











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
Temporal and spatial variation in the diversity and composition of the pollinator community of a plant species is a frequent conclusion of pollination studies (Herrera, 1989; Waser, Chittka, Price, Williams, & Ollerton, 1996; Price, Waser, Irwin, Campbell, & Brody, 2005; Winfree et al., 2018), although the amplitude of this variation may be restricted to the taxonomic groups favored by the floral syndrome (Rosas-Guerrero et al., 2014). This variation is an important condition for evaluating selection pressures on floral traits (Santiago-Hernández et al., 2019), as well as the diversity of pollinators required for the reproduction of a plant species at a scale larger than a population or locality (Winfree et al., 2018). The quantity and diversity of pollinators plays an important role in the productivity of crops worldwide, particularly in small farms aimed at the production of fruits and vegetables (Hoehn, Tschamtke, Tylianakis, & Steffan-Dewenter, 2008; Garibaldi et al., 2016; Reilly et al., 2020).
The cultivated species of the genus Cucurbita (Cucurbitaceae) (pumpkin or squash; Spanish common names include calabaza, ayote, zapallo, and pipian) comprise a group of five species with a proposed center of origin in North America that have been developed through the domestication of a collection of wild species (Hurd, Linsley, & Whitaker, 1971). The species of this genus are characterized by being monoecious, with large seasonally produced yellow flowers in which both the staminate and pistillate flowers produce large amounts of nectar. Nectaries are found in a chamber at the base of the stamens in staminate flowers and in a lower ring at the base of the pistil in pistillate flowers (Nepi & Paccini, 1993; Nepi, Guarnieri, & Pacini, 2001; Vidal, Jong, Wien, & Morse, 2006). Therefore, Cucurbita species depend on pollinating agents for their reproduction.
Floral visitors of Cucurbita include several species of the subgenera Eucera (Peponapis) and Eucera (Xenoglossa), which are native to America and have evolved specialization in collecting pollen exclusively from Cucurbita sp. (Hurd et al., 1971). These bees have been identified as the most effective pollinators of wild and cultivated species of Cucurbita, and their current distribution range has been attributed to the expansion of the crop (Hurd et al., 1971). However, across the American continent, these crops are visited by highly diverse bee species, whose composition depends on the species of Cucurbita, geographic region and season of the year (Ashworth & Galetto, 2001; Meléndez-Ramírez, Magaña-Rueda, Parra-Tabla, Ayala, & Navarro, 2002; Shuler, Roulston, & Farris, 2005; Krug, Alves-dos-Santos, & Cane, 2010; Serra & Campos, 2010; Vidal, Jong, Wien, & Morse, 2010; Zambrano, Gonzales, Hinojosa-Diaz, & Engel, 2013; Enríquez, Ayala, Gonzales, & Nuñez-Farfán, 2015; Phillips & Gardiner, 2015, Delgado-Carrillo et al., 2018; Pfister, Eckerter, Schirmel, Cresswell, & Entling, 2017). A review of bee inventories of C. pepo, C. moschata and C. maxima flowers (Digital Appendix) shows that nonnative Apis mellifera (nonnative), Eucera and several species of Bombini, Meliponini and Halictidae are the most common groups of bees visiting Cucurbita crops on this continent.
Some studies have analyzed the geographic diversity of the abundance and composition of Cucurbita crop-associated bees showing that factors such as changes in forest cover, the type of management, the area of cultivation, and altitude influence the spatial variation of these bee communities (Meléndez-Ramírez et al., 2002; Shuler et al., 2005; Krug et al., 2010; Serra & Campos, 2010; Vidal et al., 2010; Zambrano et al., 2013; Enríquez et al., 2015; Phillips & Gardiner, 2015). Spatial changes in the composition of species that pollinate a crop require the conservation of pollinator diversity at local and regional spatial scales to maintain the ecosystem services provided by local bees at different locations (Winfree et al., 2018), particularly when the pollination services of managed honeybees may be endangered by the decline in their populations (Carreck & Newman, 2010). These conservation efforts represent a challenge for our societies because of the serious threat of decline of bees due to different causes of human origin (Potts et al., 2010).
In Central America, there have been few studies of the bees associated with Cucurbita crops. In Costa Rica, Wille (1985) found 5 species of the genus Eucera in crops from the Central Valley and Guanacaste, but no information was reported for the other bees associated with these crops. In Guatemala, Enríquez et al. (2015) reported 27 species of bees, belonging to Apidae and Halictidae, visiting these crops in highlands regions. These authors conclude that there is high variation between locations in the composition of species but that a fraction of dominant species may generally ensure the pollination of the crop. Both countries are characterized by great altitudinal and climatic heterogeneity within a relatively small territory, presenting ideal conditions for studying geographic variation in the pollination of crops such as squash.
This study was aimed at increasing our knowledge about the community of visiting bees of Cucurbita pepo crops in the Central American region through an inventory of bees in different squash fields in three regions of Costa Rica. The main objective is to determine the taxonomic diversity and composition of bee species visiting Cucurbita pepo crops in different localities in Costa Rica, and their geographic and temporal pattern variation within the study area. These observations would allow us to determine whether the Eucerini species observed by Wille (1985) in Costa Rica can still be found 35 years later. In addition, this study recorded some aspects of intrafloral behavior of dominant species that may be related to pollinator potential. These observations would let us to understand better the importance of bee diversity as a pollinator of this crop in Neotropical regions.
Materials and methods
Study locations: During the years 2016-2019, collections were carried out in Costa Rica in different squash fields in the Central Valley (7 locations), in Guanacaste Province (2 locations) and in the Western sector of the Talamanca mountain range (2 locations). The landscape characteristics, climate and altitude of each location are presented in Table 1. Local climatic conditions were obtained from regional climatic analysis of the Costa Rican National Meteorological Institute (CRRH, 2008). In these locations cultivation of Cucurbita pepo is done without irrigation, which limits the growth and flowering of the crop to the rainy season (May-November). C. moschata, another squash species cultivated in Costa Rica, is grown under the same time schedule of C. pepo in Costa Rica. Flowering peak commonly occurs between May to August, period when sampling of bees was done. The maximum distance between two locations, except Cuajiniquil, Guanacaste, was 50 km. Cuajiniquil is located approximately 190 km from the Central Valley. The minimum distance was 5 km. Most of the localities were grouped in the Central Valley and Cordillera de Talamanca, relatively close to one another, but with important contrasting values of altitude and precipitation (Table 1). Most of the sampling sites were in agricultural regions with high forest fragmentation, where forest patches subsist as gallery forests of different sizes, with the exception of La Cima de Dota and Santa Cruz, where agricultural farms close to protected areas with mature or secondary forests were visited. Another important variable was the number of open flowers at the time of sampling, with localities ranging from small family crops (< 20 open flowers at the time of collection) to farm crops with more than 200 open flowers. The diversity and abundance of bees attracted to these sites may be influenced by the size of the floral display. Wild Cucurbita species or others domesticated Cucurbita crops were not observed near the visited farms.
Table 1 Sampling sites of Cucurbita pepo bees in Costa Rica, with biophysical and landscape characteristics of each location
| Region | Locality | Geographic Coordinates | Altitude (m.a.s.l) | Annual Precipitation (mm) /Rainy days | Dimension of the squash field* | Landscape features |
| Central Valley | Piedades | 9o56’10” N 84o13’26” W | 818 | 1 947/145 | Large | Semiurban/ gallery forest |
| Cerro Colón | 9o54’42” N 84o13’03” W | 1 120 | - | Large | Pastures/ Forest fragments | |
| Acosta | 9o46’ 8” N 84o12’22” W | 760 | 2 370/149 | Medium | Pastures/ Forest fragments | |
| Sta Cecilia | 10° 2’ 5” N 84° 2’17” W | 1 520 | 2 531/152 | Small | Pastures/ Forest fragments | |
| Pacayas | 9o54’24” N 83o47’42” W | 1 760 | 2 245/193 | Small | Pastures/ Crops | |
| Cervantes | 9o53’13” N 83o47’42” W | 1 250 | 2 000/193 | Large | Crops | |
| Juan Viñas | 9o53’37” N 83o44’28” W | 1 200 | 1 675/163 | Small | Pastures/Crops | |
| Guanacaste | Cuajiniquil | 10o56’08” N 85o41’13” W | 15 | 1 517/89 | Medium | Pastures |
| Sta Cruz | 10o10’13” N 85o32’15” W | 675 | 2 116/89 | Small | Crops/ Forest fragments | |
| Talamanca | La Cangreja | 9o47’45” N 83o57’20” W | 1 830 | 1 751/182 | Small | Crops/Pastures/ Fragments forests |
| La Cima | 9o39’34” N 83o54’54” W | 2 146 | 2 632/192 | Large | Crops/Pastures/Primary forests |
* - Small (< 20), medium (20-200), large (> 200 open flowers at the time of collection).
Bee collection and counting: At each site, a collection of bees was obtained from 5:00-9:00 am. Bees were trapped in male and female flowers, although most of the samples were collected in male flowers because of their higher frequency. The squash field was explored for periods of 15 min, during which all the bees observed in flowers were collected except for Apis mellifera, Trigona and Bombus species. These species were common and easy to identify in the field; thus, only the number of individuals in the flowers during each census was registered. The maximum number of individuals counted in a census was used as an estimate of the local abundance of these species. All collections were performed on sunny days. As the composition of species can change with the time of the day, 6 collections were performed, distributed among early (4:45-5:00 am; 5:15-5:30 am), middle (5:45-6:00 am; 6:15-6:30 am) and late hours (6:45-7:00 am; 7:30-8:45 am). The last time interval was a somewhat longer than the others because we observed that species composition remained stable after 7:00 a.m. (see below). It was observed that bee activity declines drastically after 9:00 a.m. At that time, most of the flowers were dominated by Trigona bees with few visits by other species, or there were no visits at all. Some localities (Piedades, Cerro Colón, La Cima de Dota and Sta Cruz) were visited once more approximately 1 week later. No new species were observed in samples obtained in these visits. It was also observed that abundance estimates of each species were approximately repeated on the second survey. It was concluded that the sampling effort was enough to obtain good estimates of bee taxonomic diversity and species composition in the studied localities for the specific flowering period. To homogenize between localities, the abundance data obtained in the first sampling were selected in the localities visited twice. All bees were identified to the species level except Agapostemon and Augochlora. Identification was performed using available taxonomic keys and the collection of the Zoology Museum of the Escuela de Biología, Universidad de Costa Rica, where voucher specimens were deposited. Eucera (Peponapis) specimens were sent to the Bee Biology and Systematics Laboratory (Utah) to help with identification.
In some localities, Piedades and Cerro Colón, there are two squash crops per year, one in the middle (August-September) and another at the end of the rainy season (November-December). To determine if composition of the floral visitors changes between crop seasons, sampling was performed during each crop season in Piedades, one in August 2019 and another in November of the same year, following the same collection intensity protocols and observation times.
Intrafloral behavioral observations: To determine whether the different floral visitors made contact with the reproductive parts of the flowers and to obtain observations of the foraging behavior of each group of bees inside the flowers, continuous video recordings were performed in flowers from 5:00 am until 7:30 am at two localities (Piedades and La Cima de Dota). Sony HDRCX700 Handycam video cameras (San Diego, CA, USA) were used. The recordings ended at 7:30 am because after this time no significant changes were detected in the activity pattern of the bees, except for their near disappearance from the flowers after 10:00 a.m. when many flowers showed signs of wilting. Six male and five female flowers were chosen for filming in La Cima de Dota in August 2016, and the same number of flowers were chosen in the town of Piedades in August 2019. This comparison allowed us to visualize the differences in behavior between localities with different floral visitors (see below). A total of 6 h of video recordings were obtained in each location, which were manually analyzed to check the foraging schedule and the behavior of different bees in each location. Special attention was paid to the pollen load (large = body densely covered by pollen, intermediate = presence of scattered pollen, more visible in corbiclae/escopae; small = pollen scattered with few or no grains) and the visit duration. A sample of 2-3 individuals of bee species observed at flowers were collected and their pollen loads visualized in a microscopy to check if it were Cucurbita pollen.
Statistical analysis: Mean values and confidence intervals of species richness (number of species) for each locality and altitudinal level were obtained by bootstrapping with 300 replicates (Hsieh, Ma, & Chao, 2016). We grouped locations in three altitude categories: low (< 1 000 m.a.s.l. 5 localities), intermediate (1 000-1 800 m.a.s.l. 5 localities) and high (> 1 800 m.s.a.l. 2 localities). A generalized linear model with a Poisson distribution was used to test the effect of altitude on species richness using altitude as a categorical variable. To test differences in the composition (abundance of each species) of the bee community at different altitudes a Multivariate Analysis of Variance with permutations (PERMANOVA) was performed using the Bray-Curtis distance, as implemented in the vegan package (Oksanen et al., 2019).
Results
Diversity and geographic variation: The species collected and their order of dominance (frequency rank order, 1= most frequent, 8= least frequent) at each locality are shown in Fig. 1. Some localities had very low visitation rates (Juan Viñas and Santa Cecilia), which is reflected in the low number of bees collected from those locations. In total, individuals belonging to 19 genera and 2 families (Halictidae and Apidae) of bees were obtained. From the samples identified to the species level, a total of 22 species were obtained, while 5 groups of bees were only identified to the genus level, for a minimum total number of 27 species. The number of species could increase by 2-3 if genera such as Augochlora (very diverse) or Agapostemon were identified to the species level. The presence of 2 species of the genus Megalopta as well as one species of Caenaugochlora (C. costaricensis) at high levels of dominance, was noted at 5 and 4 locations, respectively.
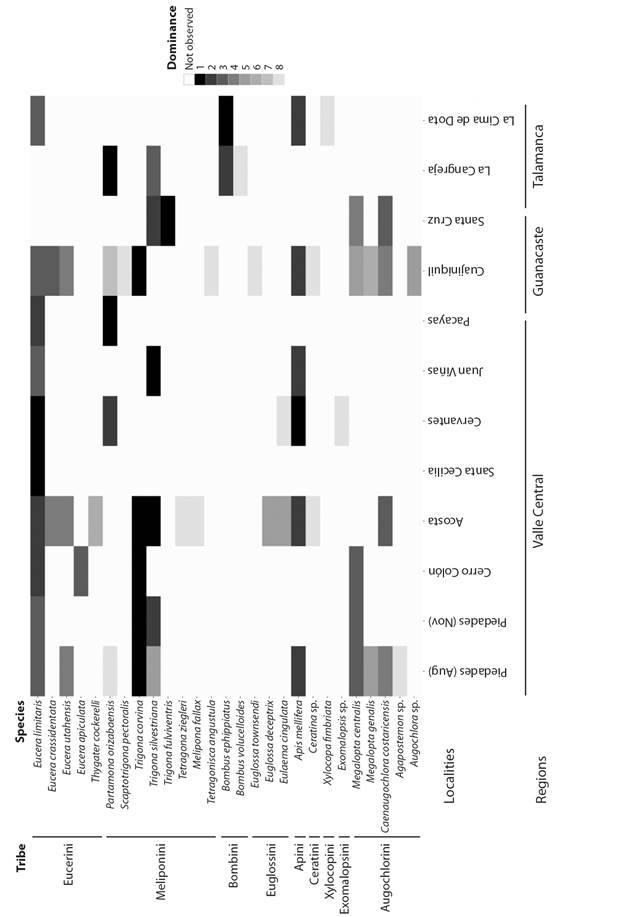
Fig. 1 Bee species in squash crops (Cucurbita pepo) at 11 locations in Costa Rica. The level of shading in each box indicates the order of dominance of each species at each locality, where black indicates the most frequent species (dominance 1). Species with very similar frequencies share the same shading. Level 8 represents rare species (frequently 1-2 individuals observed/collected).
Abundant species were observed in most localities (Fig. 1): 2 species of Meliponini tribe: Trigona corvina and Partamona orizabaensis, honey bees (Apis mellifera) and a species of bee specialized on this crop, Eucera (Peponapis) limitaris. A. mellifera workers observed in the crops are probably originated from feral colonies, as no commercial beekeeping was observed near the studied crops. Other species were frequent only in some locations: i) Bombus ephipiatus was the most frequent floral visitor in La Cima de Dota, the highest-altitude location but was rare or absent in other locations; ii) in Cuajiniquil (Guanacaste) and Acosta, Eucera (Peponapis) crassidentata was frequent. Among the two subgenera of bees specialized on Cucurbita, four species of Eucera (Peponapis) (E. limitaris, E. crassidentata, E. utahensis and E. apiculata) were collected, the last of which was only collected in one locality (Cerro Colón) as a rare species (Fig. 1).
Large differences in species richness were found between localities (Fig. 2). The localities with the highest species richness (species richness 10.5-15.2), were Piedades (Aug), Acosta and Cuajiniquil, which were found at altitudes less than 1 000 m. a. s. l. in medium-large crops and in drier climatic regimes. Small crops (La Cangreja, Juan Viñas, Sta Cecilia, and Pacayas) showed lower species diversity (species richness 1-4), although low diversity was also found in a large crop in highland regions (La Cima de Dota) (species richness 4). Crops located at low altitude levels presented higher species richness compared to medium (Z = -3.897, P <0.001) and high altitudes (Z = -2.174, P < 0.05) PERMANOVA analysis showed that bee species composition changed significantly between altitude levels (F = 2.03, R2=0.31, d.f.= 2, 9, P < 0.01).
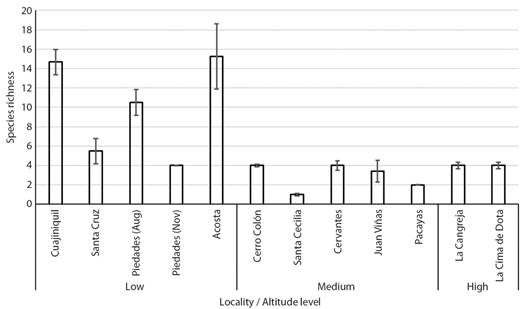
Fig. 2 Species richness of bee populations visiting C. pepo crops in each locality studied. Localities are grouped by altitude level (low 0-1 000 m.a.s.l., medium 1 000-1 800, high > 1 800). Mean values and confidence intervals were obtained by bootstrapping.
At Piedades, the two sequential collections corresponding to the two annual crops separated by 3 months (August and November) showed changes in the abundance and order of dominance of the species (Fig. 1). In November, a frequent species (Apis mellifera) and several rare species (Agapostemon sp., M. genalis, and E. utahensis) observed in August disappeared, reducing the number of species from 10 to 4 as well as the abundance of bees collected in flowers (150 to 45). In November, T. corvina frequency was 95 % of the samples, while in August this figure was approximately 60 %.
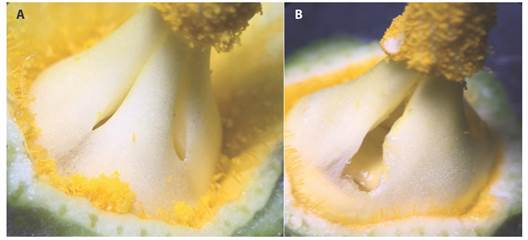
Behavior of common species: Table 2 summarizes the main observations of 5 of the most common floral visitors of C. pepo in the visited localities. All of these species touched the reproductive parts of male and female flowers. The largest and hairiest species (Eucera and Bombus) and those that visit male flowers earlier (Megalopta) were the ones that most frequently carry large pollen loads classified as “large”. A temporal sequence of foraging was observed, in which the crepuscular Megalopta sp. begins at 4:30 am (observed before the initiation of the recordings), followed by the arrival of Apis mellifera, Eucera and Bombus, among which the last genus was only observed in highland areas. Different species of Trigona or Partamona, especially Trigona corvina, arrive somewhat later but quickly invade the area around the nectaries of almost all flowers. Megalopta and Trigona bees tend to spend long times collecting nectar (Table 2). Trigona bees exhibit two interesting foraging behaviors: i) they enlarge the nectar chamber pores with their jaws (Fig. 3) seeking nectar, ii) they tend to repel other bees from flowers, because of their abundance and activity, and on rare occasions, by direct attacks. Eucera (Peponapis), Bombus and A. mellifera are characterized by short visits (16-27 s) and quick flights to flowers (Table 2).
Table 2 Behavioral patterns of the most common bee visitors of C. pepo in Costa Rica obtained from video recordings in two locations. See Methods for the classification of the pollen load and duration of visits
| Bee taxa | Pollen load | Time of foraging | Time of foraging | Median of duration of visit in seconds (s) (interquartile range) | Observations |
| First individuals observed | Last individuals observed | ||||
| Eucera (Peponapis) sp. | Large | 5:00 a.m. | rare after 7 a.m. | 18 (5-62) | Fast flights between flowers. |
| Megalopta sp. | Large | 4:30 a.m. | 6 a.m. | 225 (51-518) | First floral visitor, starts foraging at the beginning of anthesis of male flowers. |
| Bombus sp | Large | 5:00 a.m. | 10 a.m. | 16 (7-30) | Fast flights between flowers. |
| Trigona corvina | Small | 5:30 a.m. | 12 m. | 73 (39-106) | Group recruitment. Predominate at flowers from 6-7 a.m. Aggressive. Enlargement of the pores of the floral nectary. |
| Apis mellifera | Medium-Small | 5:00 a.m. | 12 m. | 27 (9-45) | Non-aggressive behavior. |
Discussion
Bee diversity within and between localities
The data obtained in this study agree with other studies carried out in various locations in the Neotropics (Digital Appendix), showing how different species of Cucurbita are visited by a diverse spectrum of bee species, where the species specialized for the collection of pollen from cucurbits are not always the most frequent (Canto-Aguilar & Parra-Tabla, 2000; Meléndez-Ramírez et al., 2002; Zambrano et al., 2013; Enríquez et al., 2015; Delgado-Carrillo et al., 2018). The groups collected in Costa Rica have been observed in these studies, such as Trigona, Bombus, Augochlora, Ceratina, Eucera (Peponapis), and Africanized bees (Apis mellifera), along with other rarer groups. These inventories have shown a group of dominant and widely distributed species in this crop, accompanied by a series of rare species with more restricted distributions.
Variations between localities in the degree of alteration of the natural habitat and the intensity of agricultural practices have been mentioned as factors that explain changes in the composition of bees species related to squash fields (Krug et al., 2010; Enríquez et al., 2015). However, Meléndez-Ramírez et al. (2002) found a high diversity of bees that visit cucurbit crops in locations with high degradation of their natural flora. To study the factors that determine changes in the composition of bee species that visit squash in Costa Rica would require a more extensive and systematic sampling of localities under variable conditions of habitat alteration. However, certain general trends can be reported. In our study, an important effect of altitude was detected in the composition and species richness of squash fields. A large crop at more than 2 000 m elevation (La Cima de Dota) showed a strong reduction in species richness, while localities at low elevations showed a greater number of species. Hoiss, Krauss, Potts, Roberts and Steffan-Dewenter (2012) have shown how altitude acts as an environmental filter on diversity in bee communities. In Costa Rica, pollination of C. pepo at high elevations seems to be ensured by the abundance of B. ephippiatus. Different species of Bombus have been reported as among the main pollinators of Cucurbita in temperate regions (Shuler et al., 2005, Petersen, Reiners, & Nault, 2013). The effect of changes in altitude can be seen when bees visiting squash plantations are compared between two locations that are very close spatially (Cerro Colón and Piedades, 5 km) but differ by 200 m in altitude. These localities show strong differences in the composition of bee species, particularly, the presence of Eucera (Peponapis) apiculata at only the highest locality. Changes in the characteristics of the life zones are also important, as evidenced by the exclusive presence of the Eucera (Peponapis) crassidentata species in dry forest (Cuajiniquil), an observation previously reported by Wille (1985). Another factor that seems relevant is the size of the crop since in small crops the species richness is lower. However, in these small crops the presence of Eucera (Peponapis) limitaris is constant. This species seems to be ensuring the pollination of small crops of C. pepo, even in deforested regions and under strong agricultural pressure (such as Pacayas and Juan Viñas).
Changes in the composition of pollinators in collections spaced 3 months apart during the rainy season were observed in Piedades. The dominance of social bees of the genus Trigona increased in the second sampling. This variation shows the importance of diversity in the pollination of this crop, where meliponine bees could be ensuring production during times of population decline of other species. The importance of pollinator complementarity during different seasons of a crop has been studied by Delgado-Carrillo et al. (2018) for Cucurbita moschata and by Genung et al. (2017) for other crops.
Bee species rarely reported in Cucurbita crops
This is the first study to show the use of Cucurbita pepo as a resource by bees of the genus Megalopta, bees with the ability to forage during twilight (Wcislo et al., 2004). Megalopta were found at high frequencies in several locations in our study. Some species of this genus show social polymorphism (Wcislo et al., 2004). In this study, two species of Megalopta that show facultative eusociality were found, and it was observed that Cucurbita nectar is used by macro- and microcephalic individuals of both species, which reveals that it is a resource used by potential workers and queens. Another genus not reported previously in C. pepo flowers is Caenaugochlora, a rare bee distributed between Mexico and Northern South America (Michener, McGinley, & Danforth, 1994). Of the various bee inventories in squash plantations in the Neotropics, only Meléndez-Ramírez et al. (2002) have reported the collection of 1 individual of this genus in the Yucatán. However, in this study, it was observed in flowers of C. pepo at different locations; it was among the 3 or 4 most frequent species and practiced early foraging on male flowers. Both Megalopta and Caenaugochlora belong to Halictidae, a family with various species reported in crops of different Cucurbita species, but where this family was represented by other genera, such as Augochlora, Lasioglossum, Augochloropsis and Agapostemon (Canto-Aguilar & Parra-Tabla, 2000; Meléndez-Ramírez et al., 2002; Zambrano et al., 2013; Enríquez et al., 2015; Delgado-Carrillo et al., 2018). These groups were relatively rare or absent in our collections. In a study of the social behavior of Halictidae in Costa Rica, Michener and Kerfoot (1967) observed the presence of Caenaugochlora costaricensis (formerly known as Pseudoaugochloropsis costaricensis) exclusively in Cucurbita flowers. Megalopta bees excavate their nests in dead wood, most frequently in fallen tree branches in the understory (Wcislo et al., 2004). C. costaricensis is a ground-nesting species whose nests have been found in slope banks in roadsides sites (Michener & Kerfoot, 1967). Therefore, these species have different nesting sites. Their presence in squash cultures is favored by small forest fragments, isolated trees and secondary growth nearby farms in the localities where these bee species were found.
Comparison with earlier Costa Rican Eucerini bees surveys
The same species of the subgenus Peponapis reported in this study were observed by Wille in 1985 (Wille, 1985). This stability may be due to the continuous presence of Cucurbita crops in the Central Valley and Guanacaste since 1985 as well as the relative capacity of these species to adapt to rural landscapes, especially small crops, where ground nesting areas are available (Phillips & Gardiner, 2015; Delgado-Carrillo et al., 2017). However, in Guanacaste no individuals of Eucera (Xenoglossa) gaabi, a species reported by Wille (1985), were observed. More collections in Guanacaste are necessary to confirm the possible loss of this species. It is interesting to note that recent inventories of bees in Cucurbita plantations in Mexico have not reported the presence of Xenoglossa subgenus (Meléndez-Ramírez et al., 2002, Delgado-Carrillo et al., 2018, Digital Appendix), even though Hurd and Linsley (1966) showed that Mexico was part of the natural distribution of these species.
Behavioral observations
The various floral visitors observed in this study display differences in foraging strategies. Eucera (Peponapis) and Bombus practice a type of foraging characterized by rapid exploration and extraction of nectar from each flower, allowing them to visit many flowers per unit of time (pers. obs.). This strategy, combined with an early foraging schedule, dense hair and relatively large size, makes these bees very efficient potential pollinators of different species of Cucurbita (Canto-Aguilar & Parra-Tabla, 2000; Serra & Campos, 2010; Delgado-Carrillo et al., 2018). Megalopta and Caenaugochlora, although not large or hairy bees, visit flowers very early, taking advantage of the onset of nectar flow in male flowers, which also makes them potential pollinators. A very different pattern was observed in Trigona, particularly in T. corvina. These bees monopolize the flowers shortly after the start of foraging, practicing a slower and more detailed exploration of flower resources, including the slight laceration of the pores leading to the nectar chamber. The congregation of T. corvina bees near the nectar chamber sometimes repels other larger bees from 7-8 a.m., such as Apis mellifera and Eucera (Peponapis). These characteristics of Trigona foraging on Cucurbita flowers were also noted by Serra and Campos (2010) in Brazil.
In summary, crops of C. pepo in Costa Rica are visited by diverse bees, among which some species may be the main pollinators and others play a secondary role. The importance level of each species in the pollination may be determined by the abundance of each species, their foraging schedule and efficiency in pollen transfer (Herrera 1987; Herrera 1989). This diversity allows the crop to be productive throughout different geographic and seasonal environments (Delgado-Carrillo et al., 2018; Hoehn et al., 2008). It could be suggested that the pollination of this crop is maintained in different geographical regions and different times of the year thanks to the spatial and temporal complementarity of pollination services by different species of bees. More information on the pollinating role of Trigona bees would allow a better understanding of the regulatory factors related to the productivity of this crop. The preservation of nesting sites and diverse floral resources (represented by other crops or wild plant species) are crucial factors to maintain the diverse bee community associated with cultivated Cucurbita, which is very important in the diet and regional culture of the Americas.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.

