











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
La degradación y conversión de hábitats naturales se han constituido en una seria amenaza para la biodiversidad (Vitousek, Mooney, Lubchenco, & Melillo, 1997; Hooke, Martín-Duque, & Pedraza, 2012). En la región tropical las grandes masas forestales han sido transformadas a paisajes dominados por pastizales y tierras agrícolas, con fragmentos de vegetación natural aislados y en diferente estado de degradación (Laurance, Sayer, & Cassman, 2014). Estos cambios en los hábitats naturales están causando una reducción en las poblaciones de vertebrados y, en muchos casos, extinciones a escala local y global, generando un proceso de defaunación de estos ecosistemas (Dirzo et al., 2014; Gardner, Bicknell, Baldwin-Cantello, Struebig, & Davies, 2019). Estos procesos de defaunación, a su vez está generando cambios en las interacciones planta-animal y limitando los servicios ecosistémicos que estos grupos brindan al ecosistema (Dirzo et al., 2014).
Uno de los servicios ecosistémicos que ha mostrado estar fuertemente impactado por la defaunación es la dispersión de semillas, lo cual es particularmente preocupante en los ecosistemas tropicales donde alrededor del 80 % de las especies de plantas dependen de animales para su dispersión (Jordano, 2000). Estudios previos señalan que los cambios en la comunidad de frugívoros generan una reducción en la tasa de remoción de semillas (Bello et al., 2015; Boissier, Henry, & Forget, 2020), con implicaciones para la biomasa vegetal y, por ende, para las tasas de almacenamiento de carbono (Bello et al., 2015; Peres, Emilio, Schietti, Desmoulière, & Levi, 2016). Un grupo importante para el mantenimiento de los procesos de dispersión de semillas lo constituyen los grandes mamíferos. Su alta capacidad de movimiento hace de este grupo altamente eficiente para la dispersión de semillas a largas distancias (Guimaraes, Galetti, & Jordano, 2008; Campos-Arceiz & Blake, 2011; Pires, Guimaraes, Galetti, & Jordano, 2018). Además, las funciones que cumplen no siempre pueden ser desempeñadas por especies de menor tamaño (Galetti & Dirzo, 2013; O’Farrill, Galetti, & Campos-Arceiz, 2013; Peres et al., 2016). El rol de los grandes mamíferos es especialmente importante para dispersar los denominados frutos anacrónicos, es decir, frutos dispersados por megafauna del Pleistoceno y que persisten en la flora contemporánea (Janzen & Martin, 1982; Guimaraes et al., 2008). La evidencia señala que los frutos anacrónicos de los bosques tropicales estacionalmente secos (BTES), estuvieron potencialmente dispersados durante el Pleistoceno por megafauna extinta que incluye especies de las familias Gomphotheriidae, Equidae, Tayassuidae y Cervidae (Janzen & Martin, 1982; Lindsey & Lopez, 2015). Actualmente, algunos de estos frutos están siendo dispersados por el venado de cola blanca, Odocoileus virginianus Trouessart (Jara-Guerrero, Escribano-Avila, Espinosa, De la Cruz, & Méndez, 2018), los cuales se caracterizan por ser de tamaño grande, colores opacos, pulpa seca y fibrosa y, en muchos casos, sin adaptaciones obvias para la dispersión por animales (Janzen & Martin, 1982; Jara-Guerrero et al., 2018). Varias de estas características hacen que únicamente animales de gran tamaño puedan ofrecer los servicios de dispersión. Lamentablemente, los grandes mamíferos son uno de los primeros grupos de animales silvestres en ser afectados por las presiones antrópicas (Dirzo et al., 2014).
En aquellos paisajes alterados predominantes en los trópicos la función de dispersión de semillas de grandes mamíferos ha sido parcialmente asumida por animales domésticos, entre estos, herbívoros como cabras y bovinos (Bartuszevige & Endress, 2008; Esquivel, Harvey, Finegan, Casanoves, & Skarpe, 2008). Estos rumiantes domésticos podrían jugar un papel importante en la dinámica de la vegetación (Esquivel et al., 2008; Guimaraes et al., 2008; Chazdon et al., 2009; Pignataro et al., 2017). Sin embargo, hasta qué punto pueden mantener los procesos naturales es algo que dependerá de los procesos de masticación y digestión y de sus preferencias de alimentación, pero también de la naturaleza del ecosistema y las características adaptativas de las especies que lo componen (Schupp, Jordano, & Gómez, 2010; Albert et al., 2015). El rol de los rumiantes como dispersores efectivos ha sido ampliamente discutido debido a que su proceso digestivo podría dañar las semillas durante la ingestión o la rumia, ya sea por desgaste o ruptura de la testa (Gardener, McIvor, & Jansen, 1993). De esta forma, su papel como dispersores depende en gran medida de las características de las semillas, como la dureza e impermeabilidad de la cubierta, características que las protegen de daños por los molares o por los ácidos gástricos (Janzen & Martin, 1982; Albert et al., 2015). A pesar de estos potenciales efectos negativos, algunos estudios señalan un alto potencial de rumiantes domésticos como dispersores de semillas y facilitadores de la germinación en diversos ecosistemas, como praderas templadas (Cosyns, Bossuyt, Hoffmann, Vervaet, & Lens, 2006; Bartuszevige & Endress, 2008; Kuiters & Huiskes, 2010), matorrales mediterráneos (Malo, Jiménez, & Suarez, 2000; Manzano, Malo, & Peco, 2005; Mancilla-Leytón, Fernandez-Alés, & Vicente, 2015; Delibes, Castañeda, & Fedriani, 2019) y en bosques secos del Neotrópico (Baraza & Valiente-Banuet, 2008). Para determinar el rol de las especies domésticas en la dispersión es importante evaluar, además de los efectos del paso por el tracto digestivo, su comportamiento alimenticio; estudios previos señalan también posibles preferencias por especies de alto contenido nutricional (Albert et al., 2015), como las leguminosas (Miceli-Méndez, Ferguson, & Ramírez-Marcial, 2008). Por otro lado, la movilidad entre áreas antropizadas y bosques nativos, puede tener efectos en la disposición final y la estructura de la comunidad. Por ejemplo, en pasturas de Texas, Estados Unidos se ha reportado que los rumiantes domésticos modifican la estructura de la vegetación al dispersar una gran cantidad semillas de Prosopis glandulosa Torr. (Fabaceae) desde el bosque a los pastizales (Kneuper, Scott, & Pinchak, 2003). En los BTES el enriquecimiento de parcelas con fecas de cabras mostró un cambio en la estructura de la comunidad con el incremento de la dominancia de especies asociadas a asentamientos humanos, como Acacia macracantha Humb. & Bonpl. ex Willd. (Rodriguez, 2006).
En los BTES del Neotrópico es urgente esclarecer el papel del ganado como dispersor de semillas debido a que la mayor parte de este ecosistema está bajo procesos de degradación crónica (Miles et al., 2006), y la ganadería junto con la extracción de leña están entre las principales presiones (Ribeiro, Arroyo-Rodríguez, Santos, Tabarelli, & Leal, 2015; Jara-Guerrero, Maldonado, Espinosa, & Duncan, 2019). No obstante, la forma tradicional de crianza del ganado caprino y bovino en estos bosques, que implica el forrajeo libre dentro del bosque a lo largo del año (Jara-Guerrero et al., 2019), podría implicar una contribución potencial para la dispersión de especies leñosas a lo largo del paisaje y para la restauración de áreas degradadas (Chazdon et al., 2009).
En el presente documento evaluamos en qué medida las cabras pueden estar aportan la dispersión de semillas de especies leñosas y las posibles consecuencias de su comportamiento alimenticio sobre la estructura de la vegetación. Específicamente planteamos tres objetivos; (1) Evaluar la importancia de las cabras como dispersores de semillas de especies forestales de los BTES; (2) evaluar el efecto del paso por el tracto digestivo sobre la germinación de las especies dispersadas; (3) analizar en qué medida la vegetación establecida define los patrones de dispersión de semillas por cabras. Nuestra hipótesis fue que el rol de las cabras como dispersoras de semillas es similar al de los ungulados silvestres, tanto en el tipo de frutos dispersados como en el efecto sobre la germinación. Por otro lado, planteamos que la composición de especies dispersadas es independiente de la oferta de recursos en el bosque, y puede ser alterada por especies con altas abundancias en zonas pobladas. Este patrón en la dispersión de semillas podría estar afectando la distribución y abundancia de semillas en el suelo del bosque y, por tanto, modificar la estructura de la vegetación en las áreas de pastoreo. De cumplirse esta hipótesis, la composición de especies dispersadas por será más homogénea a lo largo del área de estudio que la composición de especies de la vegetación establecida. Además, no se debería encontrar ninguna relación en la abundancia y riqueza entre la vegetación establecida y la abundancia y riqueza de semillas dispersadas.
Materiales y métodos
Área de estudio: El presente estudio se llevó a cabo en el BTES del cantón Zapotillo, provincia de Loja al Suroccidente de Ecuador (Fig. 1). Estos bosques son parte del hotspot Tumbes-Choco-Magdalena (Mittermeier et al., 2004) y de la región biogeográfica Pacífico Ecuatorial (Espinosa et al., 2012) y comprenden algunos de los parches de bosque más grandes y mejor conservados de esta región (Best & Kessler, 1995). El clima es tropical, con una temperatura media anual de 22.2 °C y precipitación media anual de 642 mm (Espinosa, Camarero, & Gusmán, 2018). Con una temporada seca que se extiende de mayo a diciembre, durante la cual la precipitación mensual eventualmente excede los 10 mm, y una estación lluviosa de cuatro meses, de enero a abril (Espinosa et al., 2018). La presión antropogénica en esta región aumentó desde mediados del siglo XX, principalmente debido a la expansión de áreas agrícolas, la crianza intensiva de cabras y la extracción de madera (Cueva-Ortiz et al., 2019; Jara-Guerrero et al., 2019). En la vegetación leñosa del área destacan las familias Fabaceae y Malvaceae, con 24 y seis especies, respectivamente. Las especies más abundantes en la zona de estudio son: Handroanthus chrysanthus (Jacq.) S.O.Grose, y Piscidia carthagenensis Jacq. (Cueva-Ortiz et al., 2019). Otras especies características de este ecosistema son: Ceiba trichistandra (A.Gray) Bakh., Cavanillesia platanifolia (Bonpl.) Kunth, Eriotheca ruizi (K. Schum.) A. Robyns, Terminalia valverdae A.H. Gentry, Bursera graveolens (Kunth) Triana & Planch (Aguirre, Kvist, & Sánchez, 2006).
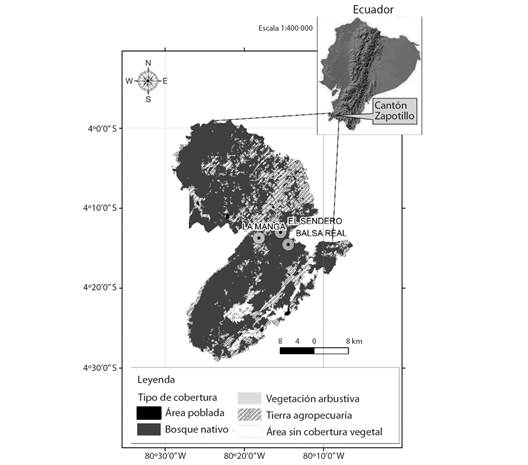
Fig. 1 Mapa del cantón Zapotillo. Se muestran las tres localidades de estudio: La Manga, Balsa Real y El Sendero. / Fig. 1. Zapotillo county map. Three study locations are shown: La Manga, Balsa Real and El Sendero.
Para el presente estudio se seleccionaron tres localidades de bosque seco separadas por al menos 5 km; Sendero, La Manga y Balsa Real (Fig. 1). Estas localidades fueron seleccionadas considerando la facilidad de acceso durante la mayor parte del año y el permiso de los propietarios para el acceso a los corrales. En cada localidad se seleccionaron dos corrales de cabras y un bloque de parcelas permanentes. Cada bloque está conformado por tres parcelas de 60 x 60 m en forma de “L”, con una separación mínima entre ellas de 200 m, las cuales fueron establecidas previamente por Cueva-Ortiz et al., (2019). En cada parcela se identificaron todos los individuos mayores o igual 5 cm de DAP.
Durante el estudio se determinó la presencia de cabras en las zonas cercanas a las parcelas permanentes, las cuales estarían forrajeando diariamente dentro de cada localidad. Se estimó un total de 47, 56, 45 cabras para El Sendero, La Manga y Balsa Real, respectivamente. La cantidad total de cabras se determinó mediante un censo con los propietarios. En cada una de las localidades el régimen de pastoreo es el mismo, el ganado pastorea libremente en las áreas de bosque cercano, dependiendo exclusivamente de los recursos que provee la vegetación. Todos los días el ganado es liberado alrededor de las 06:00 h, y regresa a los corrales al final de la tarde, entre las 17:00 a 18:00 h.
Entre la fauna silvestre que dispersa semillas se ha reportado en los BTES de esta región están algunos mamíferos como Lycalopex sechurae, Pecari tajacu, Odocolileus virginianus, Mazama americana, Cuniculus paca, Didelphis marsupialis y al menos cinco especies de murciélagos (Tirira, 2017). Además, alrededor de 27 especies de aves incluyen frutos como parte de su dieta (Ordóñez-Delgado et al., 2016).
Muestreo de fecas: Entre los meses de diciembre 2016 a junio 2017 se recolectaron mensualmente fecas de cabras en las zonas de bosque y en los corrales de cada localidad. Cada muestra estuvo compuesta por 2 kg de fecas frescas, con alto contenido de humedad. Para la recolección de fecas en bosque se estableció una franja de 5 m de ancho alrededor de los límites de cada parcela de vegetación. En las zonas de bosque no siempre se alcanzó la cantidad de muestra establecida, debido a que en la estación lluviosa las fecas se disgregaban rápidamente y, además, durante esta estación las cabras tienen patrones de movimiento más restringido (Obs. pers.), por lo que el tamaño de la muestra dependió de la cantidad de fecas disponibles. En los corrales las muestras fueron recolectadas aleatoriamente. Cada muestra estuvo compuesta de al menos 5 submuestras de fecas frescas tomadas dentro del corral. Las submuestras fueron mezcladas y pesadas hasta conseguir los 2 kg. Las fecas recolectadas tanto en bosque como en corrales se secaron a temperatura ambiente. Posteriormente fueron disgregadas y tamizadas con el fin de clasificar e identificar taxonómicamente las semillas. Para el registro de semillas dispersadas se consideró únicamente las semillas enteras. En conjunto, las muestras de fecas recolectadas tanto en bosque como en corrales se utilizaron para definir las especies que están siendo dispersadas. Las muestras de los corrales se utilizaron para evaluar los efectos del paso por el tracto digestivo sobre la germinación, mientras que las muestras recolectadas en el bosque se usaron para evaluar la relación entre la vegetación establecida y el patrón de dispersión de semillas.
Evaluación de germinación: Todas las semillas provenientes de las fecas de corrales se sembraron en sustrato de tierra negra y arena en una proporción 3:1 con el fin de evaluar el porcentaje de germinación. Las semillas provenientes del bosque no se usaron para evaluar la germinación, debido a que estaban sometidas a condiciones ambientales variables, pudiendo afectar los resultados. La siembra se la realizó en camas separadas por especie, fecha de colección y proveniencia de las muestras. Las camas se mantuvieron en un invernadero de la Universidad Nacional de Loja (4°01’55.49” S & 79°12’02.41” W) con condiciones de temperatura de entre 17 a 25 °C, se controló diariamente el riego manteniendo el sustrato húmedo. Cada tres días se realizó el control de germinación, registrando el día de germinación de cada semilla. Una vez que las semillas germinaron y alcanzaron un tamaño de alrededor de 10 cm, permitiendo confirmar la identidad, la plántula fue extraída de la cama de germinación para evitar reconteo. El control de la germinación se mantuvo por dos meses luego de la última siembra.
Para determinar la influencia del paso por el tracto digestivo sobre la germinación de semillas se realizó una siembra de semillas recolectadas directamente de los árboles como tratamiento control. Todas las especies en fructificación durante las visitas de campo y que podrían ser dispersadas por las cabras fueron recolectadas. Las semillas fueron recolectadas al azar en al menos 10 individuos cercanos a las zonas de estudio. Las semillas recolectadas fueron extraídas del fruto y una vez limpias se mezclaron para tomar una muestra aleatoria de 300 semillas por especie, a excepción de Ziziphus thyrsiflora Benth. que debido a la baja disponibilidad solo se sembraron 64 semillas. En el caso de Celtis iguanea (Jacq.) Sarg. no se encontraron individuos en fructificación luego de ser detectadas en las fecas, ni durante las visitas de campo. En el caso de Acacia macracantha Humb. & Bonpl. ex Willd, debido a un accidente durante el proceso de germinación, se perdieron las semillas sembradas y no fue posible obtener una nueva muestra de control. Las semillas fueron sembradas en seis lotes de 50 semillas, usando el mismo sustrato y condiciones que las semillas provenientes de las fecas. En el caso de Z. thyrsiflora se sembraron tres lotes de 20 semillas. Las semillas se sembraron al final del experimento y se realizó el monitoreo por 60 días.
Análisis de datos: Para evaluar los efectos del paso por el tracto digestivo sobre la germinación de semillas ajustamos curvas de supervivencia para cada especie y para cada tratamiento (semillas extraídas de las fecas provenientes de corrales y control). Las curvas se ajustaron usando la función “surv” del paquete “survival” del entorno de programación R. Posteriormente, ajustamos un test de diferencias en la curva de supervivencia usando la función “survdiff” del mismo paquete, que prueba las diferencias entre dos curvas usando un test de la familia G-rho (Therneau, 2020). Con el fin de que los datos sean comparables usamos los datos de germinación durante los primeros 60 días para todas las especies y para los dos tratamientos.
Para evaluar en qué medida las especies en la vegetación establecida afectan la dispersión de semillas evaluamos la relación de la riqueza, densidad y composición de especies entre la vegetación establecida y las semillas de las fecas obtenidas en el bosque. Utilizamos correlaciones de Pearson entre la riqueza de especies en la vegetación establecida y la riqueza de especies de semillas en las fecas del bosque. Desarrollamos un test Chi cuadrado para evaluar la relación entre la densidad de semillas encontradas en las fecas y la densidad de árboles establecidos en las parcelas de bosque. Obtuvimos la densidad de semillas dividiendo el número de semillas de cada especie para el área de muestreo de las fecas (5m x 240 m) y multiplicado por diez mil con el fin de tener un valor por hectárea. La densidad de la vegetación se obtuvo dividiendo la abundancia de cada especie para el área muestreada (60 m x 60 m) y multiplicando por diez mil. Adicionalmente, evaluamos los patrones de disimilitud entre parcelas usando los datos de presencia-ausencia tanto para la vegetación establecida como para las semillas provenientes de las fecas del bosque. Obtuvimos la disimilitud entre sitios usando la distancia de Bray Curtis usando a función “vegdist” del paquete “vegan” (Oksanen et al., 2019). Con base a las distancias obtenidas entre parcelas, separamos las distancias intra-sitio, distancias entre parcelas que se encuentran dentro de un mismo sitio, y las distancias inter-sitios, distancias entre parcelas de diferentes localidades. Con estos datos ajustamos un modelo lineal con el fin de evaluar si la disimilitud está determinada por la procedencia de la parcela (intra-sitio o inter-sitios). Todos los análisis fueron implementados en el entorno R (R Core Team, 2019).
Resultados
Dispersión endozoocora: En total se analizaron 80 muestras de fecas recolectadas entre los meses de diciembre de 2016 y junio de 2017, de las cuales 42 pertenecen a fecas obtenidas en las zonas de bosque y 38 corresponden a muestras de fecas obtenidas en corrales. Durante los muestreos de los meses de enero, febrero y junio, no se encontraron fecas en las zonas de bosque; es posible que durante este periodo las fecas se disgregaran más rápidamente debido a las lluvias. En total las cabras dispersaron semillas de 10 especies, de las cuales al menos siete son especies leñosas y seis corresponden a especies presentes en la vegetación establecida (Tabla 1). La especie Prosopis juliflora (Sw.) DC. no se registró en las parcelas de vegetación establecida, aunque esta especie se encuentra en las zonas de bosque. Otras tres especies que no se pudieron identificar se mantuvieron como morfotipos. La familia más representativa entre las especies registradas en las fecas fue Fabaceae (5 spp.), seguida por Cannabaceae y Rhamnaceae (1 spp.) (Tabla 1). Encontramos un total de 13 326 semillas, de las cuales 5 471 corresponden a las muestras de fecas encontradas en el bosque y 7 855 semillas corresponden a las fecas de los corrales. La especie dominante entre las semillas encontradas en las fecas fue Acacia macracantha. Esta especie fue la más frecuente (53/80 muestras) y abundante en las muestras recolectadas, con una abundancia relativa promedio de 62.05 % en las fecas del bosque y 83.43 % en las fecas de los corrales. Las otras nueve especies presentaron una frecuencia baja (< 50 %) y abundancias relativas por debajo del 13 % en bosque y 7 % en corrales. Albizia multiflora (Kunth) Barneby & J.W. Grimes, Celtis iguanaea y Z. trysiflora estuvieron presentes en alrededor del 50 % de las muestras de fecas, mientras que P. carthagenensis, Chloroleucon. mangense (Jacq.) Britton & Rose y P. juliflora fueron las especies menos frecuentes, con 18.4, 5.3 y 2.6 % respectivamente. En general, las especies dispersadas por cabras mostraron una baja abundancia relativa en la vegetación establecida, siendo P. carthagenensis la especie con mayor abundancia relativa (9 %), mientras que A. macracantha representó en promedio apenas un 2.33 % de la comunidad de leñosas establecidas (Tabla 1).
Tabla 1 Especies dispersadas en las fecas de cabra. / Table 1. Species dispersed in goat feces.
| FAMILIA / Especie | Vegetación | Semillas dispersadas | ||
| Frecuencia | Bosque | Corrales | ||
| FABACEAE | ||||
| Acacia macracantha | 2.33 (1.34) | 53 | 62.05 (8.54) | 83.43 (4.24) |
| Albizia multiflora | 1.67 (0.29) | 34 | 4.01 (0.93) | 5.03 (2.29) |
| Chloroleucon mangense | 4.33 (3.02) | 28 | 12.98 (2.87) | 0.05 (0.03) |
| Piscidia carthagenensis | 9.00 (1.37) | 8 | 0.01 (0.01) | 1.14 (0.52) |
| Prosopis juliflora | NA | 12 | 2.90 (0.94) | 0.16 (0.16) |
| CANNABACEAE | ||||
| Celtis iguanaea | 0.33 (0.33) | 35 | 2.4 (0.40) | 4.06 (1.98) |
| RHAMNACEAE | ||||
| Zizyphus thyrsiflora | 0.11 (0.11) | 29 | 0.22 (0.12) | 6.13 (2.69) |
| DESCONOCIDA | ||||
| sp1 | NA | 10 | 9.81 (6.63) | NA |
| sp2 | NA | 3 | 0.07 (0.04) | NA |
| sp3 | NA | 2 | 5.55 (5.50) | NA |
Vegetación: promedio (error estándar) de la abundancia relativa de individuos en la vegetación establecida por localidad. Frecuencia: frecuencia total de ocurrencia de semillas en las 80 muestras de fecas de cabra recolectadas en bosque (N = 42) y corrales (N = 38). Bosque: abundancia relativa promedio de semillas (error estándar) por localidad para fecas en bosque. Corrales: abundancia relativa promedio de semillas (error estándar) por localidad para fecas en corrales. NA: ausencia de la especie. La abundancia relativa en la vegetación establecida fue calculada del total de individuos de todas las especies encontradas en cada parcela. En la tabla se muestran únicamente los datos de las especies encontradas en fecas. / Vegetation: mean (standard error) of the relative abundance of individuals in the standing vegetation by locality. Frequency: total frequency of seeds occurrence in the 80 samples of goat feces collected from the forest (N = 42) and pens (N = 38). Forest: mean relative abundance of seeds by locality (standard error) for goat feces from the forest. Pens: mean relative abundance of seeds by locality (standard error) for goat feces from the pens. NA: species absence. The relative abundance in the standing vegetation was calculated from the total of individuals of all the species recorded in each plot. The table only shows the data for the species found in the goat feces.
Analizamos la variación temporal en la abundancia y riqueza de semillas recolectadas en los corrales, encontrando una mayor riqueza (5 spp.) y abundancia de semillas entre diciembre de 2016 y marzo de 2017. La menor riqueza y abundancia de semillas se registró entre los meses de abril a junio de 2017, con una contribución muy baja de semillas de especies diferentes a A. macracantha (Fig. 2). Por tres meses de muestreo no fue posible recuperar semillas del bosque debido a que las condiciones climáticas limitaban la acumulación de fecas, por tanto, no se presenta la variación temporal de las fecas provenientes del bosque.
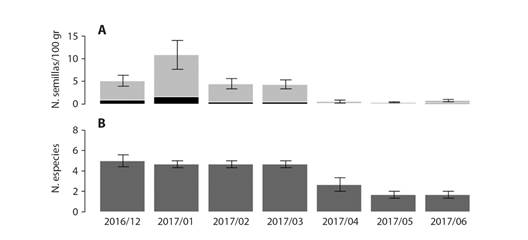
Fig. 2 Variación temporal de la riqueza y abundancia de semillas (promedio + error estándar) encontradas en las fecas de los corrales. A. Variación de la abundancia (N. semillas/100 gr.), el color gris de las barras muestra la abundancia de A. macracantha y el negro la abundancia de otras especies. B. Riqueza de especies en cada mes. / Fig. 2. Temporal variation in richness and abundance of seeds recorded in goat feces from pens (mean + SE). A. Abundance variation (N. Seeds /100 g.), grey bars show abundance of A. macracantha, and black bars show abundance of the other species. B. Species richness per month.
Efectos de la ingestión de semillas sobre la germinación: A partir de las semillas de 38 muestras de fecas de corral se registró germinación en cuatro de las siete especies leñosas sembradas. Las especies con mayores porcentajes de germinación fueron P. carthagenensis y Z. thyrsiflora, las cuales tuvieron una germinación de alrededor de 90 %, a partir de un total de semillas sembradas de 211 y 65, respectivamente. Por el contrario A. macracantha y A. multiflora presentaron porcentajes de germinación menores a 40 % partir de un total de semillas sembradas de 6 812 y 213, respectivamente (Tabla 2). Tres de las siete especies evaluadas (C. Iguanaea, C. mangense y P. juliflora) no presentaron germinación. En el caso de C. iguanea se sembraron más de 500 semillas, mientras que para las otras dos especies se sembró una cantidad menor a 20 semillas por especie (Tabla 2).
Tabla 2 Resultados de germinación a partir de semillas dispersadas por cabras y tratamiento control por especie. / Table 2. Results of germination from seeds dispersed and control treatment.
| Especie | Dispersadas | Control | ||
| Semillas | Germinación | Semillas | Germinación | |
| Acacia macracantha | 6812/1087 | 16 | NA | NA |
| Albizia multiflora | 213/83 | 39 | 300/19 | 6 |
| Celtis iguanaea | 531/0 | 0 | NA | NA |
| Chloroleucon mangense | 6/0 | 0 | 300/206 | 69 |
| Piscidia carthagenensis | 65/59 | 91 | 300/68 | 23 |
| Prosopis juliflora | 17/0 | 0 | 300/130 | 43 |
| Zizyphus thyrsiflora | 211/186 | 88 | 64/53 | 83 |
Semillas: número total de semillas sembradas / número total de semillas germinadas; Germinación: porcentaje total de germinación. NA: dato no disponible. / Seeds: total number of sown seeds / total number of germinated seeds, Germination: total percentage of germination.
Para cinco de las siete especies registradas en las fecas de corral se logró recolectar semillas directamente de la planta. Con estas especies se evaluó el efecto del paso por el tracto digestivo sobre la germinación, encontrando una variación entre especies. De acuerdo con el análisis de supervivencia en tres de las cinco especies evaluadas; A. multiflora, P. carthagenensis y Z. thyrsiflora el paso por el tracto digestivo mejoró significativamente el porcentaje, así como la velocidad de germinación (Fig. 3). En las otras dos especies analizadas; C. mangense y P. juliflora, no se obtuvo germinación de las semillas provenientes de fecas, mientras que las semillas testigo alcanzaron porcentajes de 60 y 50 %, respectivamente.
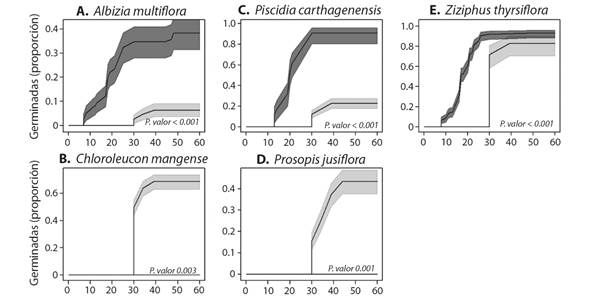
Fig. 3 Curvas de germinación basadas en el análisis de supervivencia. Las líneas con la sombra gris obscura corresponden a los datos de semillas encontradas en las fecas de cabras. Líneas con la sombra gris clara corresponden a las semillas de tratamiento testigo, recolectadas de la planta. La sombra en cada curva corresponde al intervalo de confianza. En cada figura se muestra el P calculado basado en la familia G-rho. / Fig. 3. Germination curves based on survival analysis. Lines with the dark gray shade correspond to the data of seeds from goat feces. Lines with the light gray shadow correspond to the control treatment. The shadow of each curve corresponds to the confidence interval. Each figure shows the calculated P based on the G-rho family.
Patrón de dispersión de semillas: No encontramos una correlación entre la riqueza de árboles establecidos y el número de especies dispersadas en las zonas de bosque (χ2 = -0.23, P = 0.53). Tal como esperábamos, la disimilitud entre sitios fue mayor en la vegetación establecida que en las semillas de las fecas. Los análisis basados en datos de presencia - ausencia señalan la vegetación establecida tiene una disimilitud significativamente mayor entre sitios que entre parcelas del mismo sitio, mientras que no encontramos diferencias significativas en la disimilitud de las especies registradas en las fecas provenientes del bosque (Fig. 4).
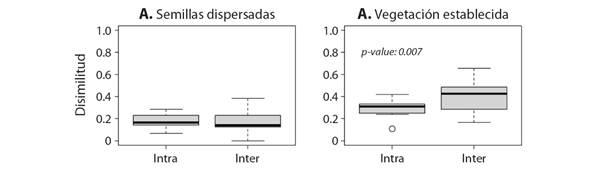
Fig. 4 Disimilitud intra-sitio (Intra) e inter-sitios (Inter) en las A. semillas dispersadas por las cabras y en B. la vegetación establecida. / Fig. 4. Dissimilarity intra-site (Intra) and inter-site (Inter) in A. seeds dispersed by goats and B. standing vegetation.
El test Chi cuadrado mostró un patrón de dispersión de semillas no asociado a la abundancia de individuos en la vegetación establecida (χ2 = 104.93, D.F. = 6, P < 0.001). La abundancia relativa de árboles en la vegetación establecida fue menor que la abundancia relativa de semillas de A. macracanta, C. iguanea y P. juliflora en las fecas, y mayor que la abundancia relativa de semillas de P. cartagenensis, C. mangense y A. multiflora (Tabla 1).
Discusión
La crianza de cabras basada en el forrajeo libre dentro de los remanentes de BTES del Neotrópico es una actividad practicada durante décadas, por lo que comprender sus posibles efectos sobre procesos ecológicos es fundamental para desarrollar acciones de conservación (Escribano et al., 2017). Múltiples estudios han evaluado el rol de las cabras como dispersores, haciendo especial énfasis en el paso de las semillas por el tracto digestivo (Miller, 1995; Kneuper et al., 2003; Baraza & Valiente-Banuet, 2008; Mancilla-Leytón et al., 2015; Tjelele, Ward, & Dziba, 2015). Nuestros resultados muestran que en los BTES las cabras están dispersando principalmente semillas de leguminosas, sin embargo, los efectos sobre la germinación son variables entre especies. Aunque inicialmente estos hallazgos, muestran que estos animales pueden estar jugando un rol importante en la dispersión de semillas de algunas especies, la frecuencia de semillas en las fecas de cabras no está asociada necesariamente a la disponibilidad de ciertas especies vegetales en el bosque, presentando una alta frecuencia de semillas de A. macracantha, una especie asociada a los asentamientos humanos.
Las especies encontradas en las fecas fueron en su mayoría especies que tienen semillas con una testa dura y que pueden resistir el paso por el tracto digestivo (Jara-Guerrero et al., 2018). Al menos un 50 % de esas especies son leguminosas, todas con vainas indehiscentes, pero, además registramos la presencia de semillas de C. iguanaea y Z. thyrsiflora, ambas con frutos tipo drupa. El paso por el tracto digestivo mostró un efecto diferencial sobre la germinación, con efectos positivos para tres de las cinco especies evaluadas, las cuales germinaron más rápido y en una mayor proporción que las semillas del tratamiento control. Este efecto diferencial fue evidente incluso entre las leguminosas, lo cual puede atribuirse a diferencias en el tamaño de las semillas. Las especies que no germinaron tienen un tamaño dos veces menor a las otras especies evaluadas. Este patrón es opuesto a lo encontrado en O. virginianus (Jara-Guerrero et al., 2018), donde las especies con semillas más grandes mostraron un efecto negativo. Algunos trabajos proponen que las especies dispersoras podrían retener las semillas dependiendo de su tamaño, de manera que durante el proceso digestivo algunas especies retienen por más tiempo semillas grandes, mientras otras especies retienen más tiempo semillas pequeñas (Picard et al., 2015). Nuestros resultados sugieren que, al contrario de O. virginianus, las cabras podrían estar reteniendo más tiempo semillas pequeñas. Sin embargo, es necesario desarrollar estudios que evalúen las relaciones entre tiempos de retención, tamaño de semilla y grosor de la testa para una mejor compresión de los efectos del proceso de digestión de las cabras sobre la germinación de semillas.
En el caso de A. macracantha, lamentablemente no contamos con un tratamiento control para la germinación, sin embargo, otros estudios señalan porcentajes de alrededor del 5 % (Hernández de Bernal et al., 2011; Maldonado-Arciniegas, Ruales, Caviedes, Ramírez, & León-Reyes, 2018). En nuestro estudio encontramos un porcentaje de germinación del 16 % en las semillas dispersadas por cabras, lo que sugiere un efecto positivo del paso por el tracto digestivo sobre la germinación. Las semillas de esta especie tienen un tamaño similar a las semillas de C. mangense (afectada negativamente), sin embargo, algunos estudios proponen que el tamaño de la semilla no siempre puede explicar los efectos diferenciados del paso por el tracto digestivo, y que los cambios en la impermeabilidad de la semilla podrían estar afectando la germinación (Picard et al., 2015). De esta forma, es posible que A. macracantha tenga una testa más impermeable que le permite sobrevivir al paso por el tracto digestivo. La alta dominancia de semillas dispersadas de A. macracantha, así como el probable efecto positivo sobre la germinación, ha sido observado en varios estudios, donde el género Acacia ha mostrado efectos consistentes de mejorar de la germinación y dominancia en la dispersión por ganado (Miller, 1995; Rohner & Ward, 1999; Bodmer & Ward, 2006; Tjelele et al., 2015).
El alto consumo de leguminosas por parte de ungulados se ha reportado también en estudios previos realizados en BTES del Neotrópico, tanto para cabras (Osorno-Sánchez, 2005; Baraza & Valiente-Banuet, 2008) como para Odocoileus virginianus (Jara-Guerrero et al., 2018). Estas especies han sido clasificadas como especies de frutos anacrónicos (Janzen & Martin, 1982; Jara-Guerrero et al., 2018), cuya dispersión depende en gran medida de grandes vertebrados. En ausencia de agentes dispersores el proceso de dispersión se puede dar por gravedad, permaneciendo la mayoría de las semillas bajo el dosel del árbol parental (Guimaraes et al., 2008). Sin embargo, muchas leguminosas son intolerantes a la sombra, además, la depredación de semillas por insectos podría verse incrementada por la alta densidad de semillas, por lo que las probabilidades de germinación bajo el parental serían bajas (Miller & Coe, 1993). De esta forma, los dispersores incrementan, tanto las probabilidades de encontrar un microhábitat propicio para el desarrollo lejos del parental, como la probabilidad de germinación (Schupp et al., 2010; Jara-Guerrero et al., 2018). En áreas de BTES donde las poblaciones de O. virginianus se han visto reducidas, el papel de las cabras como dispersoras de larga distancia puede ser clave para la regeneración del bosque, pues están dispersando alrededor de un 12 % de las especies leñosas registradas para el área de estudio (Cueva-Ortiz et al., 2019).
Por otro lado, nuestros resultados muestran que la dispersión de semillas por cabras es independiente de la estructura y composición de la vegetación establecida. Las especies dispersadas mostraron una alta dominancia de semillas de especies poco comunes en el bosque, lo cual, sumado a la baja disimilitud en la riqueza de especies dispersadas entre sitios, puede desencadenar cambios en la estructura de la vegetación (Kneuper et al., 2003; Albert et al., 2015). El pastoreo crónico puede afectar la equitatividad, promoviendo y manteniendo unas pocas especies dominantes en las áreas degradadas, generando una homogeneización de la vegetación en el paisaje (Quisehuatl-Medina, Averett, Endress, & Lopez-Toledo, 2020). Procesos similares se han reportado en otros ecosistemas secos, como el matorral seco interandino, donde el ganado ha contribuido a la homogeneización del banco de semillas (Espinosa et al., 2013). Adicionalmente, en un estudio experimental realizado previamente en nuestra zona de estudio, se registró un incremento en la dominancia de A. macracantha en parcelas con exclusión de ganado y adición de fecas de cabra (Rodriguez, 2006). Los cambios en la estructura y dominancia del bosque producidos por la dispersión de semillas de las cabras podrían ser aún más importantes si consideramos sus efectos negativos sobre la supervivencia de la regeneración en los BTES. En un estudio realizado por Cueva-Ortiz et al., (2020), se encontró que la probabilidad de supervivencia de la regeneración natural está fuertemente asociada con el acceso de las cabras al bosque. Pero además, el ganado puede generar cambios en las condiciones del suelo (Metha, Sullivan, Walter, Krishnaswamy, & DeGloria, 2008) que a largo plazo pueden afectar negativamente el crecimiento de árboles y las características estructurales de la vegetación (Breceda, Ortiz, & Scrosati, 2005; Quisehuatl-Medina et al., 2020).
En conclusión, las cabras pueden mejorar la germinación de algunas especies con semillas de testa dura, supliendo en cierta medida el rol de ungulados silvestres en áreas degradadas de BTES donde sus poblaciones se han reducido o desaparecido. No obstante, su comportamiento alimenticio y la destacada predominancia en la dispersión de ciertas especies pueden tener importantes efectos en la estructura de la comunidad vegetal. De esta forma, el rol como dispersor debería ser evaluado en conjunto con los efectos sobre la regeneración natural y estructura de la vegetación. Considerando que, en los BTES del Neotrópico, la ganadería es una de las principales actividades de subsistencia, el conocimiento sobre sus efectos positivos y negativos para la regeneración del bosque es clave al momento de plantear alternativas de manejo que puedan ser acogidas por las poblaciones locales. Posiblemente los efectos negativos producidos por las cabras están asociados a la carga ganadera que soportan estos ecosistemas. Hasta donde sabemos, no existen estudios que evalúen los efectos de diferentes cargas ganadera en este ecosistema, sin embargo, es claro que se necesita generar acciones de exclusión rotativa para facilitar la recuperación de la vegetación. También, se podría aprovechar la capacidad de dispersión de las cabras en áreas defaunadas para iniciar procesos de restauración.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

