et al., 2011). Los valores de densidad poblacional obtenidos durante el muestro fueron inferiores a los mencionados en otros bosques, como es el caso de bosques húmedos tropicales afectados por el huracán Dean en México, donde se presentaron entre 1 400 y 1 450 individuos por hectárea para diámetros superiores a los 5 cm (McGroddy et al., 2013, Pat-Aké et al., 2018). Estos valores corresponden a bosques maduros, por lo que es razonable que sean superiores, ya que estos bosques son una mezcla de especies con diferentes grados de tolerancia lumínica (Quesada-Monge, Acosta-Vargas, Garro-Chavarría, & Castilla-Ugalde, 2012), lo que potencia la densidad de individuos en el sotobosque que sobrepasen el diámetro mínimo de medición. A diferencia de esto, el bosque evaluado en este estudio es un bosque secundario, donde debido al bajo grado de desarrollo del bosque la presencia de individuos de especies intolerantes a la luz es menor (Guariguata & Ostertag, 2002). Esto sumado a que en las áreas perturbadas el grado de disturbio fue alto, lo que genera una gran cantidad de materia orgánica acumulada producto de la defoliación, descope y desraizado de árboles, así como la competencia alta y constante generada por algunas especies de bejucos y enredaderas que son más exitosas en colonizar claros, como la Scleria spp. (Cyperaceae), que impiden que le llegue la suficiente luz a individuos de especies arbóreas provenientes del banco de semillas e inmigrantes por agentes dispersores para que logren reclutarse en las áreas perturbadas (Vandermeer, de la Cerda, Boucher, Perfecto, & Ruiz, 2000; Silver, Hall, & González, 2014).
Se debe considerar que las evaluaciones mencionadas en la literatura fueron realizadas en periodos menores a un año luego de sucedido el evento, y como lo menciona Whigham, Olmsted, Cabrera-Cano, y Harmon (1991), Imbert, Labbé, y Rousteau (1996), e Imbert y Portecop (2008), hay una alta posibilidad de mortalidad de individuos con daños severos en los 2 años posteriores a la perturbación.
El valor de 20.3 % de árboles descopados obtenido en el presente estudio es muy similar con el 20.6 % encontrado por Vandecar et al. (2011), para bosques secundarios intermedios de la península de Yucatán. Estos valores también son compatibles con el 22 % mencionado por Navarro-Martínez et al. (2012) y el 18 % descrito por Pat-Aké et al. (2018) para bosques húmedos tropicales con sucesiones más avanzadas. Además, al igual que en otros estudios, la familia Fabaceae es la que aporta la mayor riqueza de especies y la que mayor abundancia de individuos dañados posee (Haas-Ek, Gonzáles-Valvidia, De Jong, Ochoa-Goana, & Aryal, 2019).
Se identificó a Couma macrocarpa (Euphorbiaceae) como la especie con mayor abundancia de individuos dañados, esto se puede deber a que es una especie con una densidad de madera de 0.47 g/cm3 (Zanne et al., 2009), considerada una densidad media. Además, esta especie alcanza los niveles más altos del dosel, y crea una copa densa con abundante follaje y foliolos de gran tamaño. Estos aspectos mencionados son claves en el aumento de la susceptibilidad al daño por viento en una especie (Boose, Serrano, & Foster, 2004; Webb et al., 2013).
La familia Fabaceae ha sido ampliamente documentada como forrajera por su alta capacidad de rebrote luego de ser sometida a fuego o daños mecánicos (Chan-Dzul, 2010; Haas-Ek et al., 2019), lo que justifica que sea la familia que aporta el mayor número de especies y la mayor abundancia de rebrotes dentro del bosque. Algunos autores mencionan que en los bosques tropicales más del 70 % de las especies tienen la capacidad de rebrotar y que esta característica posee gran relevancia en la dinámica de recuperación del bosque luego del paso de un huracán (Boucher et al., 2001; Islebe, Torrescano-Valle, Valdez_Hernández, Tuz-Novelo, & Weissenberger, 2009). Además, desde el punto de vista ecológico esta familia es muy importante en la recuperación del bosque, ya que posee una gran capacidad en la fijación de nitrógeno (López-Pérez & Boronat-Gil, 2016), sus flores y frutos resultan ser muy atractivas para polinizadores y dispersores, que aumentan la diversidad en fauna y la posibilidad de que estos dispersores aporten semillas de especies provenientes de otros sitios (Meli & Carrasco-Carballido, 2011).
Se encontró una mayor tendencia al daño por descope e inclinación en los individuos con d menores a los 40 cm, lo que concuerda con Putz y Sharitz (1991), Frangi y Lugo (1991), y Navarro-Martínez et al. (2012) quienes mencionan que en bosques secundarios o con perturbaciones antropogénicas previas los individuos jóvenes y de diámetros inferiores a los 30 centímetros poseen una mayor susceptibilidad a recibir un daño severo en comparación con individuos de diámetros superiores. Además, se encontró que individuos de diámetros inferiores a los 30 cm poseen una mayor capacidad de recuperación de la forma de copa luego de haber sido descopados, debido a un mayor potencial de rebrote presente en individuos más jóvenes (Haas-Ek et al., 2019)
Al analizar el cambio de cobertura se estimó que el huracán provocó fuertes daños en aproximadamente un 44.25 % de la cobertura inicial, valor superior a lo mencionado por Xi (2015) quien habla de valores de daño cercanos al 24 % de cobertura en un bosque maduro. Sin embargo, concuerda con Pat-Aké et al. (2018) que obtuvo valores cercanos al 50 % para bosques que fueron aprovechados previo al paso del huracán Dean. Según Everham y Brokaw (1996), Garrido et al. (2008), y Zeng, Peltola, Venäläinen, y Kellomäki (2009) los bosques intervenidos por aprovechamientos forestales se vuelven más susceptibles al viento a causa de la homogeneización de la altura del dosel y el aumento de riesgo de caída en los árboles de borde en los claros generados (Young & Hubbell, 1991).
Ferrando, Louman, Finegan y Guariguata (2001) y Heinonen et al. (2009) sugieren que, en bosques que se encuentran en zonas donde los huracanes son frecuentes o algunos eventos extremos que produzcan fuertes vientos; se deben planificar los aprovechamientos forestales de forma estratificada e identificar las áreas vulnerables y buscar la concentración y reducción de los claros producidos.
Al realizar la proyección de las curvas de acumulación de especies, tanto real como la calculada con el estimador Chao 1, se puede observar que el bosque secundario evaluado posee una mayor cantidad de especies acumulada por unidad de área. Además, la prueba de igual de varianzas de Kruskal-Wallis sugiere que existen diferencias significativas (P = 0.0004; α = 0.05) en el número de especies encontrado por unidad muestreal. El BS es el que posee la mayor media de especies acumuladas. Múltiples autores han encontrado en el estudio de cronosecuencias de bosques secundarios que la diversidad florística aumenta en función de la edad del bosque y el avance la sucesión (Guariguata, Chazdon, Denslow, & Dupuy, 1997, Ferreira, Finegan, Kanninen, Delgado, & Segura, 2002, Vílchez, Chazdon, & Milla, 2008). El bosque primario estudiado recibió dos grandes perturbaciones en un periodo de tiempo relativamente corto (Araya-Orozco, 2017), lo que generó grandes áreas de claros en el bosque, colonizadas por unas pocas especies pioneras y con la apertura del dosel superior se da el aumento del grado lumínico que atraviesa hasta el sotobosque (Guariguata & Ostertag, 2002), lo que puede ocasionar que muchas especies intolerantes a la luz sean suprimidas por el cambio abrupto en sus condiciones. Estos factores contribuyeron para que gran proporción de este bosque se encuentre en una fase dinámica de silvigénesis que asemeja la composición de un bosque secundario en etapas iniciales con la consecuente baja en la diversidad del bosque (Brokaw, 1985).
En el caso de la densidad poblacional, existen diferencias significativas (P = 0.0011; α = 0.05). El bosque secundario presentó la mayor media en la densidad de especies. Este comportamiento ya ha sido antes descrito por (Morales-Salazar et al., 2012) quienes mencionaron que en bosques secundarios con trayectorias de regeneración cercana a los 30 años y que alcanzan una sucesión intermedia-avanzada en la Península de Osa en Costa Rica, poseen una mayor densidad poblacional con respecto a los bosques primarios. La tendencia habitual para los bosques secundarios húmedos, es que en fases dinámicas la cantidad de individuos aumente conforme aumenta la sucesión hasta llegar a un estadio avanzado, donde la cantidad de individuos decrece y el bosque secundario llega a un estado homoestático de silvigénesis donde asemeja al bosque no intervenido (Guariguata et al., 1997; Ruschel, Mantovani, Sedrez, & Onofre, 2009).
El presente trabajo demostró que un 69.5 % de la masa remanente se encuentra en buen estado luego del paso del huracán Otto, mientras que un 20.3 % presentó daños por descope y un 12.7 % sufrió de inclinación sin exposición de raíces. Las siete familias más importantes fueron: Fabaceae con 24 especies, Sapotaceae con 15, Lauraceae con 11, Rubiaceae con 8, y Annonaceae, Euphorbiaceae y Myrtaceae con 7 especies cada una.
En el análisis de la composición florística se encontró que la familia Fabaceae es la que mayor riqueza de especies aporta, más abundancia de individuos dañados posee y la que tuvo mayor cantidad de especies con capacidad de rebrote, Couma macrocarpa es la especie que más se dañó a causa del efecto del huracán y las especies con mayor número de individuos rebrotados fueron Inga cooclensis, Cupania scrobiculata, Alchornea latifolia y Cespedecia spathulata.
El análisis de la masa remanente mostró que después del aprovechamiento forestal se redujo un 7.63 % de la cobertura forestal con respecto a la condición inicial del bosque secundario, mientras que el paso del huracán causó un impacto del 44.5 %. Además, el aprovechamiento forestal en un bosque puede aumentar el riesgo de afectación por el paso de huracanes.
El estado actual del sitio de estudio muestra que el bosque secundario posee una mayor diversidad de especies y una mayor densidad de individuos que el bosque primario.
Declaración: los autores declaran que todos están de acuerdo con este artículo y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes para garantizar su publicación.

