











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
La condición nutricional puede ser usada en el manejo de poblaciones naturales de importancia económica (Cifuentes et al., 2012). También puede ser útil en la predicción del crecimiento de las especies, su estado de salud y condición reproductiva (Bonilla-Flórez, Mayer, Estruch-Fuster, & Jover-Cerdá, 2017). Los índices de condición nutricional son estimadores morfofisiológicos que cuantifican el estado nutricional, que, en el caso de crustáceos marinos, ha sido definido por Moore, Smith, y Loneragan (2000) como la magnitud en que los organismos han acumulado las reservas energéticas que garantizan la realización exitosa de las funciones vitales. Como sugirieron Lopeztegui-Castillo, Capetillo-Piñar, y Betanzos-Vega (2012), en el hábitat natural, el estado nutricional deficiente de sólo un ejemplar puede deberse a causas individuo-específicas, como fallas genéticas (que condicionen insuficiencias metabólicas o vulnerabilidad a enfermedades) y la exposición individual a factores ambientales adversos. Por el contrario, un estado nutricional insatisfactorio en un número elevado de ejemplares es la expresión de la influencia generalizada de factores desfavorables.
Entre los numerosos métodos que existen para estimar la condición nutricional, destacan aquellos no destructivos (no requieren de la muerte de los ejemplares) debido a que, además de contribuir a preservar la biodiversidad, resultan eficientes en el caso de organismos de interés comercial. Los índices involucrados por tales métodos son frecuentemente utilizados, entre ellos pueden citarse el Índice de Refracción de la Hemolinfa, como estimador analítico, y la proporción entre el peso del cuerpo y la longitud (del cuerpo o del cefalotórax), como estimador morfométrico (Oliver, & MacDiarmid, 2001; Briones-Fourzán, Baeza-Martínez, & Lozano-Álvarez, 2009). Sin embargo, debido a las diferencias entre las medidas y procedimientos involucrados, se ha argumentado que cada uno de estos índices responde de distinta manera y es sensible a diferentes tipos de factores (Moore et al., 2000; Oliver, & MacDiarmid, 2001).
En la región del Caribe, la langosta espinosa Panulirus argus (Latreille, 1804) es uno de los productos de mayor valor económico (Baisre, 2018). En Cuba, el golfo de Batabanó es la zona de la que anualmente se extrae el mayor porcentaje de la captura nacional, que ha disminuido aproximadamente a la mitad del máximo histórico (Alzugaray-Martínez et al., 2018). En este golfo se ha reportado deterioro ambiental debido principalmente a la influencia de huracanes (Capetillo-Piñar, Espinosa-Sáez, Tripp-Valdez, & Tripp-Quezada, 2016) y estresores antropogénicos (Puga-Millán et al., 2013; Piñeiro-Soto, Areces-Mallea, Puga-Millán, Cobas-Gómez, & de León-González, 2017), todo lo cual ha condicionado el decrecimiento de las comunidades bentónicas (Arias-Schreiber et al., 2008; Capetillo-Piñar, Villalejo-Fuerte, & Tripp-Quezada, 2015) incluyendo una disminución del 82.7 % en el potencial alimentario disponible para las langostas de las zonas más próximas a la costa (Lopeztegui-Castillo & Capetillo-Piñar, 2008), cambios en la dinámica de los sedimentos y pérdidas en la cobertura y densidad de la vegetación submarina (Cerdeira-Estrada, Lorenzo-Sánchez, Areces-Mallea, & Martínez-Bayón, 2008).
Los organismos bentónicos que habitan los fondos blandos del golfo de Batabanó y de otras áreas pesqueras de la región del Caribe, constituyen la base alimentaria de P. argus y otras langostas de importancia comercial (Herrera et al., 1991; Martínez-Coello, Lopeztegui-Castillo, & Amador-Marrero, 2015; Briones-Fourzán et al., 2019). El deterioro ambiental en los hábitats en que reside P. argus puede provocar el decrecimiento en las comunidades bentónicas, fundamentalmente aquellas que constituyen alimento para las langostas, y condicionar cambios fisiológicos y disminución en la condición nutricional (Briones-Fourzán et al., 2019; Lopeztegui-Castillo & Martínez-Coello, 2020). En este contexto, fueron objetivos del presente estudio, determinar variaciones espaciotemporales en el estado nutricional de ejemplares comerciales de P. argus en el golfo de Batabanó, relacionarlas con variaciones reportadas en la comunidad bentónica, y establecer, mediante la aplicación de indicadores analíticos y morfométricos, que la variación de factores ambientales, categorizada como deterioro en este golfo, afecta a ambos grupos de índices debido a su potencial magnitud y su carácter generalizado.
Materiales y métodos
Área de estudio: El golfo de Batabanó, ubicado en la plataforma suroccidental de Cuba, es considerado una macrolaguna con 6 m de profundidad media (Alcolado, 1990; Cruz et al., 1990). La mayor parte de los fondos son fangosos, parcialmente cubiertos de vegetación submarina con predominio de Thalassia testudinum (K.D. Koenig, 1805) de densidad entre media y baja. Basado en la actual distribución de las subzonas de pesca, se registró información de seis áreas: Cabezo, Medios, Alonso, Traviesa, Diego y Matías (Fig. 1).
Fig. 1 Área de estudio con la ubicación de las subzonas de pesca, que se corresponden con los puntos de donde se extrajeron las langostas para muestreo de condición nutricional en el período 2011-2017. / Fig. 1. Study area with the location of the fishing sub-zones, which correspond to the points from where the lobsters were extracted for sampling of nutritional status in the period 2011-2017.
En la escala temporal, los datos se agruparon en tres períodos: “Buesa” (1963-1964), años 1963 y 1964; “Histórico” (1983-1993), años 1983, 1988 y 1993; y “Actual” (2011-2017), años 2011 y desde 2014 hasta 2017. En el período Histórico, se reportaron los mayores volúmenes de captura de esta pesquería (Baisre, 2018). En 1963-1964, la pesquería se consideró en fase de crecimiento, por lo que se asume que las condiciones del hábitat no resultaban limitantes (Puga-Millán et al., 2013). En el período Actual se han reportado afectaciones ambientales y una menor abundancia de langostas (Baisre, 2018; Puga-Millán et al., 2018). Como evidencias cuantificadas de deterioro ambiental, se utilizaron las variaciones espaciotemporales de la comunidad megazoobentónica (organismos mayores de 4 mm), de manera que los valores de densidad (DB) y biomasa (BB) de megazoobentos, y de densidad (DM) y biomasa (BM) de moluscos megazoobentónicos, reportados por Lopeztegui-Castillo y Martínez-Coello (2020) se correlacionaron con el valor de los índices nutricionales en la escala espacial (entre sitios) para los períodos Histórico y Actual, y en la escala temporal agrupando los datos existentes en ambos períodos (entre 1983 y 2017).
Índices de condición nutricional: La condición nutricional se estimó mediante tres índices no destructivos, poco costosos y de fácil y rápida aplicación. Tales índices fueron definidos en la presente investigación como “índices extensivos”, ya que, además, una vez adquiridos los instrumentos iniciales de medición (vernier, balanza, refractómetro de proteínas, cinta métrica) pueden determinarse para un número elevado de ejemplares (tantos como se desee) sin que cada nueva medición implique nuevos gastos económicos o involucre costos biológicos. Se analizaron ejemplares comerciales aparentemente saludables, con la totalidad de sus atributos morfológicos y sin signos externos de muda o reproducción. Dichos ejemplares fueron muestreados, medidos, sexados y pesados in situ, según los criterios de Cruz (2002), y luego fueron entregados vivos a la industria para la continuación del proceso productivo.
El índice de refracción de la hemolinfa (IRH), directamente proporcional a la concentración total de proteínas en el suero sanguíneo (Oliver, & MacDiarmid, 2001), se determinó, expresado en gramos por decilitros (g/dl), para el período 2011-2017. Inmediatamente después de la captura de los ejemplares, con redes de mano (chinchorros langosteros), se extrajo la hemolinfa del seno pericárdico de las langostas, ubicado en posición media dorsal dentro del cefalotórax, siguiendo los criterios de Musgrove (2001). La hemolinfa, extraída con jeringas estériles (una por ejemplar) de 3 cc, se depositó sobre el prisma de un refractómetro de mano modelo Fisher Brix (0-50 %) para realizar, a temperatura ambiente, la lectura del índice de refracción.
El índice morfométrico Klt, se calculó, para todos los años de muestreo, mediante la proporción Peso total real (Ptr) / Largo total (Lt), utilizada antaño por Buesa (1965) y recientemente empleada por Lopeztegui-Castillo et al. (2012) luego de que Lopeztegui-Castillo, Capetillo-Piñar, y Betanzos-Vega (2010) demostraran que Klt puede utilizarse como indicador eficaz de condición nutricional en langostas. El peso de los ejemplares se determinó con una balanza analítica de 0.1 g de precisión. Como medida de longitud se registró el largo total antenular, medido ventralmente desde el punto medio entre la base de las dos antenas hasta el punto medio posterior del telson, para lo cual se utilizó una cinta métrica de 0.1 mm de precisión (Cruz, 2002).
Otro índice morfométrico (FCA), correspondiente al factor de condición de LeCren (Weatherley, 1972), generalmente utilizado para estimar estado nutricional en peces, fue calculado siguiendo los criterios de Lozano-Álvarez y Aramoni-Serrano (1996), quienes lo aplicaron a dos especies de palinúridos en aguas de México. La formulación empleada fue FCA = Ptr/Pte, donde Ptr es el peso total real (observado) de las langostas y Pte el peso total (teórico) estimado a partir de una ecuación específica para cada período, según recomendaron Lopeztegui-Castillo et al., (2010), quienes encontraron diferencias significativas entre los parámetros de grupos de langostas muestreados en diferentes épocas y regiones. Para el período 1963-1964 se empleó la ecuación Pte = 1.5653 x 10-4 x Lt 2.7102, propuesta por Buesa (1965). En el período Histórico (1983-1993) la ecuación empleada fue Pte = 0.5 x 10-4 x Lt 2.8957. Para los datos del período Actual (2011-2017) se utilizó la ecuación Pte = 0.5 x 10-4 x Lt 2.8579.
Organización de los datos: Para determinar las variaciones temporales a largo plazo (60 años) de la condición nutricional, se emplearon los índices morfométricos Klt y FCA. Se utilizó la información reportada por Buesa (1965) para el período 1963-1964 (N = 29 001) y la información del período “Histórico” 1983-1993 (N = 3 123), existente en los archivos del Centro de Investigaciones Pesqueras. Sólo los datos registrados en el período “Actual”, 2011-2017 (N = 3 600), incluyeron mediciones de IRH, las que fueron utilizadas para determinar variaciones temporales a corto plazo (entre 2011 y 2017), que también se determinaron para los índices morfométricos. Para el análisis del período Actual (variaciones temporales a corto plazo), los datos se separaron en épocas de Lluvia (de mayo a octubre) y Seca (de noviembre a abril), según los criterios de García-Rodríguez, Durán-Llacer y Lapinel-Pedroso (2020). El muestro de langostas se realizó en un mes de Lluvia y un mes de Seca.
Procesamiento estadístico: Mediante la prueba de Shapiro-Wilk se comprobó que los datos sobre condición nutricional no cumplen con las premisas de una distribución normal, razón por la que se aplicaron pruebas no paramétricas. En el caso de las comparaciones pareadas (entre épocas de Lluvia y Seca, o entre pares de índices) se empleó la U de Mann-Whitney. Las comparaciones múltiples (entre los tres períodos, entre años y entre sitos) se llevaron a cabo mediante la prueba Kruskal-Wallis (KW-H). El coeficiente de correlación rho de Spearman (rs), se utilizó para estimar la asociación, por pares, entre los índices de condición nutricional determinados. Todos los análisis fueron realizados para un nivel de significación de P < 0.05.
Resultados
En el período Actual, tanto IRH [KW-H(5; 3 600) = 189.1; P = 0.001] como Klt [KW-H(5; 3 600) = 89.6; P = 0.001] mostraron diferencias entre áreas. También en el período Histórico se detectaron diferencias entre sitios, tanto para Klt [KW-H(5; 3 123) = 799.17; P = 0.001] como para FCA [KW-H(5; 3 123) = 581.60; P = 0.001]. La información registrada por Buesa (1965) no permitió realizar comparaciones estadísticas ya que sólo se reportó un valor promedio de Klt por subzona de pesca. Sin embargo, al graficar las medidas de tendencia central (medianas) en los tres períodos, se evidenció que las variaciones de Klt entre subzonas no son semejantes entre períodos (Fig. 2).
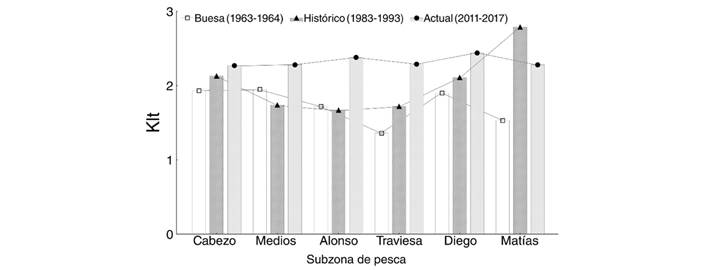
Fig. 2 Variación espacial del índice morfométrico Klt (proporción peso total / largo total) estimado para langostas Panulirus argus. Períodos Actual (2011-2017), Histórico (1983-1993) y Buesa (1963-1964). / Fig. 2. Spatial variation of the Klt (total weight / total length ratio) morphometric index, estimated for Panulirus argus lobsters. Actual (2011-2017), Historical (1983-1993) and Buesa (1963-1964) periods.
Los tres índices de condición nutricional detectaron diferencias entre años del período Actual, con los valores más bajos en el 2017. Sin embargo, mientras que IRH decrece gradual y significativamente [KW-H(4;3 600) = 155.0; P = 0.001] entre 2011 (16.6) y 2017 (14.2), Klt [KW-H(4;3 600) = 170.1; P = 0.001] y FCA [KW-H(4;3 600) = 26.6; P = 0.001] tienden a aumentar hasta 2016 y sólo en 2017 decrecieron incluso a valores inferiores a los encontrados en 2011 (Fig. 3).
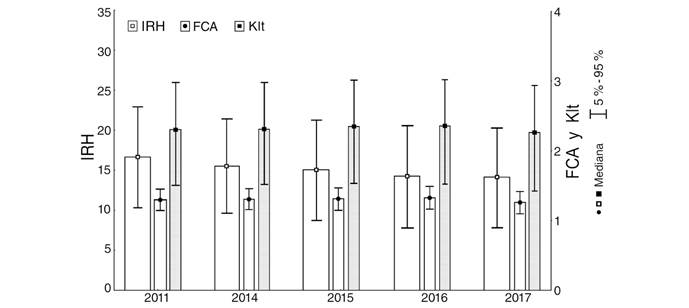
Fig. 3 Variaciones temporales a corto plazo (período Actual 2011-2017) de los índices de condición nutricional, determinados sobre ejemplares comerciales de Panulirus argus. IRH: índice de refracción de la hemolinfa; Klt: proporción peso total / largo total del cuerpo de las langostas; FCA: factor de condición. / Fig. 3. Short scale temporal variations (Current period 2011-2017) of the nutritional condition indices determined on commercial Panulirus argus lobsters. IRH: blood refractive index; Klt: total weight / total length ratio; FCA: condition factor.
El peso total de las langostas y el índice Klt no mostraron diferencias entre Seca y Lluvia. Sin embargo, tanto IRH como FCA resultaron estadísticamente mayores en Seca, hecho que se constató para cada sexo (Tabla 1).
Tabla 1 Comparación entre Seca y Lluvia, de los índices de condición nutricional y el peso total real (Ptr) de hembras, machos y el total de langostas Panulirus argus, durante el período Actual (2011-2017). Table 1. Comparison, between dry and rainy seasons, of the nutritional condition indices and the total weight (Ptr) of females, males and the total of Panulirus argus lobsters, during 2011-2017 period
| Índice | Machos N Seca = 1173; N Lluvia = 1052 | Hembras N Seca = 628; N Lluvia = 747 | Total N Seca =1800; N Lluvia = 1800 | |||
| M-W (Z) | P (< 0.05) | M-W (Z) | P (< 0.05) | M-W (Z) | P (< 0.05) | |
| Ptr (g) | -0.47 | 0.641 | 0.06 | 0.951 | 0.15 | 0.881 |
| Klt | -1.32 | 0.185 | -0.81 | 0.416 | -0.83 | 0.406 |
| FCA | -3.77 | 0.001* | -4.14 | 0.001* | -4.11 | 0.001* |
| IRH | 3.23 | 0.001* | 2.09 | 0.037* | 4.16 | 0.001* |
FCA = factor de condición; IRH = índice de refracción de la hemolinfa; Klt = proporción peso total / largo total. / FCA = condition factor; IRH = blood refractive index; Klt = real total weight / total length ratio.
Los índices FCA y Klt no presentaron diferencias entre el período Histórico (1983-1993) y lo reportado por Buesa (1965) para 1963-1964. No obstante, tanto FCA [KW-H(2;4 502) = 215.3; P = 0.001] como Klt [KW-H(2;4 502) = 361.0; P = 0.001] aumentaron en el período Actual (2011-2017) respecto al Histórico. Sólo en el caso de FCA este aumento fue significativo incluso respecto a lo reportado para el período 1963-1964 (Fig. 4).
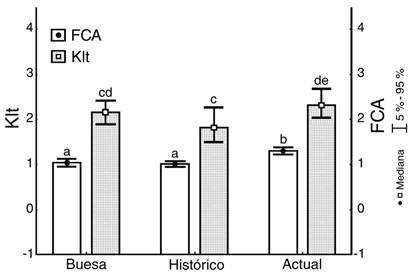
Fig. 4 Variaciones temporales a largo plazo (entre períodos, desde 1963 hasta 2017) de los índices morfométricos de condición nutricional, para ejemplares comerciales de Panulirus argus. Klt: proporción peso total / largo total del cuerpo de las langostas; FCA: factor de condición. / Fig. 4. Long scale temporal variations (among periods, from 1963 until 2017) of the morphometric indices of nutritional condition for commercial Panulirus argus lobsters. Klt: total weight / total length ratio; FCA: condition factor.
El coeficiente rho de Spearman entre los diferentes índices nutricionales, y entre éstos y el peso de las langostas, corroboró que, aunque todas las correlaciones fueron significativas, los coeficientes más altos se establecieron entre los índices morfométricos. Sin embargo, la correlación de los índices nutricionales con los valores de densidad (DB) y biomasa (BB) de bentos, y particularmente de densidad (DM) y biomasa (BM) de moluscos bentónicos, mostró que, en la escala temporal, sólo el IRH presentó elevados coeficientes, significativos en tres de los casos (Tabla 2). En la escala espacial (entre sitios) ningún coeficiente resultó significativo o superior al rs = 0.6001 encontrado entre Klt y DB.
Tabla 2 Coeficiente de correlación rho de Spearman (rs) entre índices de condición nutricional, entre los índices y el peso total real (Ptr) de ejemplares comerciales de Panulirus argus capturados durante los períodos 1983-1993 (H) y 2011-2017 (A), y entre estos indicadores y los valores de densidad y biomasa, de megazoobentos y de moluscos megazoobentónicos. Table 2. Spearman’s rho correlation coefficient (rs) between nutritional condition indices, between each index and the real total weight (Ptr) of commercial Panulirus argus lobster, captured during 1983-1993 (H) and 2011-2017 (A) periods, and between such indicators and the density and biomass values for megazoobenthos and for megazoobenthic mollusks
| Par de variables | rs | Par de variables | rs | Par de variables | rs |
| FCA H-Klt H | 0.7142* | DB-Klt | -0.2571 | DM-Klt | 0.0857 |
| FCA H-Ptr H | 0.6756* | DB-FCA | 0.1160 | DM-FCA | -0.2899 |
| FCA A-Klt A | 0.5871* | DB-IRH | 0.9013* | DM-IRH | 0.8000 |
| FCA A-IRH A | 0.3095* | DB-Ptr | -0.6057 | DM-Ptr | -0.3714 |
| IRH A-Klt A | 0.1907* | BB-Klt | -0.4857 | BM-Klt | -0.0286 |
| IRH A-Ptr A | 0.1655* | BB-FCA | 0.2609 | BM-FCA | -0.1739 |
| FCA A-Ptr A | 0.3194* | BB-IRH | 0.9097* | BM-IRH | 0.9009* |
| Klt A-Ptr A | 0.9862* | BB-Ptr | -0.7143 | BM-Ptr | -0.6001 |
DB = densidad de megazoobentos; BB = biomasa de megazoobentos; DM = densidad de moluscos megazoobentónicos; BM = biomasa de moluscos megazoobentónicos, reportados por Lopeztegui-Castillo & Martínez-Coello (2020) para el período Actual. / DB = density of megazoobenthos; BB = biomass of megazoobenthos; DM = density of megazoobenthic mollusks; BM = biomass of megazoobenthic mollusks, reported by Lopeztegui-Castillo & Martínez-Coello (2020) in the Current period. *: significación estadística (P < 0.05) / Significantly (P < 0.05).
Discusión
Klt presentó variaciones espaciales que no coincidieron entre períodos, lo que sugiere que los factores que causan cambios en el estado nutricional de P. argus influyen estocásticamente en cualquiera de las áreas, lo cual es más característico de los factores ambientales que de los antropogénicos. Una evidencia de esto es que la correlación espacial entre los índices nutricionales y la densidad y biomasa, de bentos y de moluscos bentónicos, no fue elevada ni significativa en ningún período. En el golfo de Batabanó, el impacto que factores ambientales con tales características de estocasticidad, como son los huracanes, han tenido sobre las comunidades bentónicas, ha sido ya descrito (Capetillo-Piñar et al., 2016). No obstante, el inevitable impacto de la pesca en los hábitats naturales podría contribuir a la afectación de las comunidades bentónicas e influir en la condición nutricional, por lo que se recomiendan pormenorizados estudios al respecto.
Las áreas con mayores valores de Klt (Matías en período Histórico y Diego en Actual) fueron las más próximas al borde de la plataforma, lo que pudiera deberse a que el mayor intercambio con aguas exteriores favorece el mejor desarrollo de la comunidad bentónica (Martínez-Daranas, Betanzos-Vega, Lopeztegui-Castillo, & Capetillo-Piñar, 2018). Sin embargo, la presencia de ejemplares de mayor talla y peso es más probable en estos sitios de borde, por donde los grandes reproductores, que usualmente habitan aguas profundas, pueden acceder a la plataforma (Cruz et al., 1990). Es posible que en estos sitios los índices morfométricos estén más influidos por la estructura de pesos y tallas de la población de langostas que por la disponibilidad de alimento.
A pesar de la falta de coincidencia entre períodos respecto a las variaciones espaciales de la condición nutricional, Klt aumentó gradualmente (Buesa < Histórico < Actual) en tres áreas (Cabezo, Traviesa y Diego). Aunque FCA y Klt resultaron similares en los períodos Buesa e Histórico, ambos alcanzaron sus mayores valores en el período Actual. Debido a que el deterioro de la comunidad bentónica en el golfo de Batabanó sugiere que esto no se relaciona con el aumento de los recursos alimenticios (Arias-Schreiber et al., 2008; Capetillo-Piñar et al., 2015; Lopeztegui-Castillo & Martínez-Coello, 2020), este aumento de los índices morfométricos se atribuye al aumento del peso respecto a la talla de las langostas. El carácter morfométrico de ambos índices los hace susceptibles a la presencia de individuos de mayor peso y talla, lo cual puede ser resultado de la elevada presión pesquera, que puede hacer variar la composición por tallas de la población (de León, 2005; de León, Puga-Millán, & Piñeiro-Soto, 2010). La posibilidad de que langostas de mayor talla ingresen a áreas de pesca desde aguas más profundas no puede descartarse, principalmente en áreas próximas al borde de la plataforma (Cruz, 2002).
Las variaciones temporales a corto plazo demostraron que los tres índices disminuyeron en 2017. Sin embargo, durante 2011-2016, los índices morfométricos aumentaron, de 1.31 a 1.33 el FCA y de 2.25 a 2.36 el Klt. Este aumento se contradice con la disminución presentada por IRH en igual período, lo que se explica por las características de cada índice. El menor valor de Klt significa que los ejemplares pesan menos respecto a su talla, lo que se traduce en que langostas de similar tamaño tuvieron menor peso en 2017. Un IRH menor es indicativo de una menor concentración de proteínas en la hemolinfa, lo que pudiera deberse a la menor disponibilidad de alimento en las zonas de pesca, como sugiere la elevada y significativa correlación que, en la escala temporal, se encontró entre IRH y la densidad y biomasa de bentos y de moluscos bentónicos.
Cuando los índices morfométricos se basan en proporciones que incluyen medidas de peso y longitud, como es el caso de Klt, la detección de variaciones significativas se limita a dos posibilidades: 1) los factores estresantes, perecederos o no, son suficientemente drásticos como para hacer variar el peso de los ejemplares en un tiempo relativamente corto (menor a la duración de un ciclo de muda); 2) los factores estresantes no son drásticos pero permanecen lo suficiente como para hacer variar el peso y la talla en diferente proporción (Lopeztegui-Castillo et al., 2012). Este segundo caso, opción más probable en el golfo de Batabanó, requiere que los ejemplares muden una o más veces, lo que puede demorar entre un mes y varios años dependiendo de, entre otros aspectos, la magnitud de los factores estresantes y la frecuencia de muda. Aunque los índices morfométricos resultaron mayores en el período Actual, los menores valores en 2017 sugieren que las actuales condiciones de deterioro, ya probadas en el golfo de Batabanó, tienen la magnitud adecuada y han persistido lo suficiente como para comenzar a afectar a los índices morfométricos.
El IRH, indicador analítico que manifiesta la concentración de proteínas al instante de la observación, puede detectar el impacto que, a corto plazo (incluso menos de una hora) tienen las variaciones ambientales sobre el estado nutricional de los organismos (Wang & McGaw, 2014). Las características intrínsecas a las determinaciones mediante IRH, hacen posible que detecte, más rápidamente que Klt y FCA, las variaciones metabólicas provocadas por pequeños cambios en la disponibilidad de alimento o factores abióticos como la temperatura (Wang, Robertson, Wringe, & McGaw, 2016). La gradual tendencia a la disminución en los valores del IRH, aún en áreas cercanas al borde de la plataforma, sugiere el carácter generalizado del deterioro ambiental, recientemente evidenciado también en pérdidas de vegetación submarina (Martínez-Daranas et al., 2018).
El coeficiente de Spearman fue mayor entre índices morfométricos y menor entre estos y el IRH, lo cual corrobora que los índices morfométricos y analíticos expresan información diferente y detectan cambios que ocurren en distintas escalas de tiempo (Oliver & MacDiarmid, 2001). El coeficiente de correlación de mayor magnitud se encontró entre Klt y Pt, lo que sugiere que este índice de condición nutricional es el que más se corresponde con lo que comercialmente se utiliza como marcador óptimo de buena condición en las langostas: el peso (Lopeztegui-Castillo & Amador-Marrero, 2015). Sin embargo, es posible que una langosta de elevado peso (condición óptima para el comercio) no esté fisiológicamente sana o presente una condición nutricional no totalmente satisfactoria, por lo que estos resultados deben interpretarse de forma cuidadosa.
Lozano-Álvarez y Aramoni-Serrano (1996), reportaron que el FCA estimado para Panulirus inflatus (Bouvier, 1895), presentó variaciones estacionales significativas, con mayores valores en invierno. Resultados similares se obtuvieron en el presente estudio mediante IRH y FCA, que resultaron mayores en época de Seca (meses de invierno), tanto para hembras como para machos. Este hecho pudiera deberse a la disminución de la actividad metabólica como consecuencia de la baja temperatura de las aguas en esta época del año (Rodríguez-Fuentes et al., 2017). Otra posible explicación es que los organismos estén acumulando las sustancias de reserva que servirán para enfrentar el proceso reproductivo, que en el caso de P. argus tiene su pico de máxima actividad entre febrero y junio (Cruz et al., 1990; de León, 2005).
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

