











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
El banco de semillas del suelo es uno de los indicadores ecológicos más importantes para la evaluación y monitoreo de recuperación de comunidades vegetales en diferentes tipos de ecosistemas (Martins, Almeida, Fernandes, & Ribeiro, 2008; Martins, 2009; Chazdon, 2014; Mall & Singh, 2014). El banco de semillas del suelo consiste en la acumulación de propágulos viables que provienen de comunidades de plantas de áreas adyacentes (Chazdon, 2014; Mall & Singh 2014; Martins, Borges, & Silva, 2015). Las semillas permanecen latentes en el suelo por un tiempo determinado sin perder su viabilidad, hasta que ocurre algún tipo disturbio que induce su germinación, dando inicio a la regeneración natural (Martins, 2009; Chazdon, 2014; Mall & Singh, 2014; Kildisheva et al., 2020). Existen varios atributos de los bancos de semillas que permiten evaluar su calidad y potencial para la regeneración natural o restauración activa de los ecosistemas (ej. transposición de banco), como por ejemplo, densidad de semillas viables, riqueza y composición de especies (Martins et al., 2015; Martins, 2018; Silva, Martins, Miranda-Neto, & Texeira, 2019). Sin embargo, estos atributos dependen de diferentes factores como la distancia de las fuentes de propágulos, presencia de dispersores, e intensidad en el uso de la tierra (Dalling, Swaine, & Garwood, 1997; Martins & Engel, 2007; Martins, 2018).
Después de la dispersión, algunas semillas quedan retenidas en la hojarasca, mientras que otras son depositadas a diferentes profundidades debido a sus características morfológicas (Baider, Tabarelli, & Mantovani, 2001; Menezes, Neto, Azevedo, Machado, & Nunes, 2019). Las semillas se pueden depositar a varias profundidades, dependiendo de la cantidad, tamaño y peso de las semillas, lo cual determina su período de viabilidad y capacidad de germinación (Menezes et al., 2019). La mayoría de los estudios indican una mayor densidad de semillas en las capas más superficiales del suelo donde prevalece un constante reemplazo de semillas (Baider et al., 2001; Martins et al., 2015). Así mismo, la mayor germinación de semillas ocurre en la capa más superficial del suelo, que puede estar fuertemente influenciada por las condiciones de luz y temperatura (Baider et al., 2001). Sin embargo, pocos estudios han demostrado estas variaciones de riqueza y composición de especies entre profundidades del banco de semillas.
El banco de semillas de bosques tropicales está compuesto principalmente por especies pioneras exigentes de luz, responsables de las primeras etapas de regeneración natural (Dalling et al., 1997; Martins, 2009; Miranda-Neto & Martins, 2017; Bechara et al., 2020). Sin embargo, la calidad y potencial del banco de semillas después de un disturbio es determinada por una alta riqueza de especies y variabilidad de la composición florística (Silva-Weber, Nogueira, Carpanezzi, Galvão, & Weber, 2012; Balestrin, Martins, Schoorl, Lopes, & Andrade, 2019). Por lo tanto, una alta densidad y riqueza de especies pioneras y tolerantes a la sombra de etapas avanzadas de sucesión en el mismo banco de semillas, es un indicador de alta calidad (Silva-Weber et al., 2012; Balestrin et al., 2019). Por esta razón, el banco de semillas ha sido considerado un importante indicador ecológico de degradación ambiental en diferentes escenarios de cambios en el uso de la tierra y disturbios antropogénicos, como por ejemplo la agricultura y minería como principales actividades de relevancia económica que afectan negativamente los ecosistemas (ej. Martins et al., 2008; Mall & Singh, 2014; Miranda-Neto & Martins, 2017). Así mismo, el banco de semillas permite evaluar el potencial de regeneración de áreas degradadas (Martins, 2018; Silva et al., 2019). Por otro lado, los ecosistemas de referencia sin disturbios pueden ser fuentes para la transposición de banco de semillas y nucleación durante la restauración activa (Martins, 2018; Silva et al., 2019). La nucleación es una técnica de restauración activa que consiste en el establecimiento y manejo de núcleos de plantas en áreas degradadas a partir de la transposición de bancos de semillas de ecosistemas conservados para promover la recuperación natural (Martins, 2018).
La minería es una de las principales actividades económicas de Brasil por parte del sector privado, que comprometen mayores esfuerzos e inversiones en actividades de mitigación ambiental y restauración ecológica (Silva et al., 2019; Martins, Villa, Balestrin, Nabeta, & Silva, 2020). Por este motivo, las compañías mineras son responsables de la restauración de las áreas afectadas como procedimiento legal en Brasil (Martins et al., 2020). En este contexto, cabe resaltar que recientemente algunas áreas fueron afectadas por el impacto de los relaves mineros después de la ruptura de la represa de Fundão en Mariana, Minas Gerais, Brasil (Martins et al., 2020). Los relaves un conjunto de desechos de procesos mineros usualmente constituido por una mezcla de rocas molidas, agua y minerales (Campanharo et al., 2021). Por esta razón, las actividades de restauración ecológica han recibido alta prioridad en estas áreas afectadas mediante la aplicación combinada de diferentes métodos y técnicas de restauración activa y pasiva (Martins et al., 2020). Sin embargo, todavía existe la urgente necesidad de evaluar diferentes indicadores ecológicos que permitan identificar niveles de recuperación y ajustar métodos de restauración aplicados, como por ejemplo el monitoreo de bancos de semillas.
En este estudio se evaluó la variación de la riqueza, composición de especies, abundancia y parámetros fitosociológicos del banco de semillas a dos profundidades de relaves mineros en Mariana, municipio de Mariana, Minas Gerais, Brasil. Los principales parámetros fitosociológicos, fueron el número de individuos de todas las especies, frecuencia absoluta, frecuencia relativa, densidad absoluta y densidad relativa; así como grupos funcionales (estrategias de regeneración y síndromes de dispersión). Se tiene como premisa que este estudio es distinto de los trabajos tradicionales sobre el banco de semillas del suelo al comparar dos profundidades de relaves mineros. Por lo tanto, se establece como hipótesis que la profundidad del relave minero determina cambios en la distribución de la diversidad y estructura del banco de semillas debido a la diferenciación de grupos funcionales (estrategias de regeneración y síndromes dispersión). Específicamente se presume que existe una marcada variabilidad de la calidad del banco de semillas en términos de riqueza de especies, composición florística, abundancia y densidad de semillas entre profundidades del relave minero, a pesar de la poca diferencia de profundidad entre capas superficiales de relave (entre 0-2.5 cm, y entre 2.5-5 cm). Como predicciones establecimos que el banco de semillas de la capa superior P1 (0-2.5 cm) está dominado por especies pioneras y anemocóricas, e influenciado por la lluvia de semillas reciente desde los bosques cercanos. Por el contrario, el banco de semillas de la capa inferior P2 (2.5-5 cm) puede presentar menor riqueza de especies y densidad de semillas viables, como resultado de relaves mineros acumulados.
Materiales y métodos
Descripción del área de estudio: Descripción del área de estudio: El estudio se llevó a cabo entre julio y agosto de 2018, en un área afectada por los relaves mineros resultantes de la ruptura de la represa de Fundão en el 2015 (Fig. 1A) en Brasil (Fig. 1B), Minas Gerais (Fig. 1C), específicamente en el municipio de Mariana (23º02’70”-23º02’85” N & 48º28’59”-48º28’63” W, Fig. 1D). El área de estudio se encuentra dentro de los límites de la Área Ambiental I de actuación de la Fundación Renova. Esta área corresponde a la zona de Minas Gerais, conocida como Quadrilátero Ferrífero dentro del bioma del Bosque Atlántico, con una altitud promedio de 712 metros. El clima de la región, según la clasificación de Köppen, es del tipo Cwa, con una precipitación promedio de 1 307 mm y una temperatura promedio de 20.4 °C (Martins et al., 2020).
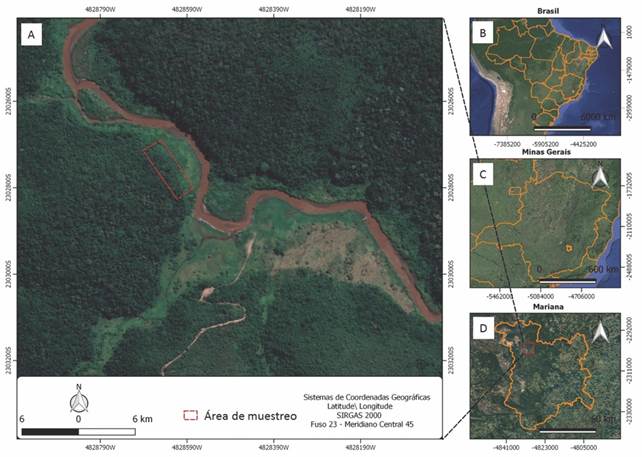
Fig. 1 Área de estudio en el río Gualaxo relación con A. América del Sur y B. Brasil, C. Minas Gerais y, D. Mariana. / Fig. 1. Localization of the study area in the Gualaxo river according to A. South America B. Brasil, C. Minas Gerais and, D. Mariana.
Los relaves acumulados en el área de estudio presentan diferentes profundidades (ca 80-100 cm) sobre una topografía plana y homogénea a lo largo del río. La vegetación ribereña a lo largo del río Gualaxo do Norte se clasifica como bosque estacional semidecidual. Asimismo, cabe mencionar que el área de estudio tenía una larga historia de uso de suelo a base de pasto para el ganado antes de la acumulación de relaves mineros por el colapso de la presa de Fundão (Campanharo et al., 2020; Campanharo et al., 2021).
Muestreo del banco de semillas: Se establecieron 15 parcelas de 2 × 2 m en área con relaves mineros en enero de 2019, completando un poco más de tres años después de la ruptura de la represa. Las parcelas fueron distribuidas bajo un diseño sistemático a lo largo de una hilera de 70 m con 5 m de separación en el área de relaves acumulados localizada entre dos fragmentos forestales separados por 250 m de distancia (Fig. 1). El banco de semillas fue muestreado a dos profundidades diferentes, una capa superior (P1, hasta 2.5 cm) y una capa inferior (P2, 2.5-5.0 cm) del relave acumulado. Para recolectar el banco de semillas se usó un marco de tubería de PVC de 40 × 40 cm, que se arrojó aleatoriamente dos veces dentro de cada parcela, donde posteriormente fueron colectadas muestras de relaves para cada profundidad. Por lo tanto, se recolectó un total de 30 muestras de banco de semillas para cada profundidad.
Las 60 muestras de relaves se colocaron en bolsas de plástico transparentes debidamente etiquetadas y se enviaron para un vivero de la Universidad Federal de Viçosa, Viçosa, Minas Gerais. Las muestras fueron colocadas en bandejas plásticas de 0.25 × 0.30 × 0.05 m con orificios de drenaje y dispuestas sobre mesones de concreto de 1 m de altura en invernadero con un 50 % de tela de sombra para evitar la contaminación externa. Las muestras de relave se sometieron a riego por aspersión programado (cuatro riegos diarios de tres minutos de duración) durante seis meses. El banco de semillas del suelo se evaluó a lo largo de este período con base en el método de emergencia de plántulas (ej. Silva et al., 2019). Las plántulas emergentes se contaron e identificaron una vez cada dos semanas e inmediatamente fueron retiradas de las bandejas. Para corroborar que el banco de semillas no esté contaminado, se colocaron dos recipientes con arena gruesa esterilizada como un control, lo que permitirá registrar la germinación sólo de las semillas colectadas en el área de estudio (ej. Silva et al., 2019).
La identificación taxonómica de plantas germinadas se realizó de acuerdo con el sistema del Grupo de Filogenia Angiosperma (APG IV). Los nombres de las especies se verificaron utilizando la base de datos en el sitio web del Jardín Botánico de Missouri. Las plantas se identificaron a nivel de familia y especie. Las especies encontradas en los bancos de semillas se clasificaron, según la categoría sucesional o estrategia de regeneración, como especies: pioneras, secundarias iniciales, secundarias tardías. También se realizó una clasificación con respecto a las formas de vida (herbáceas, arbustos y árboles) y el síndrome de dispersión (autocóricas, anemocóricas y zoocóricas), además del origen de las especies (exóticas y nativas).
Análisis de datos: Para comparar la riqueza de especies entre los bancos de semillas de las dos profundidades se estimaron curvas de rarefacción y extrapolación basadas en el número de parcelas con el primer número de Hill, q = 0 (Chao et al., 2014). Se hicieron extrapolaciones basadas en la presencia y ausencia de especies en las parcelas (Colwell et al., 2012). La rarefacción se estimó como la media de 100 repeticiones de bootstrapping para estimar intervalos de confianza del 95 %. Cuando los intervalos de confianza del 95 % no se superponen, significa que los números de especies diferían significativamente en P < 0.05 (Colwell et al., 2012). Estas estimaciones se obtuvieron utilizando el paquete “iNEXT” (Hsieh, Ma, & Chao, 2016) del software estadístico R (R Core Team, 2018). Para comparar la densidad de individuos de especies (datos no distribuidos normalmente) entre profundidades se usó la prueba de Wilcoxon para muestras pareadas (P < 0.05). Para esta variable, se probó la distribución normal con la prueba de Shapiro-Wilk, evaluando la gráfica Q-Q, y la homogeneidad de varianza mediante la prueba de Bartlett usando el programa R (R Core Team, 2018).
Realizamos un análisis de coordenadas principales (PCoA), basado en la matriz de disimilitud de Bray-Curtis para comparar los efectos de la profundidad sobre los patrones de composición de especies utilizando el paquete “vegan” en el programa R (R Core Team, 2018). Así mismo, utilizamos el análisis de varianza multivariado con permutaciones (PERMANOVA, 9 999 permutaciones) para determinar las posibles diferencias en la composición de especies utilizando la rutina ‘adonis’, disponible en el mismo paquete “vegan” (R Core Team, 2018).
A partir del número de individuos de todas las especies (NI), se estimaron la frecuencia absoluta (FA), la frecuencia relativa (FR), la densidad absoluta (DA) y la densidad relativa (DR) de todas las especies muestreadas en los bancos de semillas. La frecuencia relativa de las especies se obtuvo de la frecuencia absoluta, dividiendo el número de parcelas en las que está presente la especie por el número total de parcelas. La densidad relativa se obtuvo a partir de la densidad absoluta calculada a partir del número total de especies presentes en una parcela dividida por el área total muestreada (Balestrin et al., 2019; Villa et al., 2019).
Las parcelas también fueron agrupadas con datos de abundancia de cada especie, a partir de una matriz de Bray-Curtis (disimilitud florística) usando el método de agrupamiento de medias aritméticas no ponderadas (UPGMA). De esta forma, se utilizó el índice de Sørensen para estimar el grado de similitud florística entre las parcelas. Todos los análisis se llevaron a cabo usando el programa R (R Core Team, 2018), excepto para el análisis UPGMA que se utilizó el programa PC-Ord 5.14 (ej. Villa et al., 2019).
Resultados
Flora del banco de semillas: Después de seis meses de monitoreo del banco de semillas, se registró un total de 1 362 plántulas germinadas considerando ambas profundidades. Del total de plantas registradas, 1 165 pertenecen a la capa superior (P1), que corresponde a aproximadamente el 85.5 % del total; mientras que la capa inferior (P2) se observó un total de 197 plántulas germinadas. Del total de plántulas germinadas, 47 especies correspondieron a 18 familias en P1; mientras que en la capa inferior del relave (P2), se registraron 12 familias y 23 especies.
Densidad y riqueza de especies: Considerando cada profundidad de relave muestreado, se observó un total de 485.4 individuos m² en P1, y de 82.09 individuos m² en P2. La familia que obtuvo la mayor densidad en número de individuos fue Cyperaceae, que equivale a 58.2 % y 60.4 % del total, para P1 y P2 respectivamente. Cuando se compara la variabilidad del número de individuos se observa que existen diferencias significativas (z =1,23, P < 0.001) entre profundidades (Fig. 2A). La riqueza de especies también difirió significativamente (P < 0.05) entre las profundidades. La riqueza de especies en el banco de semillas de la capa superior fue hasta más de 50 % mayor que en la capa inferior (Fig. 2B).
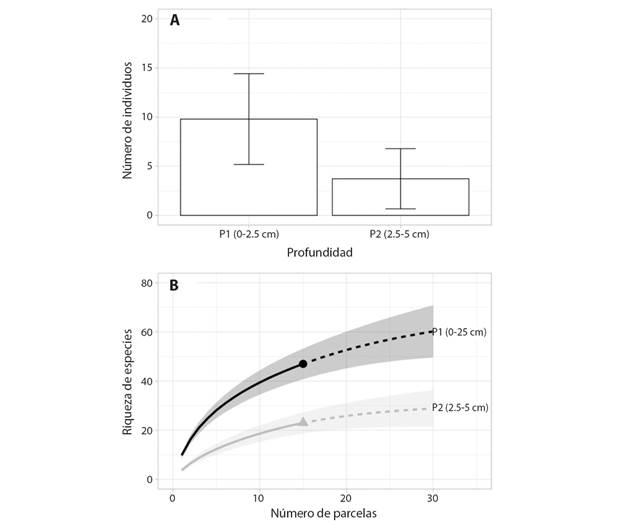
Fig. 2 A. Diferencias en la variación del número de individuos por parcela entre las dos profundidades (P1 y P2) de relaves. B. Curvas de rarefacción (línea continua) y extrapolación (líneas discontinuas) de riqueza de especies basada en el número de muestras de dos profundidades (P1 y P2) de relaves mineros en Mariana, Minas Gerais, Brasil. Las curvas de rarefacción y extrapolación presentan las líneas que representan los valores medios y las bandas la desviación estándar con intervalos de confianza del 95 %. /. Fig. 2. A. Differences in the variation in the number of individuals per plot between the two tailings depths (P1 and P2). B. Rarefaction curves (continuous line) and extrapolation (dashed lines) of species richness based on the number of samples from two depths (P1 and P2) of mining tailings in Mariana, Minas Gerais, Brazil. The rarefaction and extrapolation curves present the lines that represent the mean values and the standard deviation bands with 95 % confidence intervals.
Composición florística y parámetros fitosociológicos: Las familias con el mayor número de especies fueron Asteraceae (14 especies), Cyperaceae y Poaceae (cinco especies cada una) en ambas profundidades del relave. En la capa superior, se destacaron las familias Asteraceae, Solanaceae, Onagraceae y Lamiaceae, que representaron alrededor del 79 % del total (Tabla 1).
Tabla 1 Parámetros fitosociológicos en la capa superior del relave (P1 = 0-2.5 cm). / Table 1. Phytosociological parameters in the upper layer of the tailings (P1 = 0-2.5 cm)
| Nombre científico | NI | DA | DR | FA | FR |
| Cyperus rotundus L. | 365.0 | 2281.3 | 31.33 | 0.80 | 8.11 |
| Digitaria sanguinalis (L.) Scop. | 218.0 | 1362.5 | 18.71 | 0.33 | 3.38 |
| Cyperus difformis L. | 168.0 | 1050.0 | 14.42 | 0.27 | 2.70 |
| Hypolytrum pungens (Vahl) Kunth | 45.0 | 281.3 | 3.86 | 0.33 | 3.38 |
| Solanum paniculatum L. | 42.0 | 262.5 | 3.61 | 0.53 | 5.41 |
| Cyperus distans L. f. | 34.0 | 212.5 | 2.92 | 0.40 | 4.05 |
| Hyptis suaveolens (L.) Poit. | 27.0 | 168.8 | 2.32 | 0.20 | 2.03 |
| Ludwigia tomentosa (Cambess.) H. Hara | 27.0 | 168.8 | 2.32 | 0.20 | 2.03 |
| Solanum americanum Mill. | 26.0 | 162.5 | 2.23 | 0.67 | 6.76 |
| Pluchea sagittalis (Lam.) Cabrera | 24.0 | 150.0 | 2.06 | 0.53 | 5.41 |
| Erechtites hieraciifolius (L.) Raf. ex DC. | 21.0 | 131.3 | 1.80 | 0.60 | 6.08 |
| Melastomataceae sp. | 17.0 | 106.3 | 1.46 | 0.13 | 1.35 |
| Echinochloa colona (L.) Link | 16.0 | 100.0 | 1.37 | 0.13 | 1.35 |
| Lepidium virginicum L. | 16.0 | 100.0 | 1.37 | 0.47 | 4.73 |
| Vernonia polyanthes(Spreng.) Less. | 16.0 | 100.0 | 1.37 | 0.67 | 6.76 |
| Gnaphalium pensylvanicum Willd | 14.0 | 87.5 | 1.20 | 0.20 | 2.03 |
| Eleusine indica (L.) Gaertn. | 10.0 | 62.5 | 0.86 | 0.33 | 3.38 |
| Scoparia dulcis L. | 10.0 | 62.5 | 0.86 | 0.27 | 2.70 |
| Asteraceae sp. | 8.0 | 50.0 | 0.69 | 0.13 | 1.35 |
| Gnaphalium purpureum L. | 6.0 | 37.5 | 0.52 | 0.27 | 2.70 |
| Acanthospermum australe (Loefl.) Kuntze | 5.0 | 31.3 | 0.43 | 0.07 | 0.68 |
| Solanum mauritianum Scop | 5.0 | 31.3 | 0.43 | 0.20 | 2.03 |
| Begonia cucullata Willd. | 4.0 | 25.0 | 0.34 | 0.07 | 0.68 |
| Eclipta alba (L.) Hassk. | 4.0 | 25.0 | 0.34 | 0.07 | 0.68 |
| Ixora gardneriana Benth | 4.0 | 25.0 | 0.34 | 0.13 | 1.35 |
| Eleocharis elegans(Kunth) Roem. & Schult. | 3.0 | 18.8 | 0.26 | 0.07 | 0.68 |
| Mikania sp. | 3.0 | 18.8 | 0.26 | 0.20 | 2.03 |
| Brachiaria plantaginea (Link.) Hitchc | 2.0 | 12.5 | 0.17 | 0.07 | 0.68 |
| Clidemia hirta (L.) D. Don | 2.0 | 12.5 | 0.17 | 0.13 | 1.35 |
| Conyza canadensis (L.) Cronquist | 2.0 | 12.5 | 0.17 | 0.13 | 1.35 |
| Ludwigia alternifolia L. | 2.0 | 12.5 | 0.17 | 0.07 | 0.68 |
| Mikania cordifolia(L.f.) Willd | 2.0 | 12.5 | 0.17 | 0.13 | 1.35 |
| Não identificada 1 | 2.0 | 12.5 | 0.17 | 0.13 | 1.35 |
| Phyllanthus tenellus Roxb. | 2.0 | 12.5 | 0.17 | 0.07 | 0.68 |
| Ageratum conyzoides L. | 1.0 | 6.3 | 0.09 | 0.07 | 0.68 |
| Cecropia hololeuca Miq. Commelina diffusa Burm. f. | 1.0 1.0 | 6.3 6.3 | 0.09 0.09 | 0.07 0.07 | 0.68 0.68 |
| Erechtites valerianifolius (Link ex Spreng.) DC. | 1.0 | 6.3 | 0.09 | 0.07 | 0.68 |
| Glycine wightii (Graham ex Wight & Arn.) Verdc. | 1.0 | 6.3 | 0.09 | 0.07 | 0.68 |
| Não identificada 2 | 1.0 | 6.3 | 0.09 | 0.07 | 0.68 |
| Paspalum paniculatumL. | 1.0 | 6.3 | 0.09 | 0.07 | 0.68 |
| Rubus rosifolius Sm. | 1.0 | 6.3 | 0.09 | 0.07 | 0.68 |
| Sida rhombifolia L. | 1.0 | 6.3 | 0.09 | 0.07 | 0.68 |
| Spermacoce latifolia Aubl. | 1.0 | 6.3 | 0.09 | 0.07 | 0.68 |
| Trema micrantha (L.) Blume | 1.0 | 6.3 | 0.09 | 0.07 | 0.68 |
| Triumfetta semitriloba Jacq. | 1.0 | 6.3 | 0.09 | 0.07 | 0.68 |
| Vernonanthura phosphorica (Vell.) H. Rob. | 1.0 | 6.3 | 0.09 | 0.07 | 0.68 |
| Total | 1165 | 7282.4 | 100.0 | 9.91 | 100.0 |
Número de individuos de todas las especies (NI), frecuencia absoluta (FA), frecuencia relativa (FR), densidad absoluta (DA) y densidad relativa (DR). / Number of individuals of all species (NI), absolute frequency (FA), relative density (FR), absolute density (DA) and relative density (DR).
En la capa inferior, se destaca la presencia de las familias Solanaceae, Asteraceae y Lamiaceae, consecutivamente, que representan el 47.11 % del total de semillas emergentes (Tabla 2).
Tabla 2 Parámetros fitosociológicos en la capa inferior del relave (P2 = 2.5-5.0 cm). / Table 2. Phytosociological parameters in the lower layer of the tailings (P2 = 2.5-5.0 cm)
| Nombre científico | NI | DA | DR | FA | FR |
| Cyperus rotundus L. | 79.00 | 493.75 | 40.10 | 0.60 | 16.07 |
| Digitaria sanguinalis (L.) Scop. | 27.00 | 168.75 | 13.71 | 0.40 | 10.71 |
| Cyperus difformis L. | 23.00 | 143.75 | 11.68 | 0.33 | 8.93 |
| Solanum americanum Mill. | 12.00 | 75.00 | 6.09 | 0.47 | 12.50 |
| Cyperus distans L. f. | 11.00 | 68.75 | 5.58 | 0.13 | 3.57 |
| Hypolytrum pungens (Vahl) Kunth | 6.00 | 37.50 | 3.05 | 0.07 | 1.79 |
| Hyptis suaveolens (L.) Poit. | 5.00 | 31.25 | 2.54 | 0.07 | 1.79 |
| Pluchea sagittalis (Lam.) Cabrera | 5.00 | 31.25 | 2.54 | 0.07 | 1.79 |
| Scoparia dulcis L. | 5.00 | 31.25 | 2.54 | 0.27 | 7.14 |
| Echinochloa colona (L.) Link | 2.00 | 12.50 | 1.02 | 0.07 | 1.79 |
| Eclipta alba (L.) Hassk. | 2.00 | 12.50 | 1.02 | 0.13 | 3.57 |
| Eleusine indica (L.) Gaertn. | 2.00 | 12.50 | 1.02 | 0.13 | 3.57 |
| Erechtites hieraciifolius (L.) Raf. ex DC. | 2.00 | 12.50 | 1.02 | 0.13 | 3.57 |
| Ludwigia tomentosa(Cambess.) H. Hara | 2.00 | 12.50 | 1.02 | 0.07 | 1.79 |
| Melastomataceae sp. | 2.00 | 12.50 | 1.02 | 0.13 | 3.57 |
| Paspalum paniculatumL. | 2.00 | 12.50 | 1.02 | 0.13 | 3.57 |
| Rubus rosifolius Sm. | 2.00 | 12.50 | 1.02 | 0.07 | 1.79 |
| Sida rhombifolia L. | 2.00 | 12.50 | 1.02 | 0.07 | 1.79 |
| Solanum paniculatum L. | 2.00 | 12.50 | 1.02 | 0.13 | 3.57 |
| Glycine wightii (Graham ex Wight & Arn.) Verdc. | 1.00 | 6.25 | 0.51 | 0.07 | 1.79 |
| Gnaphalium pensylvanicum Willd | 1.00 | 6.25 | 0.51 | 0.07 | 1.79 |
| Lepidium virginicum L. | 1.00 | 6.25 | 0.51 | 0.07 | 1.79 |
| Vernonia polyanthes(Spreng.) Less. | 1.00 | 6.25 | 0.51 | 0.07 | 1.79 |
| Total | 197.0 | 1231.2 | 100.0 | 3.73 | 100.0 |
Número de individuos de todas las especies (NI), frecuencia absoluta (FA), frecuencia relativa (FR), densidad absoluta (DA) y densidad relativa (DR). / Number of individuals of all species (NI), absolute frequency (FA), relative density (FR), absolute density (DA) and relative density (DR).
Banco de semillas por categoría de dispersión y regeneración: Con respecto al origen de las especies, se observó que sólo el 2.7 % son de origen exótico y 97.3 % nativas (Tabla 3). En relación con el grupo ecológico de regeneración, ambas profundidades presentaron un mayor número de individuos pioneros y secundarios. En P1, se encontraron 71 % de los individuos pioneros y 14 % de los secundarios iniciales; mientras que en P2, 58 % pioneros y 23 % secundarios iniciales. Del número total de individuos, dos especies se identificaron como secundarias tardías en P1, Ixora gardneriana y Rubus rosifolius; mientras que en P2 solo se registró Rubus rosifolius. Por otro lado, se observó que, del total de especies germinadas, el 89.35 % correspondió a herbáceas, seguido por 7.05 % arbustivas, 0.9 % de especies arbóreas y 0.3 % de trepadoras o lianas. Finalmente, con la clasificación de síndromes de dispersión se observó que el 84.12 % de las especies fueron anemocóricas, 9.87 % zoocóricas y 3.09 % autocóricas (Tabla 3).
Tabla 3 Especies clasificadas de acuerdo a la estrategia de regeneración, síndrome de dispersión, origen y forma de vida. / Table 3. Species classified according to regeneration strategies, dispersal syndrome, origin and forms of life
| Familias/Especies | ER | DISP | ORG | FV |
| Asteraceae | ||||
| Acanthospermum australe (Loefl.) Kuntze | Nc | Zoo | N | H |
| Ageratum conyzoides L. | Pi | Zoo | N | H |
| Asteraceae sp. | - | - | - | - |
| Conyza canadensis (L.) Cronquist | Pi | Ane | E | H |
| Eclipta alba (L.) Hassk. | Si | Ane | N | H |
| Erechtites hieraciifolius (L.) Raf. ex DC. | Pi | Ane | N | H |
| Gnaphalium pensylvanicum Willd | Si | Ane | N | H |
| Gnaphalium purpureum L. | Si | Ane | N | H |
| Mikania sp. | - | - | - | - |
| Mikania cordifolia (L.f.) Willd | Pi | Ane | N | T |
| Pluchea sagittalis (Lam.) Cabrera | Nc | Ane | N | H |
| Vernonanthura phosphorica (Vell.) H. Rob. | Pi | Ane | N | B |
| Vernonia polyanthes (Spreng.) Less. | Pi | Ane | N | B |
| Begoniaceae | ||||
| Begonia cucullata Willd. | Pi | Ane | N | H |
| Brassicaceae | ||||
| Lepidium virginicum L. | Pi | Auto | E | H |
| Cannabaceae | ||||
| Trema micrantha (L.) Blume | Pi | Zoo | N | A |
| Commelinaceae | ||||
| Commelina diffusa Burm. f. | Pi | Auto | N | H |
| Cyperaceae | ||||
| Cyperus difformis L. | Pi | Ane | N | H |
| Cyperus distans L. f. | Pi | Ane | N | H |
| Cyperus rotundus L. | Pi | Ane | N | H |
| Eleocharis elegans(Kunth) Roem. & Schult. | Nc | Auto | N | H |
| Hypolytrum pungens (Vahl) Kunth | Nc | Ane | N | H |
| Fabaceae | ||||
| Glycine wightii (Graham ex Wight & Arn.) Verdc. | Nc | Nc | E | T |
| Lamiaceae | ||||
| Hyptis suaveolens (L.) Poit. | Pi | Zoo | N | H |
| Malvaceae | ||||
| Sida rhombifolia L. | Si | Auto | N | H |
| Triumfetta semitriloba Jacq. | Nc | Auto | N | A |
| Melastomataceae | ||||
| Clidemia hirta (L.) D. Don | Si | Zoo | N | B |
| Melastomataceae sp. | - | - | - | - |
| Onagraceae | ||||
| Ludwigia alternifolia L. | Nc | Nc | E | H |
| Ludwigia tomentosa (Cambess.) H. Hara | Pi | Ane | N | B |
| Phyllantaceae | ||||
| Phyllanthus tenellus Roxb. | Nc | Auto | N | H |
| Plantaginaceae | ||||
| Scoparia dulcis L. | Si | Auto | N | H |
| Poaceae | ||||
| Brachiaria plantaginea (Link.) Hitchc. | Pi | Ane | E | H |
| Digitaria sanguinalis (L.) Scop. | Pi | Ane | N | H |
| Echinochloa colona (L.) Link | Pi | Ane | N | H |
| Eleusine indica (L.) Gaertn. | Pi | Ane | E | H |
| Paspalum paniculatumL. | Nc | Auto | N | H |
| Brachiaria plantaginea (Link.) Hitchc. | Pi | Ane | E | H |
| Rosaceae | ||||
| Rubus rosifolius Sm. | St | Zoo | N | B |
| Rubiaceae | ||||
| Ixora gardneriana Benth. | St | Zoo | N | A |
| Spermacoce latifolia Aubl. | Si | Auto | N | H |
| Solanaceae | ||||
| Solanum americanum Mill. | Pi | Zoo | N | H |
| Solanum mauritianum Scop. | Pi | Zoo | N | A |
| Solanum paniculatum L. | Pi | Zoo | N | B |
| Urticaceae | ||||
| Cecropia hololeuca Miq. | Pi | Zoo | N | A |
Se indican las estrategias de regeneración de las especies, tales como pioneras (Pi), secundarias iniciales (Si), secundarias tardías (St); síndrome de dispersión tales como autocóricas (Auto), anemocóricas (Ane), zoocóricas (Zoo); origen como nativas (N) y exóticas (E) y por sus formas de vida, herbáceas (H), arbustos (B) y árboles (A). / Regeneration strategies of species, such as pioneer (Pi), early secondary (Si), late secondary (St) species; dispersal syndrome such as autochoric (Auto), anemochoric (Ane), zoochoric (Zoo); origin between native (N) and exotic (E) and by their forms of life, herbaceous (H), shrubs (B) and trees (A) are indicated.
Patrones de similitud florística: Los resultados demuestran una alta similitud florística entre las parcelas de las dos profundidades (Fig. 3). Por ejemplo, un grupo identificado con aproximadamente un 85 % de similitud florística, principalmente de especies gramíneas, se encuentra entre las parcelas P3, P18, P19 y P13. El análisis de dendrograma bidireccional indicó que la vegetación en el área de estudio podría dividirse en seis grupos con parcelas compartidas entre profundidades, mostrándose una alta similitud florística (rango entre 70 y 90 %) entre parcelas y grupos (Fig. 3).
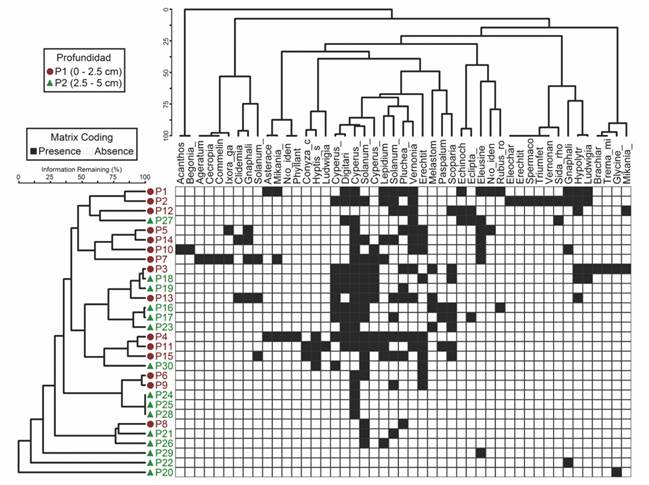
Fig. 3 Dendrograma de conglomerados bidireccional generado en base al índice de disimilitud de Bray-Curtis, que muestra la distribución de 49 especies en dos profundidades de relaves mineros y 30 parcelas en Mariana, Brasil. /. Fig. 3. Two-way cluster dendrogram generated based on Sorensen dissimilarity index, which shows the distribution of 49 species at two depths of mining tailings and 30 plots in Mariana, Brazil.
Composición de especies: La composición de especies no tuvo diferencias significativas entre las dos profundidades (Permanova: F1,28 = 2.72, P = 0.11), corroborando la alta similitud florística observada (Fig. 4). A pesar de la alta variabilidad de la composición florística en el estrato inferior (P2) explicada principalmente por el PCoA1 (52 %), existe una alta superposición de los dos grupos de profundidad en el espacio multivariante (Fig. 4).
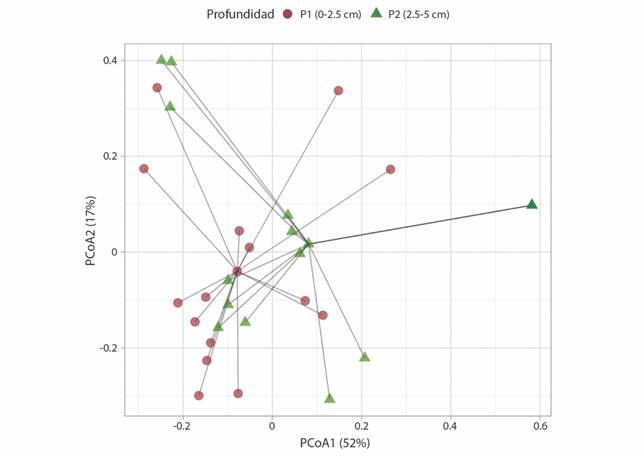
Fig. 4 Análisis de Coordenadas Principales (PCoA) basado en la métrica de disimilitud de Bray-Curtis entre en dos profundidades de relaves mineros en Mariana, Brasil. Se indican las capas superiores (P1 = 0-2.5 cm) e inferiores (P2 = 2.5-5.0 cm) de relaves. /. Fig. 4. Principal Coordinate Analysis (PCoA) based on the Bray-Curtis dissimilarity between the two depths of the mine mudslide in Mariana, Brazil. The upper (P1 = 0-2.5 cm) and lower (P2 = 2.5-5.0 cm) tailings layers are indicated.
Discusión
Los resultados de este estudio demostraron que actualmente prevalece un banco de semillas viables compuesto principalmente por especies nativas en las capas superficiales de relaves mineros acumulados en Mariana. La presencia de un banco de semilla en relaves mineros estudiados representa un importante indicador de recuperación de comunidades vegetales. La novedad de esta investigación fue observar la marcada diferencia existente entre las dos capas de relaves en términos de riqueza, y estructura (ej. abundancia y densidad de las semillas), a pesar de la poca diferencia de profundidad (0-5 cm) corroborando la hipótesis propuesta en esta investigación. Se presume que la composición de especies refleja una dispersión reciente el banco de semillas de la capa superior P1, además probablemente podría estar influenciado por la lluvia de semillas de los fragmentos forestales próximos. Por otro lado, el banco de semillas de la capa inferior P2, parece ser resultado de las capas de relaves mineros acumulados. Este patrón de banco de semillas puede tener importantes repercusiones sobre las tendencias futuras de recuperación de la vegetación en el área de estudio. Por este motivo, se discuten y establecen inferencias basadas en investigaciones y experiencias en diferentes escenarios de cambios en el uso de la tierra de paisajes tropicales, principalmente haciendo énfasis en los impactos de actividades mineras.
Diferentes estudios de bosques tropicales reportan que las semillas viables de especies pioneras almacenadas en el suelo pueden desempeñar un papel importante en la rápida regeneración después de los disturbios naturales o antropogénicos (ej. Dalling, Swaine, & Garwood, 1998; Martins & Engel, 2007; Mall & Singh, 2014; Sousa et al., 2017), generalmente depositadas en las primeras capas superficiales del suelo (Baider et al., 2001; Menezes et al., 2019). En este sentido, los resultados de este estudio demuestran un importante almacenamiento de semillas viables de especies pioneras en la primera capa del relave, lo que podría indicar su alta calidad y potencial para la regeneración natural. Por el contrario, bancos de semillas de baja calidad con baja densidad de especies pioneras puede retrasar la sucesión secundaria (Wijdeven & Kuzee, 2000). Este banco de semillas en las capas superficiales del relave en Mariana, representa un importante indicador ecológico clave para la restauración de la vegetación, presentando valores de densidad y riqueza de especies superiores a lo reportado previamente en áreas en recuperación después de actividades de minería (ej. Martins et al., 2008; Miranda-Neto & Martins, 2017; Miranda-Neto, Martins, & Silva, 2020). Además, se observó una alta proporción de especies nativas con dispersión anemocórica que representa un conjunto de características funcionales de relevancia para promover la regeneración natural inicial, como ha sido reportado en previos estudios en bosques de la Mata Atlántica (Correia & Martins, 2015; Balestrin et al., 2019; Miranda-Neto et al., 2020).
Nuestros resultados proporcionan datos de referencia sobre la calidad y potencial del banco de semillas en el área de estudio; es decir, alta riqueza de especies y densidad de semillas, como indicadores clave para seguir monitoreando y llevando a cabo diferentes estrategias de restauración en áreas impactadas por relaves de minería en Mariana. Por un lado, se puede conducir la regeneración natural de acuerdo a los niveles de calidad de los bancos de semillas, y por otro lado, a través de diferentes métodos de restauración activa en áreas de relaves con bancos de semillas de menor calidad y potencial de regeneración. Así mismo, los resultados de alta riqueza de especies y densidad de semillas reportadas en este estudio, permite presumir que es posible aplicar el traslado de semillas colectados en bosques secundarios próximos, los cuales probablemente también son responsables de la alta calidad de los bancos de semillas evaluados. Fue evidente que las áreas impactadas por los efectos de los relaves acumulados, generó la eliminación de la toda la cobertura vegetal existente; sin embargo, en este estudio se demuestra que el potencial de los bancos de semillas para la renovación y el rebrote pueden ser determinantes para la regeneración natural.
A pesar de las diferencias de riqueza, densidad y abundancia de especies entre profundidades, no se observaron diferencias en la composición florística. Sin embargo, las especies dominantes encontradas en los bancos de semillas en áreas afectadas por actividades mineras en paisajes de bosques tropicales han sido principalmente pioneras que promueven una rápida regeneración natural (Miranda-Neto et al., 2020; Martins, Fonseca, Cosimo, & Balestrin, 2021). Por ejemplo, los resultados indican que la proporción de especies pioneras entre profundidades varía entre 70 y 58 %, mientras que de especies secundarias iniciales entre 14 y 23 %. Estos grupos de regeneración de rápido crecimiento y exigentes de luz durante la regeneración (Villa et al., 2019), presentan semillas persistentes en el suelo, la mayoría viables para la germinación a largo plazo, hasta cuando se presenten las condiciones favorables para su germinación, principalmente después de un disturbio (Chazdon, 2014; Mall & Singh, 2014; Bechara et al., 2020; Miranda-Neto et al., 2020). Por el contrario, un menor número de especies secundarias tardías (tolerantes a la sombra) en el banco de semillas, representa un indicador del nivel de disturbio, ya que estas semillas presentan baja viabilidad y alto grado de depredación como principales limitaciones para permanecer en el suelo durante largos periodos de tiempo (Wijdeven & Kuzee, 2000; Chazdon, 2014). También es posible que exista una limitación de dispersión de semillas de especies secundarias tardías tolerantes a la sombra, y por su característica recalcitrante no ingresan al banco de semillas del suelo, por lo tanto, muchas especies clave con un banco de semillas persistente pueden ser escasas para promover eficazmente la recuperación natural de la vegetación (Török, Helm, Kiehl, Buisson, & Valkó, 2018; Kildisheva et al., 2020; Mudrák, Řehounková, Vítovcová, Tichý, & Prach, 2021). La presencia de especies pioneras de árboles como Cecropia hololeuca, Solanum mauritianum y Trema micrantha, han sido reconocidas como especies clave que comienzan el proceso de regeneración forestal en áreas degradadas de la región (Martins & Engel, 2007; Martins, 2009; Miranda-Neto et al., 2020). Por este motivo, se destaca la importancia de estas especies pioneras para acelerar la regeneración natural, así como para establecer condiciones microclimáticas adecuadas para la posterior colonización de especies secundarias tardías (Chazdon, 2014; Martins et al., 2015; Villa et al., 2019).
Los resultados de este estudio muestran que, del total de especies germinadas, se observó un total de 485.4 individuos m² en P1, y de 82.09 individuos m² en P2. Las especies pioneras germinan, crecen y se reproducen rápidamente, por lo que su abundancia en el banco de semillas aumenta muy rápidamente (ej. Silva et al. 2019; Martins et al., 2021). Además, es probable que está marcada diferencia entre las dos profundidades de los relaves refleja la importancia de la proximidad de los bosques para enriquecer el banco de semillas, dinámica de transporte y acumulación de relaves, y características funcionales de las semillas (ej. tamaño, peso, síndrome de dispersión), como ha sido repostado en previos estudios (Baider et al., 2001; Menezes et al., 2019; Martins et al., 2021). En este sentido, en bosques tropicales de Panamá se registraron principalmente germinantes de árboles pioneros que tenían entre 330 y 1 090 semillas m2 dependiendo de la estación seca y húmeda, respectivamente (Dalling et al., 1997). Por otro lado, en la región de la Mata Atlántica de Brasil, los bosques siempreverdes maduros comprendían entre 389 y 482 semillas viables m2 (Baider et al., 2001). Todos estos estudios de bosques húmedos siempreverdes muestran que existe poca o ninguna asociación entre la composición florística y el banco de semillas, o entre la lluvia anual de semillas y el banco de semillas (Dalling et al., 1998; Mall & Singh, 2014). Nuestro estudio no fue una excepción, ya que no encontramos especies de árboles de sucesión tardía representadas en el banco de semillas según lo definido por los estudios florísticos de bosques tropicales. Sin embargo, del total de especies germinadas 89.35 % correspondió a herbáceas, seguido por 7.05 % arbustivas, 0.9 % de especies arbóreas, y 0.3 % de trepadoras o lianas, todas muy importantes en las etapas iniciales de sucesión. Sólo el 2.7 % de las especies registradas en este estudio son de origen exótico, es decir, el banco de semillas de relaves tiene buena calidad para la regeneración natural con especies nativas. Probablemente esto indica que el banco de semillas se recupera muy rápidamente. Sin embargo, es difícil predecir la trayectoria futura del proceso de regeneración de la vegetación.
Con la clasificación de síndromes de dispersión de especies registradas en los bancos de semillas, el 84.12 % de las especies fueron anemocóricas, 9.87 % zoocóricas, y 3.09 % autocóricas, lo que tiende a coincidir con resultados de otros estudios en bosques de la Mata Atlántica (ej. Baider et al., 2001; Martins et al., 2008; Silva et al., 2019). Por ejemplo, en otros estudios también se ha observado un patrón similar de síndromes de dispersión con especies anemocóricas (entre 50 y 60 %) y zoocóricas (entre 15 y 30 %) en bosques tropicales (Chazdon, 2014). De esta forma, el patrón de composición florística observado en los bancos de semillas, principalmente la alta proporción de especies anemocóricas, permite presumir la alta similitud florística entre parcelas y profundidades. Además de la importancia del síndrome de dispersión, dicha agrupación florística puede estar relacionada con ambientes con disturbios a plena exposición solar y sin cobertura vegetal, lo que puede favorecer la llegada de semillas desde los bosques cercanos (Martins et al., 2021; Múdrak et al., 2021). Así mismo, es notable una tendencia hacia un mayor grupo de especies de dispersión herbácea, pionera y anemocórica, que son características notables de este banco de semillas, lo que puede estar induciendo una rápida sucesión secundaria.
En este estudio se determinó que el banco de semillas viables en las capas superficiales de relaves mineros acumulados en el área de estudio, conformados principalmente por especies de origen nativo, pioneras, y anemocóricas, presentan buena calidad y potencial para la regeneración natural. Estos resultados demuestran que este patrón de diversidad y características funcionales de las especies (síndromes de dispersión y estrategia de regeneración) pueden ser determinante para promover la regeneración natural de la vegetación en las áreas afectadas con acumulación de relaves mineros en Mariana. Además, este patrón de diversidad y estructura del banco de semillas puede tener importantes repercusiones sobre estrategias de restauración y monitoreo de áreas afectadas por relaves, ya que ambas profundidades de los relaves reflejan la importancia de la proximidad de los bosques para enriquecer el banco de semillas. El enfoque utilizado en nuestro estudio demuestra una base para la distribución de la abundancia y diversidad de especies en la gestión y ecología forestal aplicada, que puede utilizarse para la identificación de especies clave y grupos funcionales que pueden promover la recuperación de los bosques mediante la regeneración natural y restauración activa.
Declaración: los autores declaran que todos están de acuerdo con este artículo y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes para garantizar su publicación.

