











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Los tomoptéridos son poliquetos holoplantónicos con una vasta distribución en los oceános y mares de nuestro planeta, desde las regiones polares hasta las tropico-ecuatoriales, en las capas superficiales, las intermedias y las abisales hasta más de 3000 m de profundidad, así como en las zonas neríticas y principalmente en las oceánicas (Day, 1967; Day,1975; Ushakov, 1972; Fernández-Álamo, 2006; Fernández-Álamo & Thuesen, 1999). Sus relaciones filogenéticas con otros poliquetos son discutibles e inciertas y la mayoría de los autores los consideran como un grupo aberrante, demasiado especializado para poder llevar una existencia pelágica, aunque en general se relacionan con los filodócidos (Støp-Bowitz, 1948; Tebble, 1960, Tebble, 1962; Dales & Peter, 1972; Ushakov, 1972; Orensanz & Ramírez, 1973).
Estos poliquetos han sido descritos como depredadores de otros tomoptéridos y de varios grupos de organismos del zooplancton como quetognatos y larvas de peces (McIntosh, 1924; Lebour, 1923; Åkesson, 1962; Pettibone, 1963; Rakuza-Susczewski, 1968). Aunque su presencia en el plancton es escasa, en algunas regiones particulares, su abundancia es considerablemente mayor, por lo que Southern (1911) y Pettibone (1963) asumieron que debían jugar un papel importante en el alimento de algunos peces. Por ejemplo, Åkesson (1962) registró que Tomopteris helgolandica, en las aguas de Suecia, puede llegar a ser tan abundante que satura las redes de arrastre y la autora tuvo la oportunidad de observar en el Southamton Oceanography Center en Inglaterra, muestras de zooplancton de regiones abisales conteniendo gran abundancia de tomoptéridos. Otro aspecto interesante de estos organismos del holoplancton es su utilización en aspectos zoogeográficos, ya que se han encontrado relaciones significativas entre la distribución de algunas especies y los movimientos de las masas de agua (Ekman, 1953; Dales, 1955; Tebble, 1960, 1962).
En el Pacífico oriental los registros de las especies de la familia Tompteridae son aislados y escasos (Rosa,1908; Chamberlin, 1919; Dales, 1957; Berkeley & Berkeley, 1960; Fernández-Álamo, 1983, Fernández-Álamo,1987, Fernández-Álamo,2000; Rozbaczylo, Antezana & Pinochet,1987). Algunas especies recolectadas frente a las costas de América Central han sido documentadas por Treadwell (1928),Treadwell (1941), Berkeley & Berkeley (1961), Berkeley & Berkeley (1960), Berkeley & Berkeley (1964); Vicencio-Aguilar & Fernández-Álamo (1996); Jiménez-Cueto, Suárez-Morales & Morales-Ramírez (2012).
Este trabajo tiene como finalidad describir la estructura y la diversidad de especies de tomoptéridos recolectados en la región de surgencias del Domo Térmico de Costa Rica en el Pacífico centroamericano, así como revisar los antecedentes de estos poliquetos en el área de estudio y sus afinidades zoogeográficas.
Materiales y métodos
El Domo Térmico de Costa Rica es una interesante región situada en el Pacífico centroamericano, importante desde varias perspectivas, tanto científicas como productivas. Se trata de un fenómeno océano-atmosférico en el que se combinan la dinámica particular de las corrientes marinas y el patrón de vientos, originando un afloramiento de aguas frías, ricas en sales disueltas, principalmente nitratos y fosfatos necesarias para el desarrollo de una abundante producción de fitoplancton y zooplancton, básicos para acrecentar y mantener una compleja y próspera comunidad biológica que incluye una alta diversidad de peces, reptiles, aves y mamíferos, lo que la convierte en un espacio crítico para conservar y estudiar la biodiversidad marina del Pacífico oriental en particular y para el planeta en general (Cromwell, 1958; Wyrtki, 1966, Wyrtki, 1981; Fiedler, 2002; Kessler, 2006; Fiedler & Talley, 2006). Por ello la región del Domo Térmico de Costa Rica ha sido declarada como Zona de Importancia Ecológica y Biológica, lo que propicia la conservación, la protección y el estudio de esta notable región (Jiménez, 2016).
El Laboratorio de Oceanografía Física del Instituto de Ciencias del Mar y Limnología de la Universidad Nacional Autónoma de México a cargo del Dr. José Barberán organizó entre 1979 y 1982 cuatro campañas oceanográficas, llamadas DOMOS, para estudiar las características físicas, químicas y biológicas de la región del Pacífico centroaméricano. Los datos oceanográficos de estos estudios se encuentran en Barberán, Gallegos, & Padilla (1984) y Barberán, Gallegos, Fernández & Mee (unpub. data). Cortés (2009) mencionó varios de los trabajos realizados con algunas de las muestras de zooplancton recolectadas en estas expediciones.
El material zooplanctónico fue recolectado en 33 de las localidades realizadas durante el crucero oceanográfico “Domo IV” a bordo del B/O “Puma” del 26 de julio al 12 de agosto de 1982. La cobertura del área abarcó entre los ~2° y 12°11’ N y los 84°30’a 92° 57’ W (Tabla 1). Los muestreos de zooplancton se obtuvieron mediante arrastres oblicuos de 200 m de profundidad a la superficie, con redes estándar de 1.0 m de diámetro en la boca, 3.5 m de manga, 0.33 mm de malla con medidores de flujo TSK-946. De cada una de las muestras, con ayuda de un microscopio estereoscópico, fueron separados los organismos e identificados a nivel de especie utilizando los trabajos de Støp-Bowitz (1948), Tebble (1962) y Ushakov (1972). Con el material separado se elaboró una colección de referencia que se encuentra depositada en el Laboratorio de Invertebrados de la Facultad de Ciencias de la UNAM, en la Ciudad de México.
TABLA 1 TABLE 1 Ubicación de las localidades muestreadas y datos de temperatura (°C) y salinidad (ups) a diferentes profundidades durante la campaña oceanográfica “Domo IV” al Domo Térmico de Costa Rica, durante julio-agosto de 1982 Location of sampled localities and temperature data (°C) and salinity (ups) at different depths during the oceanographic campaign “Domo IV”, Costa Rica Thermal Dome, July-August 1982
| Estación | Latitud | Longitud | Temperatura | Salinidad | ||||
| Superficial | 50m | 75m | Superficial | 50m | 75m | |||
| 2 | 10°00’ | 90°30’ | 27.8 | 15.3 | 13.7 | 33.5 | 34.7 | 34.7 |
| 9 | 10°03’ | 87°04’ | 28.3 | 19.3 | 15.8 | 32.3 | 34.5 | 34.6 |
| 12 | 9°30’ | 86°01’ | 28.4 | 20.6 | 17.3 | 32.4 | 34.5 | 39.7 |
| 21 | 9°00’ | 84°30’ | 28.8 | 23.7 | 18.2 | 31.7 | 33.9 | 34.7 |
| 29 | 9°00’ | 87°30’ | 28.1 | 16.5 | 14.9 | 33.5 | 34.6 | 34.8 |
| 32 | 9°00’ | 89°00’ | 27.6 | 15.6 | 14.3 | 33.5 | 34.7 | 34.8 |
| 35 | 9°01’ | 90°31’ | 28.2 | 19.1 | 15.2 | 33.0 | 34.6 | 34.6 |
| 38 | 9°00’ | 92°00’ | 28.0 | 18.9 | 14.2 | 33.3 | 34.6 | 34.7 |
| 42 | 8°00’ | 91°30’ | 28.1 | 17.7 | 14.0 | 33.0 | 34.6 | 34.7 |
| 45 | 8°00’ | 90°00’ | 28.2 | 19.4 | 14.3 | 33.2 | 34.6 | 34.7 |
| 48 | 8°00’ | 88°30’ | 28.3 | 17.3 | 15.0 | 33.2 | 34.6 | 34.7 |
| 55 | 6°30’ | 87°00’ | 28.3 | 18.0 | 15.7 | 33.0 | 34.7 | 34.8 |
| 58 | 5°00’ | 87°00’ | 27.9 | 20.9 | 17.6 | 33.0 | 34.7 | 34.8 |
| 61 | 5°00’ | 88°29’ | 27.5 | 21.3 | 16.5 | 33.0 | 34.6 | 34.6 |
| 66 | 5°00’ | 89°29’ | 27.5 | 23.1 | 15.0 | 32.9 | 34.5 | 34.7 |
| 70 | 4°00’ | 89°59’ | 27.1 | 26.7 | 16.0 | 33.2 | 33.8 | 34.6 |
| 74 | 2°30’ | 90°00’ | 26.5 | 22.6 | 15.8 | 34.0 | 34.4 | 34.9 |
| 77 | 3°30’ | 88°30’ | 26.5 | 22.9 | 18.7 | 33.2 | 34.4 | 34.8 |
| 82 | 6°26’ | 88°30’ | 27.8 | 17.9 | 15.3 | 32.9 | 34.6 | 34.8 |
| 89 | 10°00’ | 88°30’ | 27.9 | 16.8 | 14.8 | 33.5 | 34.7 | 34.7 |
| 92 | 11°29’ | 88°30’ | 28.5 | 27.6 | 20.7 | 32.7 | 33.3 | 34.4 |
| 96 | 11°59’ | 89°00’ | 28.7 | 28.4 | 25.3 | 32.6 | 32.9 | 33.9 |
| 98 | 11°12’ | 90°07’ | 28.6 | 26.9 | 20.6 | 32.7 | 33.7 | 34.4 |
Para jerarquizar la dominancia de las especies determinadas se aplicó la prueba de Olmstead y Tukey (Sokal & Rohlf, 1998). Las especies se caracterizaron en dominantes, constantes, ocasionales y raras a partir de la relación entre las densidades de los organismos y sus frecuencias de aparición. El criterio de discriminación se basa en la media de la frecuencia de aparición relativa para el eje de las “X” y de la media respectiva del logaritmo natural (LN) de la sumatoria de la densidad absoluta (D) para el eje de las “Y”, de tal forma que al trazar ambas se perfilan cuatro cuadrantes. Las especies dominantes, son aquellas que presentan la densidad y la frecuencia mayor a ambas medias. Las constantes aparecen con densidades menores a la media, pero la frecuencia es mayor. Las ocasionales son la que tienen densidad mayor a la media y frecuencia menor, mientras que las raras muestran densidades y frecuencias menores a ambas medias.
Resultados
La Tabla 1 presenta la temperatura y la salinidad observadas en la superficie, 50 y 75 m mostrando las variaciones de estos factores en la región superior donde fueron tomadas las muestras de zooplancton. En el primer caso se pudieron detectar las localidades donde está ocurriendo el afloramiento de aguas más frías y en el segundo, en general, como la capa superficial menos salina que forma una burbuja (Barberan et al., 1984).
Se separaron y determinaron más de 1000 ejemplares de poliquetos holoplanctónicos pertenecientes a la familia Tomopteridae, de los cuales 859 fueron determinados a nivel de especie, presentando el siguiente arreglo sistemático:
Phylum Annelida Lamarck, 1801
Clase Polychaeta Grube, 1851
Orden Phyllodocida Dales, 1962
Familia TOMOPTERIDAE Grube, 1850
Género Tomopteris Eschscholtz, 1825
Localidad tipo: Pacífico (mares del sur)
Tomopteris elegans Chun, 1887. Localidad tipo: Islas Canarias
Tomopteris euchaeta Chun, 1887. Localidad tipo: Mar Mediterráneo
Tomopteris nationalis Apstein, 1900. Localidad tipo: Nápoles
Tomopteris planktonis Apstein, 1900. Localidad tipo: Atlántico (regiones central y norte)
La abundancia y la distribución de las especies de Tomopteris determinadas en la región de estudio se puede observar en la Fig. 1. Tomopteris elegans fue el poliqueto mejor representado en el área de estudio, con la mayor abundancia relativa (62 %) y la mayor frecuencia de aparición (83%) observándose en 19 localidades y en algunas de ellas, con un elevado número de organismos, por ejemplo, la estación 58 en donde hubo 177 (Tabla 2). De acuerdo a la relación entre su abundancia y su porcentaje de aparición se caracterizó claramente como la especie dominante (Fig. 2, Tabla 2). Tomopteris planktonis ocupó el segundo lugar en abundancia con 22% del total y una frecuencia de aparición elevada de 78 %, por lo que también se caracterizó como especie dominante. Tomopteris nationalis apareció, de acuerdo a los valores de su abundancia y a su porcentaje de aparición, como una especie constante. Comparativamente, los valores más bajos de abundancia y porcentaje de aparición, caracterizaron a T. euchaeta como especie rara.
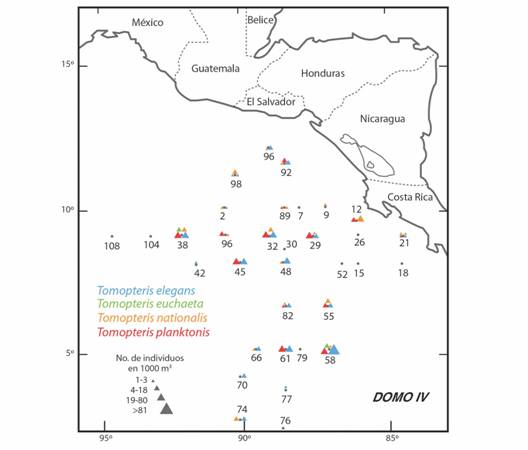
Fig. 1 Fig. 1 Abundancia y distribución de las especies de Tomopteris (Tomopteridae) registradas durante la campaña oceanográfica “Domo IV” en julio-agosto de 1982 en el Domo Térmico de Costa Rica. Abundance and distribution of the species of Tomopteris (Tomopteridae) recorded in the Costa Rica Thermal Dome during the “Domo IV” oceanographic campaign in July-August 1982.
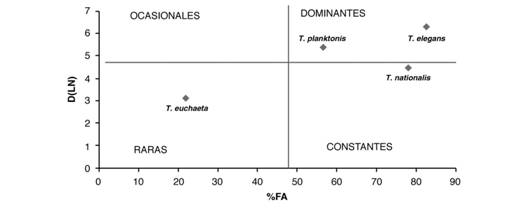
Fig. 2 Fig. 2 Análisis de dominancia específica de los tomoptéridos del Domo Térmico de Costa Rica durante la campaña oceanográfica “Domo IV” entre julio-agosto de 1982. D (LN)= Logaritmo natural de la densidad; %FA= Frecuencia de aparición. Analysis of specific dominance of the tomopterids in the Costa Rica Thermal Dome during the oceanographic expedition “Domo IV” between July-August, 1982. D (LN)= Natural logarithm of density; % FA = Frequency of appearance.
TABLA 2 TABLE 2 Abundancia relativa (%A) y frecuencia de aparición (%FA) de tomoptéridos (Polychaeta) durante la campaña oceanográfica “Domo IV” al Domo Térmico de Costa Rica, julio-agosto de 1982 Relative abundance (%A) and frequency of occurrence (%FA) of tomopterids (Polychaeta) during the oceanographic campaign “Domo IV”, Costa Rica Thermal Dome, July-August 1982
| Estación | T. elegans | T. euchaeta | T. nationalis | T. planktonics |
| 2 | 1 | 0 | 1 | 0 |
| 9 | 0 | 0 | 1 | 0 |
| 12 | 0 | 0 | 21 | 13 |
| 21 | 2 | 1 | 1 | 7 |
| 29 | 14 | 0 | 1 | 25 |
| 32 | 63 | 0 | 12 | 36 |
| 35 | 0 | 0 | 2 | 7 |
| 38 | 43 | 6 | 5 | 39 |
| 42 | 2 | 0 | 0 | 0 |
| 45 | 22 | 8 | 0 | 22 |
| 48 | 74 | 0 | 3 | 0 |
| 55 | 7 | 0 | 5 | 5 |
| 58 | 177 | 6 | 3 | 28 |
| 61 | 72 | 0 | 0 | 22 |
| 66 | 7 | 0 | 2 | 0 |
| 70 | 13 | 0 | 0 | 0 |
| 74 | 5 | 0 | 5 | 0 |
| 77 | 3 | 0 | 0 | 0 |
| 82 | 4 | 0 | 1 | 3 |
| 89 | 0 | 0 | 3 | 2 |
| 92 | 7 | 0 | 3 | 7 |
| 96 | 15 | 0 | 3 | 0 |
| 98 | 1 | 1 | 17 | 0 |
| A% | 62 | 2.6 | 10.4 | 25.2 |
| FA% | 83 | 22 | 78 | 57 |
Discusión
Rouse & Pleijel (2003) mencionan que los tomoptéridos, en general, son relacionados con los filodocidos, aunque actualmente existen pocas evidencias disponibles para ello, ya que las homologías morfológicas con otros poliquetos están oscurecidas por su apariencia altamente divergente, tanto en las formas adultas, como en sus primeros estados ontogenéticos descritos por Åkesson(1962). Por esas razones sugirieron encontrar evidencias moleculares y una ampliación de los organismos estudiados, que puedan garantizar sus relaciones filogenéticas, incluyendo la posibilidad que se definan fuera de los filodócidos e incluso fuera del clado Acicula. También se debe señalar que la sistemática del grupo es compleja y requiere de una profunda revisión de los tipos en las colecciones de varios museos (Fernández-Álamo, 2018) para establecer de forma inequívoca la validez de las especies.
Treadwell (1928) hizo el primer registro de tomoptéridos para el Pacífico frente a Centro América con la descripción de dos nuevas especies, Tomopteris opaca y T. tentaculata, cuyos ejemplares fueron recolectados a los 4º 50´N y 87º W, al sur de Isla de Coco, Costa Rica; las descripciones son cortas e incompletas y Hartman (1956) consideró que se trata, para la primera especie, de dos organismos maltratados que podrían corresponder a T. nisseni. Para la segunda opina que se trató de juveniles de alguna otra especie. Sin embargo, estimó que por la presencia de los primeros setígeros y los órganos en rosetas en los troncos parapodiales de las dos primeras patas, pueden pertenecer a T. nationalis de la que Jiménez-Cueto et al. (2012) registraron un ejemplar en una localidad llamada Wafer al sur de Isla de Coco, Costa Rica. En este estudio T. nationalis fue caracterizada como especie constante (Fig. 2). Day (1967) asumió que T. apsteini, posiblemente sería sinónima con T. nationalis y Fernández-Álamo (2000) apoyó esta idea. La afinidad zoogeográfica de T. apsteini, de acuerdo con Tebble (1962), es de aguas tropicales y subtropicales y se encuentra distribuida en el Pacífico norte.
Las especies dominantes fueron T. elegans y T. planktonis. La primera también tiene afinidad por las aguas tropicales y subtropicales, pero su límite en el Pacífico norte se extiende hasta la zona de transición con las aguas subpolares, mientras que la segunda ha sido definida como una especie cosmopolita, que en general, se distribuye en todas las masas de agua, desde la superficie hasta 750 m de profundidad (Tebble, 1960, Tebble, 1962), aunque Ushakov (1972) no la encontró en la región noroccidental del Pacífico.
En relación a T. elegans Chun, 1887, Tebble (1962), Day (1967) y Dales & Peter (1972) la definen con status valido y diferente de T. pacifica Izuka, 1914. Sin embargo, en el portal World Register of Marine Species (http://www.marinespecies.org/index.php) se considera no válida y como sinónima T. pacifica, definiendo a Chun (1887) y a Berkeley (1924) como autores de la descripción original; lo que lleva a confusión si se toma en cuenta que Dales & Peter (1972) en su Synopsis of the Pelagic Polychaeta determinan que la especie de Berkeley, 1924 síse considera sinónima deT. pacifica Izuka, 1914, pero no con la descrita por Chun en 1887. En este contexto, y por experiencia de la autora, se tomó el criterio de Dales & Peter (1972) y se aconseja hacer una revisión del material tipo para definir con certidumbre esta situación.
Tomopteris euchaeta es considerada por la mayoría de los especialistas como representante de un género diferente, sin embargo Day (1967) argumentó que Rosa (1908) basó su nuevo género Enapteris en observaciones de juveniles y es apoyado por las observaciones de Fernández-Álamo (1983), Fernández-Álamo (2000) quien señaló que hay un error en la descripción de las pínulas parapodiales, por presentar dos zonas, una externa densa y una interna hialina que sí rodea los troncos parapodiales (principal criterio para la conformación del género Enapteris) y por lo tanto pertenece a Tomopteris. Sobre esta base en el presente estudio se considera válida a Tomopteris euchaeta que se registró como una especie rara, pero que Fernández-Álamo (1983), Fernández-Álamo (2000) la registró bien distribuida desde Cabo Corrientes, México (20º N) hasta Callao, Perú (20º S) y desde las costas de América hasta los 125º W en el Pacífico Tropical Oriental.
Es importante señalar que el patrón de distribución y abundancia de las especies de esta familia de poliquetos en el área de estudio confirma su afinidad por las regiones oceánicas y de batimetria profunda como ha sido señalado por Day (1975). La característica más sobresaliente de estos gusanos es que en las especies dominantes T. elegans y T. planktonis se observó un notorio incremento en su abundancia (Tabla 1, Tabla 2), relacionado con la región donde la termoclina fue más somera según lo registrado por Barberán et al (1984) en la misma campaña oceanográfica.
Tebble (1960), Tebble (1962) estableció que la producción total de plancton es mayor en las altas latitudes que en los trópicos, sin embargo, la costa occidental de África es la excepción, debido a los efectos de la corriente fría de Benguela por lo que el número total de organismos del plancton es 10 veces mayor, en los primeros 50 m de profundidad, que en las zonas del Antártico, la subtropical y la tropical del Atlántico sur y en el Pacífico norte. Este fenómeno coincide con el alto registro de la abundancia de los tomoptéridos en la región del Domo Térmico, en donde el número de ejemplares es considerablemente mayor (Tabla 2) que lo registrado por Tebble (1960), Tebble (1962) y en otras regiones diferentes como el Atlántico norte (Støp-Bowitz, 1948) y el Pacífico Tropical Oriental (Fernández-Álamo, 1983). Como se mencionó, hay ciertas regiones donde existe una elevada abundancia de estos poliquetos, como las aguas cercanas a Suecia (Åkesson, 1962) y las muestras de aguas profundas que fundamentan las suposiciones de Southern (1911) y Pettibone (1963) de que deben ser importantes como alimento de varios peces y que coincide con la riqueza en la biodiversidad y la productividad observada en el Domo Térmico de Costa Rica (Wehrtmann &Cortés, 2009).
También es importante considerar si los resultados obtenidos en el periodo de estudio pueden estar influenciados por el efecto de El Niño 1982-1983 (Romero-Centeno et al., 2003; Yuan & Yamagata, 2014), por lo que sería relevante poder comparar los resultados aquí obtenidos, con los de otras muestras recolectadas durante los periodos fríos de La Niña y las de periodos intermedios entre ambos fenómenos.
Declaración de ética: la autora declara que está de acuerdo con esta publicación; que no existe conflicto de interés de ningún tipo; y que ha cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

