











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Las islas oceánicas y los montes submarinos son conocidos como “puntos calientes” o “hotspots” de biodiversidad marina, en especial de especies pelágicas (Worm, Lotze, & Myers, 2003; Morato, Hoyle, Allain, & Nicol, 2010). Dicha formaciones se caracterizan por presentar condiciones oceanográficas y topográficas locales que promueven el transporte y la circulación de nutrientes favoreciendo así la productividad, la biomasa y la diversidad de especies (Worm et al., 2003; Lavelle & Mohn, 2010; Lizano, 2012). Al ser áreas clave para la alimentación y el descanso de especies pelágicas altamente migratorias, estos sitios han sido considerados como corredores biológicos naturales (Klimley & Nelson, 1984; Klimley, 1987; Pitcher et al., 2008; Hearn, Ketchum, Klimley, Espinoza & Peñaherrera, 2010; Heidemeyer, 2015). Dada la importancia biológica de estas formaciones ubicadas en aguas abiertas, el garantizar su protección puede ser una manera efectiva de lograr la conservación de varias especies a la vez (Worm et al., 2003). A lo largo del Pacífico Tropical Oriental (PTO), se han establecido una serie de reservas marinas ubicadas alrededor de las diversas islas, archipiélagos y montes submarinos que conforman el Corredor Marino del Pacífico Tropical Oriental (CMAR) como Malpelo y Gorgona (Colombia), Isla del Coco (Costa Rica), Coiba (Panamá) e islas Galápagos (Ecuador). El objetivo principal de dichas reservas marinas es proteger y manejar los recursos marinos que se encuentran dentro de esta región (Meneses, 2005; Rothschild, 2005).
Desafortunadamente, los montes submarinos han sido históricamente codiciados y sobreexplotados por las pesquerías de todo el mundo (Rogers, 1994). En Costa Rica, tanto la Isla del Coco como los montes submarinos adyacentes han sido blanco de la explotación pesquera por parte de flotas nacionales y extranjeras (Lizano, 2012; Rodríguez & Rosero, 2018). Por lo que, la presión pesquera (legal e ilegal) existente en estos sitios constituye una de las principales amenazas a su integridad ecológica (Arias, Pressey, Jones, Álvarez-Romero, & Cinner, 2016; Rodríguez & Rosero, 2018). Aunque dicha actividad está enfocada principalmente a la captura de atún (Scombridae) y dorado (Coryphaena hippurus), los tiburones de las familias Carcharhinidae, Alopiidae y Sphyrnidae suelen constituir una parte importante de las capturas (Dapp, Arauz, Spotila & O’Connor, 2013; López-Garro, Zanella, Martínez, Golfín-Duarte, & Pérez-Montero, 2016; Rodríguez & Rosero, 2018; Nalesso et al., 2019).
Diversos estudios han documentado la conectividad entre las poblaciones de tiburones y tortugas del CMAR y las zonas costeras de Centroamérica (Bessudo et al., 2011; Ketchum et al., 2014a; Nalesso et al., 2019; Heidemeyer, 2015; MigraMar, 2019). Dicha conectividad resalta la importancia que tienen estas estas islas en la supervivencia de las especies altamente móviles o migratorias. Sin embargo, todavía existe muy poca información sobre el papel que juegan los montes submarinos en la conservación de especies amenazadas de extinción como los tiburones. Este estudio presenta la primera evidencia de asociación de un tiburón zorro (Alopias pelagicus) con el monte submarino Las Gemelas, y la primera evidencia de conectividad espacial de éste monte submarino con la Isla del Coco, ubicada ~50 km al noreste, por parte de un tiburón martillo (Sphyrna lewini).
Materiales y métodos
Área de estudio: El Parque Nacional Isla del Coco (PNIC) se ubica a 550 km al suroeste de la costa pacífica de Costa Rica y comprende el área terrestre de la Isla del Coco (23.85 km2), así como sus aguas circundantes en un radio de 22.2 km (Cajiao, 2008) abarcando una extensión marina de protección absoluta de 1 989 km2. Por su parte, la recientemente creada Área Marina de Manejo Montes Submarinos (AMM-MS) consiste en un rectángulo de 9 649 km2 de extensión ubicado en las aguas circundantes al PNIC, comprendiendo la porción noreste y suroeste de la Cordillera Volcánica del Coco (CVC). De acuerdo con la zonificación del AMM-MS existen dos zonas de protección absoluta, una que consiste de una “ceja” de 742 km2 que colinda con el límite Noreste del PNIC y que actúa como zona de amortiguamiento del PNIC (SINAC, 2013), y otra de 2 733 km2 en el extremo suroeste que se ubica sobre los montes submarinos “Las Gemelas”, cuyas cimas se encuentran entre 150 y 180 m de profundidad respectivamente (Cajiao, 2008; Decreto ejecutivo Nº 36452-MINAET, 2011; Rojas & Alvarado, 2012; SINAC, 2013) (Fig. 1).
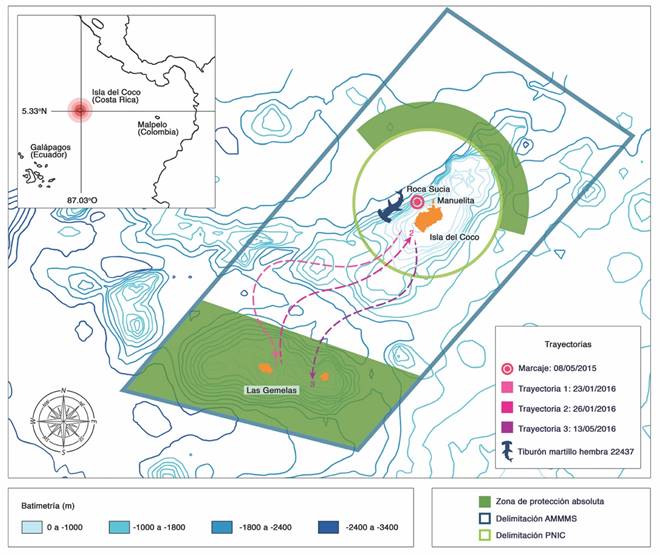
Fig. 1 Fig. 1 Movimientos del tiburón martillo ID 22437 entre el Parque Nacional Isla del Coco (PNIC) y el Área Marina de Manejo Montes Submarinos (AMM-MS) durante mayo del 2015 y mayo del 2016. Las líneas discontinuas sugieren la posible trayectoria realizada por el tiburón. Movements of scalloped hammerhead shark ID 22437 between Cocos Island National Park (CINP) and the Seamounts Marine Management Area during May 2015 and May 2016. Dotted lines suggest the potential shark’s trajectory.
Marcaje y monitoreo: El 8 de mayo del 2015 se colocó un transmisor acústico externo (V16, Vemco Ltd.; ID 22437, 158 dB, transmisión aleatoria entre 40 - 80s; duración de la batería ~3.7 años) en una hembra de S. lewini de aproximadamente 250 cm de longitud total (LT) en el sitio conocido como Roca Sucia, ubicado al noroeste de la Isla del Coco. El transmisor acústico consiste de un cilindro de PVC con un diámetro de 1.6 cm x 7 cm de largo que emite una señal acústica a 69 kHz el cual fue atado a un dardo de acero inoxidable mediante un cable de acero inoxidable de 12 cm de largo. El dardo se fijó al tejido muscular del tiburón ubicado debajo de la primera aleta dorsal mediante un arpón modificado durante la realización de buceo SCUBA, de manera que el transmisor quedó colgando externamente.
Los movimientos del tiburón marcado fueron registrados mediante una red de siete receptores acústicos (VR2W, Vemco Ltd.) instalados en puntos estratégicos alrededor de la Isla del Coco donde se conoce que ocurre la agregación de tiburones martillo adultos, específicamente en los alrededores del islote Manuelita, Ulloa, Alcyone, Roca Sucia, Dos Amigos Pequeño y Punta María (ver Nalesso et al., 2019). Dos receptores acústicos adicionales (VR2AR, Vemco Ltd.) fueron instalados el cinco de mayo del 2015 a 180 y 190 m de profundidad en diferentes cumbres del monte submarino conocido como Las Gemelas, localizado aproximadamente a 50 km al suroeste de la Isla del Coco (Starr, Green, & Sala, 2012). Para garantizar la correcta instalación de los receptores se recurrió al uso de sumergibles tripulados y equipados con brazos mecánicos. Las pruebas para determinar el rango de detección acústica de los receptores en la Isla del Coco estimaron un alcance de entre 150 y 300 m (Nalesso et al., 2019). Los receptores acústicos alrededor de Isla del Coco fueron recuperados y reinstalados cada seis meses por buzos, mientras que los dos receptores en Las Gemelas fueron recuperados al cabo de 15 meses mediante una señal acústica que activó el sistema de liberación. Una vez liberados los receptores fueron transportados hacia la superficie por medio de boyas. Desafortunadamente, uno de los receptores acústicos no pudo ser recuperado, probablemente debido a alguna falla en el sistema de flotación. Los datos acústicos de todos los receptores fueron descargados a una base de datos electrónica administrada por MigraMar.
Durante la expedición a Las Gemelas, se realizó una jornada de pesca de investigación entre las 15:00 y las 21:00 hrs, utilizando un palangre corto modificado (300 m de línea madre, 20 anzuelos circulares #14 sin barba), que resultó en la captura de un tiburón zorro pelágico macho (Alopias pelagicus) de 236 cm de LT, el cual fue equipado con un transmisor acústico interno (V16, Vemco Ltd.; ID 32359; 158 dB; transmisión aleatoria entre 120 - 230s; duración de la batería ~3.2 años). En este caso, se procedió a realizar una pequeña incisión quirúrgica en el vientre del tiburón para insertar el transmisor en la cavidad peritoneal. Todos los métodos utilizados durante esta investigación fueron aprobados y realizados bajo los permisos de investigación ACMIC-I-2015-008 y 2016-I-ACMIC-07 otorgados por el Área de Conservación Marina Cocos (SINAC-MINAE).
Análisis de datos: Los datos de presencia-ausencia del tiburón martillo alrededor de los receptores fueron analizados de manera descriptiva. El tiburón se consideró presente en el área de estudio cuando dos o más detecciones fueron registradas durante el mismo día por cualquiera de los receptores. Los días y sitios de detección fueron graficados en una línea de tiempo para determinar los movimientos, sitios visitados y las posibles trayectorias realizadas. Además, se calculó el índice de residencia en el área de estudio (IR), dividiendo el número de días en que el tiburón fue detectado por cualquiera de los receptores entre el número total de días de monitoreo (Espinoza, Heupel, Tobin, & Simpfendorfer, 2015). Debido a que la red de receptores no cubría toda el área del PNIC y el AMM-MS, el índice de residencia es un valor mínimo aproximado del tiempo que permaneció el tiburón dentro del área de estudio, ya que el tiburón pudo estar presente en la Isla del Coco o en los montes submarinos sin ser detectado por los receptores acústicos (Nalesso et al., 2019).
Resultados
Desde la fecha de su marcaje el 8 de mayo del 2015 el tiburón martillo ID 22437 fue detectado 139 veces de manera intermitente por los receptores de la Isla del Coco y Las Gemelas hasta mayo del 2016, las cuales nunca ocurrieron por más de tres días consecutivos. Los datos acústicos muestran que el tiburón martillo permaneció en los alrededores de la Isla del Coco durante siete días después de su marcaje en Roca Sucia. Durante este tiempo el tiburón visitó repetidamente Roca Sucia y realizó un desplazamiento a Manuelita, para regresar un día después (Fig. 1 y Fig. 2). Después de ocho meses de ausencia, se registró la presencia del tiburón martillo por primera vez en Las Gemelas el 23 enero del 2016. Tres días más tarde el tiburón regresó a la Isla del Coco, donde fue detectado nuevamente por el receptor de Roca Sucia. El tiburón fue detectado por última vez 28 días más tarde en Las Gemelas el 13 mayo 2016 (Fig. 1 y Fig. 2). El índice de residencia de S. lewini en el área de estudio fue de 0.02. Roca Sucia fue el sitio más visitado por el tiburón desde su marcaje, con un total de 114 detecciones, en comparación con 6 en Manuelita y 19 en Las Gemelas. Todas las detecciones ocurrieron entre las 7:00 y las 12:00 horas. Adicionalmente, se registró la presencia un tiburón zorro pelágico en Las Gemelas el 18 de marzo del 2016, casi un año después de su marcaje (5 mayo del 2015). A. pelagicus permaneció durante cinco días más en este monte submarino, siendo detectado un total de 30 veces.
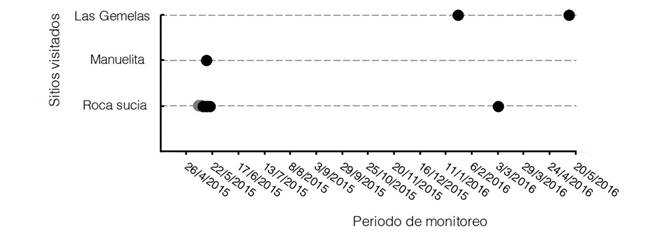
Fig. 2 Fig. 2 Registro de presencia-ausencia del tiburón martillo ID 22437 en los sitios ubicados dentro del PNIC y el AMM-MS, entre mayo 2015 y mayo 2016. El punto gris indica la fecha de marcaje del tiburón. Presence-absence of scalloped hammerhead shark ID 22437 at sites located at Cocos Island National Park (CINP) and Seamounts Marine Management Area, during May 2015 and May 2016. The gray dot indicates tagging date.
Discusión
El presente estudio constituye la primera evidencia de asociación entre un tiburón zorro con el monte submarino Las Gemelas, y el primer reporte de conectividad espacial entre éste y el PNIC por parte de un tiburón martillo. Se ha propuesto que los tiburones y otras especies marinas son capaces de utilizar diversas pistas ambientales, químicas y/o magnéticas para orientarse y navegar a través del océano (Lohmann, Lohmann, & Endres, 2008; Nosal, Chao, Farrara, Chai, & Hastings, 2016). Es posible que el tiburón martillo se haya desplazado a través de la Cordillera Volcánica del Coco utilizando las anomalías magnéticas de estos sitios para orientarse durante su viaje (Klimley, 1993).
Todas las detecciones acústicas del tiburón martillo ID 22437 ocurrieron únicamente durante la mañana, lo cual es congruente con estudios que han demostrado que S. lewini es una especie altamente gregaria que tiende a formar grupos alrededor de islas y montes submarinos en aguas relativamente someras (25-30 m) principalmente durante el día (Klimley & Nelson, 1984). Se ha propuesto que la formación de grandes grupos de S. lewini alrededor de estos sitios puede ser una estrategia para optimizar su energía, antes de emprender el viaje hacia aguas profundas durante la noche probablemente para alimentarse (Klimley & Nelson, 1984; Holland, Wetherbee, Peterson, & Lowe, 1993; Bessudo et al., 2011; Ketchum et al., 2014b).
La baja residencia y los largos periodos de ausencia exhibidos por el tiburón martillo ID 22437 muestran que este tiburón utilizó de manera temporal la Isla del Coco y el monte submarino Las Gemelas. Los periodos de ausencia (e. i. días, semanas o meses) han sido observados en otras islas y montes submarinos del PTO, lo cual hace pensar que se trata de un comportamiento común en la especie (Klimley & Nelson, 1984; Bessudo et al., 2011; Nalesso, 2014; Nalesso et al., 2019; Ketchum et al., 2014a). Basados en datos provenientes de 84 tiburones martillo monitoreados entre 2005 - 2013, Nalesso et al. (2019) encontraron que los tiburones martillo de la Isla del Coco no permanecen de manera constante en la isla, sino que estos suelen ausentarse por periodos de nueve meses o más. Aunque la ausencia de detecciones durante varios meses a la vez podría estar relacionada con el desplazamiento del tiburón a lugares de la isla donde no había receptores instalados, es muy probable que el tiburón martillo ID 22437 haya realizado viajes a lugares alejados, como aguas continentales u otras islas del CMAR. Diversas investigaciones han planteado que S. lewini podría estar utilizando las islas o montes submarinos como puntos de referencia durante sus migraciones diarias o estacionales (Klimley & Nelson, 1984; Hearn, et al., 2010; Ketchum et al., 2014a; Hoyos-Padilla, Ketchum, Klimley & Galván-Magaña, 2014). Los movimientos de ida y vuelta realizados por S. lewini entre las islas Malpelo, Cocos y Galápagos ya han sido debidamente documentados en varios estudios (Bessudo et al., 2011; Ketchum et al., 2014a; Nalesso et al. 2019). Estos movimientos entre islas sugieren que las poblaciones de Galápagos y Malpelo pueden estar utilizando la Isla del Coco, y potencialmente Las Gemelas como puntos de referencia durante sus migraciones estacionales a zonas costeras de Centroamérica y Sudamérica (Nalesso et al. 2019). Recientemente se ha sugerido que las especies altamente migratorias utilizan la Cordillera Volcánica del Coco como una “vía” o autopista para trasladarse entre los sitios de agregación y alimentación ubicados en aguas abiertas del PTO (Peñaherrera-Palma et al. 2018).
La residencia temporal de los tiburones martillo en el PNIC podría estar relacionada con sitios de alimentación, de refugio, estaciones de limpieza, o con eventos reproductivos (Hamilton & Watt, 1970; Clarke, 1971; Klimley & Nelson, 1984; Dewar et al., 2008; Bessudo et al., 2011; Oliver, Hussey, Turner, & Beckett, 2011; Torres-Rojas et al., 2013; Ketchum et al., 2014a). Hasta la fecha solo ha sido reportado un evento de apareamiento en la Isla del Coco (Salinas de León, Hoyos-Padilla, & Pochet, 2017). Investigaciones realizadas por Sibaja-Cordero (2008) y Nalesso (2014) han señalado que los tiburones martillo en la Isla del Coco suelen permanecer o regresar continuamente a sitios como Alcyone, Roca Sucia y Manuelita donde existen estaciones de limpieza permanentes. Por lo tanto, los movimientos entre Manuelita y Roca Sucia, pueden estar impulsados probablemente por la disponibilidad de espacio en las estaciones de limpieza ubicadas en estos sitios.
A pesar de los esfuerzos de investigación realizados en los últimos años, aún existe muy poca información sobre la fauna que habita en los montes submarinos y sobre su relación con otros montes submarinos, islas oceánicas y zonas costeras (Rogers, 1994; Cortés & Blum, 2008; Cortés et al., 2012). Aunque los movimientos de un solo individuo pueden no ser una representación fiel del comportamiento de toda la población, este hallazgo recalca la importancia de seguir investigando las posibles interconexiones ecológicas de los montes submarinos con la Isla del Coco y su relación con otras islas del PTO (Cortés & Blum, 2008). Si bien la conectividad entre las poblaciones de S. lewini de las islas Malpelo, Cocos y Galápagos ha sido demostrada en varios estudios (Bessudo et al., 2011; Ketchum et al., 2014a; Nalesso et al., 2019), hasta el momento no se había obtenido evidencia acerca de la conectividad entre las poblaciones de estas islas oceánicas y los montes submarinos adyacentes.
Adicionalmente, la presencia periódica de especies amenazadas de extinción como A. pelagicus y S. lewini en Las Gemelas confirma la importancia de estos montes submarinos para las especies altamente móviles que convergen en estos hábitats pelágicos (Wehrtmann & Cortés, 2009; Morato et al., 2010). A. pelagicus es una especie mayormente oceánica que habita en aguas profundas tropicales y subtropicales del Pacífico e Indo-Pacífico y de la cual se conoce muy poco sobre su biología y ecología (Reardon, Márquez, Trejo, & Clarke, 2009). Al ser sitios conocidos por su alta productividad y disponibilidad de presas, es probable que S. lewini y A. pelagicus hayan coincidido en el monte submarino Las Gemelas para alimentarse. Sin embargo, dado el poco conocimiento que se tiene sobre la dinámica ecológica de estos sitios, sería indispensable recabar mayor información que nos ayude a fundamentar o rechazar esta hipótesis.
Tanto A. pelagicus como S. lewini son importantes componentes de la pesca artesanal y comercial, lo cual ha contribuido de manera importante al declive de sus poblacionales a escala mundial (Baum et al. 2009; Zanella, López-Garro, & Arauz, 2009; Reardon et al., 2009; Hearn, Utreras, & Henderson, 2010; White, Myers, Flemming, & Baum, 2015). Entre las estrategias más recomendadas para lograr la conservación de las especies marinas están aquellas que logran minimizar las interacciones con pesquerías (Nalesso et al., 2019). En este sentido, la creación de AMPs con políticas de protección estricta alrededor de las islas y montes submarinos se ha propuesto como una estrategia para conservar y proteger la biodiversidad marina en aguas abiertas (Worm et al., 2003). Sin embargo, la efectividad de dichas AMPs sigue siendo cuestionada, principalmente por la falta de información científica acerca de las especies que se desea proteger, así como por la carencia de recursos económicos disponibles para su manejo y protección (Hearn et al. 2010; White et al., 2015; Arias et al., 2016).
Aunque el AMM-MS posee zonas de protección absoluta donde se prohíben las actividades pesqueras, estas aún son permitidas en el resto del área marina (Fig. 1). Durante sus recorridos entre el PNIC y Las Gemelas, el tiburón martillo se desplazó más de 30 km a través de aguas donde se permite la pesca, incrementando así el riesgo de ser capturado por las flotas pesqueras que faenan en esta área. Lo anterior, resalta la necesidad de contar con mejor información sobre la distribución y dinámica espacial de especies altamente móviles que nos permita evaluar y proponer esquemas de zonificación que resulten efectivos y acordes a los objetivos de conservación para los que fueron creados el PNIC y el AMM-MS (Hearn et al., 2010; Bessudo et al., 2011).
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

