











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
Isla del Coco National Park (ICNP) is an oceanic island located in the Eastern Pacific and has belonged to Costa Rica since 1869. It was declared a National Park in 1978 (Bolívar, Rovinsky, & Ching, 2000), and is considered a natural laboratory for the study of biodiversity (Madriz, 2009).
The Isla del Coco´s forests are evergreen, thanks to the high humidity. They are dominated by the endemic tree Sacoglottis holdridgei (Cuatec, 1964) (Humiriaceae); that reaches 25 meters in height and a meter in diameter. (Porras-Jiménez, 2012, Trusty, Kesler, & Delgado, 2006). The crowns are the support for epiphytes and nesting for marine birds including the migratory species Gygis alba (White Tern). The understory is clear, mostly dominated by bushes and the grass Hypolytrum amplum (Poepp. & Kunth in Kunth, Enum., 1837) (Cyperaceae, Trusty et al., 2006), and regeneration of canopy species are scarce (Acosta-Vargas, 2016). The forest has a low diversity compared with the Costa Rica continental forests. Plant diversity reached 296 species of vascular plants, and ferns are the most diverse group (Estrada-Chavarría, Sánchez-González, & Rodríguez-González, 2020). Thirty-eight species are endemic, including the trees S. holdridgei and Cecropia pittieri (B.L. Rob. 1912) (Urticaceae), and the tree fern Cyathea alfonsiana (L. D. Gómez, 1971) (Cyatheaceae, Trusty et al., 2006, Estrada-Chavarría et al., 2020).
The study of the flora of the ICNP, began when the naturalist George W. Barclay, collected plant specimens in the year 1838 (Zamora, 2008). Recent studies of Porras-Jiménez (2012) started the documentation of tree mortality in the forest of Isla del Coco, leading to changes in the floristic composition and vegetation structure. This study aroused interest in additional research focused on population status and structural changes in the forests (Acosta-Vargas, 2016). In general, the investigation had been focused on flora descriptions (Rivera, 2006; Trusty et al., 2006; Zamora, 2008). Topics related to forest dynamics, forest structure, growth rates, and ecological processes are poorly known in Isla del Coco.
The forest’s dynamics are linked to mortality and recruitment (Lewis et al., 2004). Finegan (1992) mentions that disturbance plays a significant role, since the death of individuals causes the formation of clearings, activating natural regeneration, and thus keeping the diversity and structure of the forest. Alterations in the natural regeneration process, conditions the permanence of the forest structure and plants’ composition. Those effects are stronger in insular ecosystems, where some ecological processes are absent (Madriz, 2009). The problems of natural regeneration become bigger and more complex with the introduction of large herbivores onto the islands. Herbivores impact the development of the forest due to their branch grazing, changing growth patterns and altering the abundance and survival of plant regeneration (Dirzo, 2001). Herbivores can also affect the availability of tree seeds and their dispersion, which is a critical process for forest maintenance (Janzen, 1970). In addition, they can cause alterations to the soil of the forests via compaction (Sierra, 2001). The introduction of the feral pigs to ICNP was recorded (Sus scofra) in 1793, and white-tailed deer (Odocoileus virginianus) in 1935 (Montoya, 2016).
Forest growth is assessed by monitoring the size increase of trees over time and reported in terms of diameter, basal area, or volume increase (Louman, Quiros, & Nilsson, 2001). Growth rate can be affected by factors such as site conditions (light, heat, humidity), soil (nutrients, pH), competition between individuals, and the species performance (Castillo-Ugalde, 1991; Valerio, Esquivel, & Salas, 1998; Gadow, Sanchez, & Alvarez, 2007; Quesada-Monge, Castillo-Ugalde, Lobo-Segura, & Barrantes, 2010; Bowman, Brienen, Gloor, Phillips, & Prior, 2013). This investigation was carried out to describe the forest population status and its species, for determining threats and actions to restore and preserve the forests of Isla del Coco.
Material and methods
Site: Isla del Coco National Park (ICNP) is in the Eastern Pacific in the coordinates 05°30’57” N & 87°05’40” W; at an approximate distance of 500 km from Puntarenas, Costa Rica. It has a terrestrial area of 24 km2 and a marine protected area of 2011 km2 (SINAC, 2016). It is of volcanic origin, and the only emerging point of the Cocos Ridge (Castillo et al., 1988). Isla del Coco receives an annual rainfall from 5 000 to 7 000 mm. The average annual temperature of 25.5 °C, the minimum is of 23.1 °C and the maximum is of 27.6 °C (Herrera, 1986; Gutiérrez-Fonseca, Ramirez, Umaña & Springer, 2013; Fig. 1). There are two forest types are in Isla del Coco: the Premontane Pluvial Rainforest (PPR) located under 300 m.a.s.l (meters above sea level). with an area of 18.51 km2 and the Cloud Forest (CF) from 300 to 634 m.a.s.l. covering an area of 4.68 km2 (Porras-Jiménez, 2012).
Experimental design: The baseline for this study consists of data obtained from 15 Permanent Sampling Plots (PSP) of 50 x 50 m (0.25 ha), subdivided into subplots of 10 x 10 m and established in the altitudinal gradient. The first nine PSP were placed in the PPR in 2006 (Rivera, 2006), and measured again in 2012 and 2017 giving six and eleven years of observation. The second group of six PSP were placed in the CF in 2012 (Porras-Jiménez, 2012), and measured again in 2017 giving five years of observation (Fig. 1). All individuals with a diameter at breast height (DBH) ≥ 5 cm were measured in diameter and height.
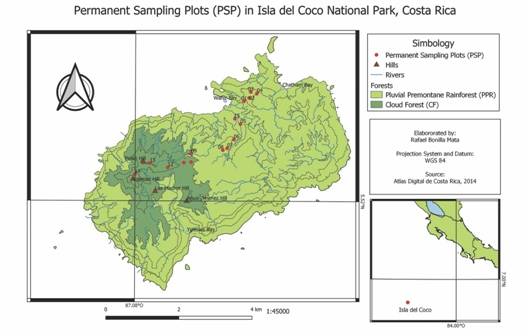
Fig. 1 Permanent sample plots established in the Premontane Pluvial Rainforest (PPR) and the Cloud Forest (CF). Isla del Coco National Park, Costa Rica, 2017.
Isla del Coco has a low tree diversity, and the forest structure depends on two species in both forests. Sacoglottis holdridgei, which is a dominant species accounting around 60 % of the canopy trees, and the canopy codominant Ocotea insularis (Meisn. Mez, 1889) (Lauraceae). In the middle canopy in the PPR the most abundant species are Henriettea succosa (Aubl. DC. 1828 and Henriettella odorata Markgr. 1941), (Melastomataceae). In the CF, the middle canopy is dominated by the tree fern Cyathea alfonsiana (Cyatheaceae) that is accompanied by Miconia dodecandra Cogn. 1887 (Melastomataceae), Myrsine pellucidopunctata Oerst. 1862 (Primulaceae). The species cited before were included for their higher abundance and importance in the future of the forest structure.
Mortality: The annual mortality rate was obtained using the Nebel et al., (2001) model:
Rm = (ln(No)-ln(No-Nm))/∆t,
Rm: Mortality rate, No: Number of individuals in the first measurement, Nm: Number of dead individuals in the last measurement, ∆T: Time differential (5, 6, 11 years).
Recruitment: The annual recruitment rate used the model of Phillips et al., (1994)
Rr = (ln(No-Nm-Nr)-ln(No-Nm))/∆t,
Rr: Recruitment rate, No: Number of individuals in the first measurement; Nm: Number of dead individuals in the last measurement, Nr: Number of trees recruited in the last measurement, ∆T: Time differential (5, 6, 11 years).
Regeneration: In each PSP, two transects of 1 x 50 m were established for regeneration sampling, corresponding to an intensity of 4 % of the PSP area. The regeneration of the same species surveyed in the PSP was identified at the species level, abundance recorded, and classified into seedlings of 0.3-1.5 m of height, and low saplings from 1.5 height to 4.9 cm of diameter (Louman et al., 2001).
Growth rate: We calculated the average growth rate (AGR) for both the diameter and the basal area by diametric category using the baseline data obtained by Rivera (2006) and Porras (2012). The growth analysis was conducted on forests and tree species. Two periods of growth were used for the Premontane Pluvial Rainforest: 2006-2017 (PPR 2006-2017) and 2012-2017 (PPR 2012-2017), for the Cloud Forest the period used was 2012-2017) (CF 2006-2017). The average growth rate was calculated as follows
AGR = (x1-x2)/t,
x1: diameter or final basal area, x2: diameter or initial basal area and t: Time. (5, 6, 11 years).
Data analysis: Mortality, recruitment, regeneration, and growth analyses were carried out for forests and species, and the values obtained extrapolated to hectare (ha). We reported the results using the confidence intervals (α = 0.05), the T-test was applied for independent samples to compare between forests and species. The results were compared with the studies conducted by Rivera (2006) and Porras-Jiménez (2012).
Results
The statistical test showed differences between forests and species in growth patterns, mortality, recruitment, and regeneration.
Global patterns in forests population: In terms of mortality, the PPR reported mortality of 61 ind./ha. (±42; SD = 55; N = 9) and an annual mortality rate of 5.75 %. The CF mortality rate was 339 ind./ha. (±96; SD = 91; N = 6), representing a yearly mortality rate of 6.31 %, in the period 2012-2017. Mortality between forests types is significant (Media test: p < 0.0001; N = 15), being higher in the Cloud Forest.
The recruitment for the period 2012-2017 in the PPR was 104 ind./ha. (±48; SD = 62; N = 9), for an annual rate of 5.38 %, and in the CF was 205 ind./ha. (±84; SD = 76; N = 6) for an annual rate of 5.90 %. Recruitment is statically different (p < 0.0001; N = 15).
The regeneration found in the PPR was 4 867 seedlings/ha (±1772; N = 9) and 789 low saplings/ha (±325; N = 9). The CF reported 8 217 seedlings/ha (±4936; N = 6), and 483 low saplings/ha (±437; N = 6). The global account of regeneration reported a significant difference between forests (p < 0.05; n = 29).
Population structure in both forests showed a J inverted pattern characteristic of an uneven-aged forest, and both forests reported individuals in all diametric classes (Fig. 2). An important characteristic of the CF population is the high number of individuals in the classes: seedlings 8 217 ind./ha. (±4 936; N = 6), 59 % more than in the PPR, and 5-10 cm 1 167 ind./ha. (±352; N = 6), 643.7% more than the PPR. This huge difference between forests was because of the high abundance of C. alfonsiana in the lower strata which represents 91.9 %, or 1 072 ind./ha. (±417; N = 6) in the class 5-10 cm, given as result a dense understory.
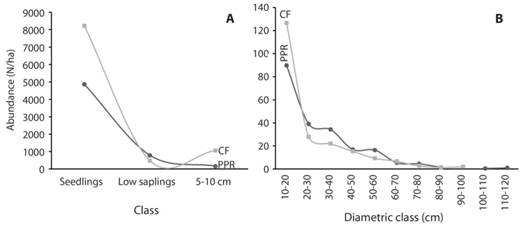
Fig. 2 Diametric distribution of the abundance (N/ha) for regeneration and individuals less than 10 cm (< 10 cm) (A) and for individuals > 10 cm (B), Premontane Pluvial Rainforest and the Cloud Forest, Isla de Coco National Park, Costa Rica, 2017.
Species analysis in the Premontane Pluvial Rainforest: A more significant variation was found between the results of previous years (Fig. 3). Sacoglottis holdridgei dominates the canopy, in 2012 it represented 45 % of all species recruited, but in 2017 it represented 23 %, reflecting a reduction in recruitment of 15 %. Mortality reached 20 ind./ha. (±18; SD = 23; N = 9), representing 30 % of the total mortality of the species found (Fig. 3E and Fig. 3F). The regeneration reported 56 ind./ha. (±41; SD = 53, N = 9, Fig. 3G) in the seedlings class, low sapling was not reported. The diametric distribution by species showed that S. holdridgei supported the forest structure over 15 cm in diameter, confirming its dominance in this forest (Fig. 4).
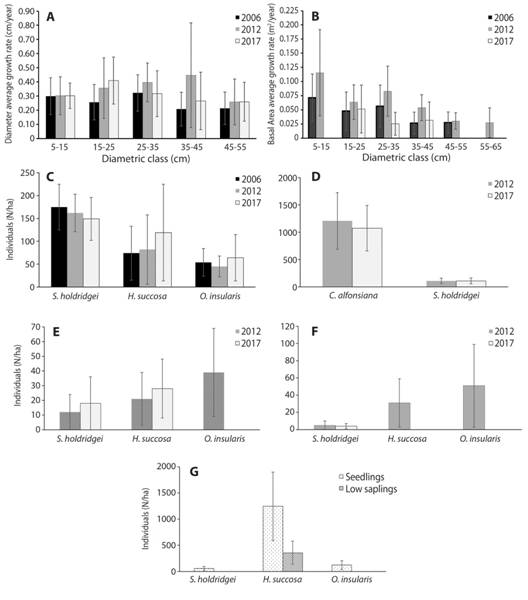
Fig. 3 Dynamics and growth of the forests: A) average growth rate in diameter (cm/year) per year, B) average growth rate in basal area (m2/year), C) abundance of each species (N/ha) in the Premontane Pluvial Rainforest (PPR), D) abundance (N/ha) in the Cloud Forest (CF), and E) mortality (N/ha), F) recruitment (N/ha), and G) regeneration (N/ha) in the PPR, Isla del Coco National Park, Costa Rica, 2017.
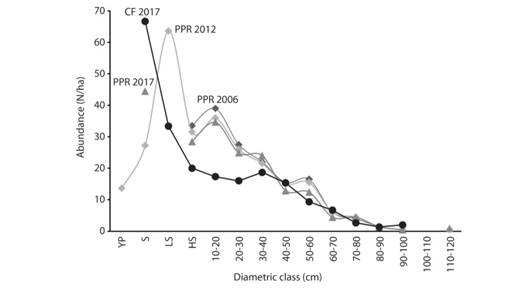
Fig. 4 Population structure of Sacoglottis holdridgei in three different times: 2006 (Rivera, 2006), 2012 (Acosta-Vargas, 2016), and 2017 (present study), for the Premontane Pluvial Rainforest (PPR) and the Cloud Forest (CF), Isla del Coco National Park, Costa Rica.
Ocotea insularis trees produce many shoots under DBH. In the period 2012-2017 they only report recruitment and mortality on their shoots, but no new individuals. Additionally, O. insularis reported 122 seedlings/ha (±84; N = 9), but low sapling was not reported.
Melastomataceae species increased their abundance in the last years, mainly H. succosa, which passed from 19 % in 2006 to 31 % in 2017. On the other hand, mortality was 23 ind./ha. (±20; SD = 26, N = 9). Furthermore, H. succosa had the highest regeneration, accumulating 28 % of the individuals found. It is reported in greater quantities in clearings caused by the fall of death or large trees, where H. succosa was the most abundant with 1 600 ind./ha. (±1 230; SD = 886, N = 9; Fig. 3G).
In terms of regeneration, H. succosa is the most abundant species. The species complex of the Melastomataceae family (include H. succosa, H. odorata, Miconia dodecandra) supports J-inverse forest structure in the low classes of seedlings and low-saplings.
Species analysis in the Cloud Forest: In this forest, the species dynamics were more stable in the years of study (Fig. 3D). The species with the highest mortality was the tree fern C. alfonsiana with 272 ind./ha. (±121; SD = 115, N = 6), representing 80 % of the mortality for this forest, in turn, it had the highest recruitment with 155 ind./ha. (±89; SD = 84, N = 6). Its regeneration reported 2667 seedlings/ha (±2 151; SD = 2050, N = 6), and did not report low saplings. The upper canopy species did not present recruitment or mortality in the CF.
Forests growth in diameter: The forests reported different increases in diameter being those statistically different in the same period 2012- 2017 (p = 0.0414; n = 15, Fig. 3A). In the Premontane Pluvial Rainforest, the increase in the period 2006-2017 was 0.29 cm/year (± 0.08; SD = 0.09; N = 523), and during period 2012- 2017 it was 0.34 cm/year (± 0.09, SD = 0.07; N = 807); the growth rates reported were statistically different between periods (p = 0.0044; N = 5).
On the other hand, the cloud forest showed annual diametric growth for the period 2012-2017 of 0.33 cm/year (±0.07; SD = 0.07; N = 366). The largest increases reported were in the categories 15-25 cm and 25-35 cm for both forests (Table 1).
TABLE 1 Diametric and basimetric increment of the periods of study for the Premontane Pluvial Rainforest (PPR) and Cloud Forest (CF), Isla del Coco National Park, Costa Rica
| Diametric increase (cm year-1) | ||||||||||||
| PPR 2006-2017 | PPR 2012-2017 | CF 2012-2017 | ||||||||||
| X | CL | SD | N | X | CL | SD | N | X | CL | SD | N | |
| Forest | 0.29 | 0.08 | 0.09 | 523 | 0.34 | 0.09 | 0.07 | 807 | 0.33 | 0.07 | 0.07 | 366 |
| Specie | - | - | - | - | - | - | - | - | - | - | - | - |
| O. insularis | 0.50 | 0.30 | 0.36 | 159 | 0.57 | 0.30 | 0.39 | 74 | 0.27 | 0.23 | 0.22 | 87 |
| H. succosa | 0.19 | 0.09 | 0.11 | 70 | 0.26 | 0.1 | 0.13 | 182 | - | - | - | - |
| H. fascicularis | - | - | - | - | - | - | - | - | - | - | - | - |
| S. holdridgei | 0.25 | 0.05 | 0.06 | 278 | 0.31 | 0.08 | 0.1 | 312 | 0.32 | 0.12 | 0.29 | 156 |
| Basimetric increase (m² ha-1 year-1) | ||||||||||||
| PPR 2006-2017 | PPR 2012-2017 | CF 2012-2017 | ||||||||||
| X | CL | SD | N | X | CL | SD | N | X | CL | SD | N | |
| Forest | 0.306 | 0.101 | 0.121 | 523 | 0.433 | 0.129 | 0.167 | 807 | 0.260 | 0.092 | 0.088 | 366 |
| Specie | - | - | - | - | - | - | - | - | - | - | - | - |
| O. insularis | 0.106 | 0.064 | 0.061 | 74 | 0.145 | 0.126 | 0.164 | 74 | - | - | - | - |
| H. succosa | 0.015 | 0.013 | 0.016 | 70 | 0.040 | 0.038 | 0.050 | 182 | - | - | - | - |
| H. fascicularis | - | - | - | - | - | - | - | - | - | - | - | - |
| S. holdridgei | 0.187 | 0.053 | 0.063 | 278 | 0.232 | 0.05 | 0.07 | 312 | 0.171 | 0.060 | 0.057 | 156 |
X: Average; CL: Confidence limits (95 %); SD: Standart Desviation; N: Sample.
Forests growth in basal area: The PPR reported in 2017 a basal area of 21.01 m2/ha (±5.47 SD = 5.22; N = 9); compared with the previous surveys it represents an increase of 0.13 m²/ha in the period 2006-2017 and a decrease of -0.20 m²/ha (0.92 %) in the period 2012-2017. The Cloud Forest registered a basal area of 20.95 m²/ha (±3.54; SD = 3.37; N = 6); this value decreased -0.58 m²/ha (2.7 %) compared to the 2012 baseline. Increases between forests are not statistically different for the period 2012-2017 (p = 0.2778; N = 15; Fig. 3B; Table 1).
Growth of canopy species: Ocotea insularis presented the largest diametric increase in BPP in the period 2006-2017 with 0.50 cm/year (±0.30; SD = 0.36; N = 156), and in the period 2012- 2017 0.57 cm/year (±0.41; SD = 0.39, N = 144); in the Cloud Forest the increase was 0.27 cm/year (±0.23; SD = 0.22; N = 56). For the 2012-2017 period for both forests, the reported growth is statistically different (p < 0.0001; N = 15).
Sacoglottis holdridgei in PPR (2012-2017) reported an increase of 0.31 cm/year (±0.08; SD = 0.10; N = 312) and in the period 2006-2017 0.25 cm/year (±0.05; SD = 0.06; N = 278). The Cloud Forest reported an increase of 0.32cm/year (±0.12; SD = 0.10; N = 156). In the same period the diametric growth between the PPR and CF was not statistically different (p = 0.8772; N = 15). Sacoglottis holdridgei is the only species that presents individuals in all diametric categories.
Henriettea succosa: presented an increase of 0.19 cm/year (±0.11; SD = 0.13; N = 70) for BPP 2006-2017 and 0.26 cm/year (±0.10; SD = 0.13; N = 182) for BPP 2012-2017. This species is absent in Cloud Forest.
Basal area growth by species: In the Premontane Pluvial Rainforest S. holdridgei reported in 2017 14.84 m2/ha (±5.78; SD = 5.5, N=9) a lower value than in 2012, corresponding to a reduction in basal area of 0.58 m2/ha (3.8 %); Similarly, O. insularis reported a loss of 0.11 %. All Melastomataceae species surveyed reported an increase of 0.27 m2/ha (1.3 % of total basal area 2017); especially, H. succosa which represented 68.8 % of the increased basal area, and 32.1 % of the basal area loss by S. holdridgei (Fig. 5).
For the Cloud Forest in 2017, S. holdridgei registered 13.25 m2/ha (±4.32; SD = 4.12, n = 6), and only 0.9 % of basal area loss. The species with the biggest impact was C. alfonsiana with a loss of 3.75 %, which in 2017 accumulated 4.20 m2/ha (±2.69; SD = 2.56, N = 6). Ocotea insularis was the species that reported the greatest increase with 0.32 m2/ha corresponding to a 1.5 % recovery of the basal area and followed by Melastomataceae with an increase of 0.34 % (Fig. 5).
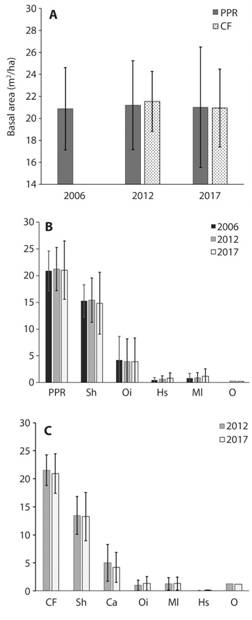
Fig. 5 A) Basal area comparison per type of forest (PPR: Premontane Pluvial Rainforest and CF: Cloud Forest) in three different times of measurement: Rivera (2006), Porras-Jiménez (2012) and present study (2017). B) Basal area comparison between species in the PPR and C) in the CF (Sh: Sacoglottis holdridgei, Ca: Cyathea alfonsiana, Oi: Ocotea insularis, Hs: Henriettea succosa, Ml: other Melastomataceae species, and O: other species) in three different times of measurement (2006, 2012, 2017). Isla del Coco National Park, Costa Rica.
Discussion
Due to the increasing anthropogenic degradation of forests, investigations in forest dynamics and growth for conservation and restoration are necessary. For Isla del Coco National Park forests, these investigations are unknown, but are highly required, from degradation caused by because of introduced herbivores and climate change.
Added to the isolation, the small size of Isla del Coco combined with a little altitudinal gradient (0-620 m.a.s.l), we found significant statistical differences in recruitment, mortality, regeneration and growth rates between both forests. Even though, the PPR and CF share the same canopy species, dynamic and ecological process, along with abiotic factors and geological origin (Castillo et al., 1988; Bergoeing, 2012) made them different as a response for adaptation. That condition only reaffirmed that this is an ecosystem of great diversity (Trusty et al., 2006) and complexity.
Patterns in species and their influence in forests: Here, we presented the results for the periods 2006-2017 and 2012-2017 which showed different patterns. The mortality in the ICNP´s forests reached higher values than in continental mature tropical forests that range from 0.9 to 2.91 % (Marín, Nygard, Rivas, & Oden, 2005). Also, it was even higher than the 2.99 % reports by Porras-Jiménez (2012) during the period 2006-2012.
This condition of mortality for this window of 12 years of observation in the PPR showed that values around 3 % are characteristic of Isla del Coco. Moreover, we can associate this with a new period of forest dynamics, reflected in the high mortality of 65 % at a canopy level by individual replacement, and a resources liberation. Sacoglottis holdridgei accumulated all the mortality, and in contrast, H. succosa took advantage and recruiting 38 % of individuals, and 71 % of the total regeneration.
This pattern was observed in 2013 (Acosta-Vargas, 2016), where species of Melastomataceae family represented 45 % of the regeneration. Here, we see a sustained increase in Melastomataceae dominance.
In the period 2006-2017 the PPR grew in diameter at a rate of 0.29 cm/year, a higher rate than the one reported by Porras-Jiménez (2012) of 0.20 cm/year in the period 2006-2012. In the time frame 2012-2017, the rate increased to 0.34cm/year, as a result of higher growth rates in the species S. holdridgei, O. insularis, and H. succosa. As a direct result, the growth in the basal area increased for the same period reaching 0.433 m2/ha/year. This acceleration in growth is the result of the release of resources by mortality, mainly large trees of S. holdridgei. The remaining stand grows and recovers the basal area within the values that the site can support. This type of condition has been reported in forests of La Cangreja National Park (Acosta-Vargas, 2012) and the Osa Peninsula, Costa Rica, (Quesada-Monge, Acosta-Vargas, Garro-Chavarría, & Castillo-Ugalde, 2012).
The species presented a diametric and basal area increase according to their ecological guild. The Melastomataceae species recorded are durable heliophytes; all are fast-growing, relatively long-lived and need light for their establishment; meanwhile, O. insularis and S. holdridgei are shade tolerant. Sciophytes have less growth than heliophytes, but their growth increases if a canopy opening happens because those species require a percentage of light to move from intermediate diametric categories to maturity, (Porras-Jiménez, Acosta-Vargas, Quesada-Monge, & Castillo-Ugalde, 2014; Finegan, 1993; Lamprecht, 1990). It was seen that durable heliophyte species showed the highest increase in both forests, similar to performance was reported in a natural forest in Venezuela (Ramírez, Acevedo, Ataroff & Torres 2009).
The Cloud Forest showed a more stable dynamic at canopy level in contrast with PPR, whose the recruitment rate is under the mortality rate. In terms of individuals, they are nearly balanced with a net loss of 0.37 % in the PPR, and 0.41 % in the CF, a pattern reported in continental forests (Lieberman, Lieberman, Peralta, & Hartshorn, 1985).
Furthermore, this stability is usual for a forest in the homeostatic stage. The most critical changes in the CF were at the low canopy with a high rate of mortality of C. alfonsiana, which reached 80 % but also reported high recruitment and regeneration. In particular, C. alfonsiana is a key species defining the CF structure.
These patterns of mortality, recruitment, and regeneration presented two conditions. First, the forest structure fits the J-inverted model, typical of uneven-age forest (Lamprecht, 1990). This population structure supposes a guarantee of permanence in the time because of the stock of young trees that will reach the canopy.
The second is the abundance of canopy species, from which S. holdridgei is the species close to fit the J-inverted model above a DBH > 5 cm. Although, its regeneration is low, and by 2017 it replicates the trend reported in 2013 (Acosta-Vargas, 2016). Above DBH > 20 cm S. holdridgei define the forest population structure; in this sense, it is a mono-specific forest, and S. holdridgei shapes the heights and the structure of the canopy. Ocotea insular is a companion species, and its population structure is incomplete in both forests.
Overall, Sacoglottis holdridgei is the most important species in the forest dynamics of the ICNP because it is an endemic species and presents a continuous horizontal distribution (Porras-Jiménez et al., 2014). Comparing the current state of the population of S. holdridge with the reported by Acosta-Vargas (2016), some changes happened in four years, when the population structure looked improving the regeneration for 2017, at least for seedlings (0.3-1.5m), but young plants (0-0.3m) and low saplings did not report individuals (Fig. 4). Dissimilarities found are related to factors such as spatial distribution, because the regeneration counted was in 50 % of the same sites surveyed in 2013, 33 % did not register regeneration both in 2013 and 2017; this means that regeneration has not taken place in some areas of Isla del Coco because of filters that avoid its establishment such as: seed predation by micromoth larvae (Blastobasidae) that affects 70 % of the fruits (Retana-Corrales, 2018), pigs’ effects by soil removal (Sierra, 2001), and rats that gnawed 68 % of the fruits (Gómez, 2004); also a depleted seed bank, herbivory and absence of disperser (Acosta-Vargas, Rovere, Camacho- Sandoval 2020).
Another important aspect is that since 2006, the population of S. holdridgei has been decreasing (Rivera, 2006, Fig. 4), evidencing the high mortality rate and limited recruitment in the PPR that starts from the lowest diametric class. In the case of the CF, population was stable. Comparing the CF with the PPR, the Cloud Forest presents an older population because the abundance in the diametric classes between 10-40 cm is an average 35 % lower than in the PPR. It talks to the topic of forest limitations and a global problem of tree replacement in the medium and long term (Fig. 4).
This species (S. holdridgei) also had the highest mortality in all the diametric categories and reported low values of recruitment and regeneration, unlike other species. In comparison with the data obtained by Acosta-Vargas (2016) of 27±20 seedlings/ha, S. holdridgei reported an increase in the number of low saplings/ha with 56 (±41; SD = 53, N = 9; Fig. 1d), but there are no significant differences (CL α = 0.05); however, it did not report any individuals in the low sapling category.
Forecasting forest changes and their impacts on diversity: What is happening nowadays in the Isla del Coco´s forests is a transformation of its structure and composition. A process that took place decades ago with the introduction of herbivores and potentially increased affect with climate change. Changes are stronger in the Premontane Pluvial Rainforest (PPR), where the continuous stress caused by white-tailed deer and pigs is still going on. This change brings with it the loss of dominance of S. holdridgei, and this induces a shift in forest vertical and horizontal structure.
The change that the forest faces leads to the substitution of S. holdridgei with Melastomataceae species, and especially H. succosa, all them middle canopy species. This species replacement will create a dramatic shift in the future forest with a tremendous impact in microclimate conditions, and ecological processes such as birds nesting, epiphytic support, and seed banks. The forest structure is modified, losing canopy height and even a less long-lived forest species. Finally, the epiphyte biodiversity of bromeliads, orchids, and ferns will be affected by the loss of the substratum provided by the old Sacoglottistrees.
Everything described above was recorded in PSP 5 in the period 2012-2017. This PSP had the highest mortality of trees at the canopy, 58 % corresponding to Sacoglottis, and the highest recruitment and regeneration of individuals of Melastomataceae family, mainly H. succosa represented 70 % of individuals’ recruitment and regeneration.
Henriettea succosa is a durable heliophyte species. As a pioneer species it occupies clearings for its establishment and needs light for its development (Lamprecht, 1990). So, the space realized by the death of trees in the canopy is occupied by H. succosa in the absence of fast-growing species belonging to the upper strata, and regeneration of the dominant species S. holdridgei and O. insularis.
In the long term, the greenery of Isla del Coco will be keep, but it will be supported by another kind of forest in structure and species composition. Here, we emphasize the herbivores caused damage, as ecosystem engineers who are altering the native forest and species population. White-tailed deer and pigs’ impact on the island is enormous; their introduction created an incomplete trophic chain, characterized by the absence of predators that can control their population. Around 300 years of pigs’ presence and 80 years of white-tailed deer presence is not enough time for ecosystem and species to adapt, to evolve and to deal with the new members.
But not everything is wrong, PSP 5 presented the largest diametric increase with 0.60 cm/year (±0.09; SD = 0.39; N = 78) in the period 2012-2017. Consequently, all the species surveyed in this plot speeded their growth. Henriettea succosa showed a diametric increase of 0.45 cm/year (±0.08; SD = 0.26; N = 38); while O. insularis and S. holdridgei, both shade tolerant, reported 0.94 cm/year (±0.27; SD = 0.53; N = 17) and 0.54 cm/year (±0.14; SD = 0.29; N = 20) respectively.
This increase is even higher than the previous period from 2006 to 2012. This dynamic pattern was explained by Finegan (1993): shade-tolerant species trend to increase their diameter if openings in the canopy take place, which is the pattern reported for forests in the Central Pacific of Costa Rica (Acosta-Vargas 2012), and the Osa Peninsula (Quesada et al., 2012). This condition will result in trees with larger diameters, which balance the basal area supported by the site.
The need to act: Isla del Coco National Park is a unique site, due to its endemism, climate, and native fauna and flora. Joined with its history, beauty, and breathtaking landscapes helped it to be become declared a World Heritage and Humanity and Wetland RAMSAR site. No matter those prestigious designations for preservation, threats still affect the forest future.
The tree canopy species have not been regenerated and recruited at the same rate that equates with their mortality. In the future, the forest will present a change in the current structure and composition, mainly in the PPR rather than in the CF.
The CF may be more sensitive to the El Niño phenomena, due to changes in the cloudiness pattern and horizontal rain. Furthermore, tree canopy death will bring drier conditions affecting the rich fern community at the understory level.
As a way of stopping degradation for restoring the forest, control of herbivores is mandatory and urgent, just if the idea is to preserve the native flora and fauna of Isla del Coco.
We recommend active forest restoration, increase PSP net, climate monitoring, and more investigation in ecology to fill up information gaps. In this way, we can better understand and help the ecosystem, especially the Cloud Forest.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.

