











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Los atributos de la comunidad bentónica han sido con frecuencia utilizados para estimar la calidad del hábitat y el nivel de impacto ambiental en varias regiones del mundo (Çinar et al., 2015; McLaverty et al., 2020). Para el estudio del zoobentos se han establecido cuatro grupos fundamentales a partir del tamaño de los organismos: microbentos, meiobentos, macrobentos y megabentos (Sakshaug, Helge-Johnsen, & Kovacs, 2009). Aunque existen múltiples criterios acerca de los límites de tamaño definidos para cada categoría, la repercusión ecológica de cada grupo es única y está objetivamente determinada. Respecto a los organismos mayores de 4 milímetros, clasificados como megazoobentos (Alcolado, 1990; Alcolado et al., 1998; Sakshaug et al., 2009), se ha demostrado que tienen una relevante función como bioindicadores de contaminación ambiental, impactos antrópicos y biodiversidad (Alcolado, & Espinosa, 1996).
También se ha hecho notar la importancia del megazoobentos como componente de la trama trófica y alimento esencial para varias especies comerciales entre las que destacan las langostas (Colinas-Sánchez, & Briones-Fourzán, 1990; Espinosa et al., 1990). A nivel mundial, y particularmente en Cuba, los moluscos bentónicos han sido reportados como componentes esenciales en la dieta de varias langostas espinosas entre las que se encuentra Panulirus argus (Latreille, 1084), principal recurso pesquero de Cuba y uno de los productos más apreciados en el mercado internacional (Herrera et al., 1991; Puga-Millán et al., 2013; Puga-Millán et al., 2018).
La mayor parte de los estudios de bentos en Cuba implican sólo componentes de la macro o meiofauna (Arias-Scheirber et al., 2008; Armenteros, Williams, Creagh, & Capetillo-Piñar, 2008; Hidalgo, Toledo, & Granados-Barba, 2015). Respecto al megazoobentos de fondos blandos un menor número de estudios se ha desarrollado, por lo que el tema requiere de estudios que aporten información acerca del empleo de sus propiedades para, como han sugerido Labra, Hernández-Miranda y Quiñones (2020), inferir cambios en las condiciones del hábitat y el flujo de energía trófica en los ecosistemas. El golfo de Batabanó, región pesquera más importante de Cuba (Cruz et al., 1990; Baisre, 2018), ha sido de las pocas en la que se han desarrollado investigaciones que incluyen al megazoobentos, siendo el trabajo de Alcolado (1990) la principal obra de referencia.
El interés pesquero en el golfo de Batabanó ha motivado el estudio de procesos de elevada repercusión ecológica, siendo numerosas las investigaciones que describen significativos cambios en la flora y fauna bentónica (Arias-Scheiber et al., 2008; Cerdeira-Estrada, Lorenzo-Sánchez, Areces-Mallea, & Martínez-Bayón, 2008). El escenario actual, que evidencia el decrecimiento de las capturas de múltiples recursos entre los que destaca P. argus, le confiere aún mayor relevancia a este tipo de estudios (Baisre, 2018; Puga-Millán et al., 2018). En este contexto, fue objetivo de la presente investigación, determinar variaciones espacio-temporales en la densidad y biomasa del megazoobentos de los fondos blandos del golfo de Batabanó, e inferir, haciendo énfasis en los moluscos, el efecto de tales variaciones en la disponibilidad de alimento para las langostas.
Materiales y métodos
El presente estudio se realizó en los fondos blandos del golfo de Batabanó, en el suroeste del Cuba, en el Mar Caribe, considerado una extensa macrolaguna con 6 m de profundidad media y predominio de fondos blandos cubiertos de pastos marinos (Thalassia testudinum Banks ex König) de densidad entre media y baja (Alcolado, 1990). Se registraron datos del 2007, 2008, 2009, 2010, 2015, 2017 y 2018, tratados como “período 2007-2018”, en 12 de los 15 sitios muestreados por Alcolado (1990) en 1983. Además, se utilizaron los datos de 1983 como referencia para realizar comparaciones de densidad y biomasa de los organismos con los obtenidos en el presente estudio (Alcolado, 1990) (Fig. 1).
Fig. 1 Fig. 1 Ubicación de los sitios de muestreo para colecta de megazoobentos (excluyendo esponjas, corales y gorgonáceos) en los fondos blandos del golfo de Batabanó, Cuba. Las estaciones representadas pertenecen al período 2007-2018. Location of the sampling sites for the collection of megazoobenthos (excluding sponges, corals and gorgonians) in the soft bottoms of the Gulf of Batabanó, Cuba. The stations represented belong to the 2007-2018 period.
Los muestreos se realizaron mediante arrastres (tres réplicas por sitio). La metodología seguida tanto para el muestreo en 1983 como para el presente estudio se especifica en Alcolado (1990). La rastra utilizada en el período 2007-2018, que posee en su entrada un área de 56 x 25 cm2, no fue la misma empleada en 1983, pero ambas presentan similitudes estructurales y funcionales que hacen comparables los resultados siempre que la abundancia se estandarice como densidad de organismos (ind/m2) y la biomasa se exprese en unidades de área (g/m2). En 2007-2018 cada arrastre (réplica) tuvo una duración promedio de 40 s, en los que el barco se movió a 2 nudos, para un área total arrastrada de 21 m2. De 2007 a 2015 se utilizaron sólo como valor anual de densidad y biomasa debido a que no se cuenta con los datos respectivos de cada sitio. En 2017 y 2018 fue posible realizar un muestreo pormenorizado, por sitios, de ambas variables, lo que permitió un análisis espacial más detallado y la comparación por Clases (Gastropoda y Bivalvia) de los moluscos encontrados.
La fauna recolectada (organismos vivos excluyendo esponjas, corales y gorgonáceos, que no han sido reportados en la dieta de P. argus en Cuba) se seleccionó en un tamiz de 4 mm, se almacenó en formaldehido neutralizado al 10 % y se empleó para determinar los valores de densidad (ind/m2) y biomasa (g/m2) a partir del total de organismos, su peso y el área total (21 m2) de cada réplica. La biomasa (peso seco), se determinó mediante una balanza técnica de 0.01 g de precisión. Mediante microscopía estereoscópica se identificaron los organismos, hasta nivel de especie en caso de los moluscos y hasta el menor taxón posible los restantes. La identificación de moluscos se basó en las descripciones de Mikkelsen, y Bieler (2007) y Redfern (2013). El resto de los grupos se identificaron según criterios generales (Alcolado, 1990) y el apoyo de especialistas en comunidades bentónicas de la plataforma cubana.
Para los análisis, los organismos megazoobentónicos se separaron en dos grupos: moluscos (alimento principal de P. argus) y bentos (todos los organismos excluyendo moluscos). A falta de un sistema de clasificación que establezca un límite de tamaño superior para el megazoobentos, y asumiendo que, según el criterio de los autores, es poco probable que las langostas depreden organismos cuyo tamaño exceda los 6 cm de diámetro menor (cinco veces el tamaño de la boca de una langosta adulta), se contabilizaron sólo organismos menores a 6 cm.
Las variaciones en el potencial alimentario disponible para las langostas son una de las principales implicaciones de los cambios en la densidad y biomasa de bentos. Por esta razón, los datos del 2017 y 2018 se emplearon además para estimar la disminución del potencial alimentario a partir de los valores de densidad y biomasa de moluscos reportados por (Alcolado, 1990) en 1983. Ante la carencia de un índice que cuantifique la disponibilidad de alimento para P. argus, la variación en el potencial alimentario se estimó a partir del promedio de los porcentajes en que disminuyeron (entre 1983 y 2018) la densidad y la biomasa de moluscos, menos el 20 % de la disminución de otros grupos. Esta cantidad se definió basado en que, según Herrera et al. (1991), los moluscos constituyen el 80 % de la dieta de las langostas que se alimentan en el seibadal, por lo que otros grupos integrantes del megazoobentos sólo conforman el 20 % del alimento disponible.
Procesamiento estadístico: Mediante la prueba de Kolmogorov-Smirnov se comprobó que los datos no cumplen con las premisas de una distribución normal, por lo que se aplicó estadística no paramétrica con un nivel de significación de P < 0.05. Las comparaciones pareadas entre años y entre grupos taxonómicos se efectuaron mediante la prueba U de Mann-Whitney. Las comparaciones múltiples entre años y entre sitios se realizaron aplicando la prueba Kruskall-Wallis. Para estimar si las variaciones en la densidad y biomasa de megazoobentos (sin incluir moluscos), se relacionan con la densidad y biomasa (variables dependientes) de moluscos megazoobentónicos, se aplicó un análisis de regresión lineal simple. Para determinar la tendencia en ambas variables (densidad y biomasa), se construyó un diagrama de dispersión bidimensional para cada una de ellas, con los años (1983, 2007-2018) como variable independiente (eje X). La relación entre los puntos se describió mediante una recta.
Resultados
Los valores de biomasa y densidad de organismos del megazoobentos, y de moluscos en particular, resultaron bajos en 2017 y 2018 (Tabla 1). Tanto la densidad como la biomasa de bentos resultaron similares (U de Mann-Whitney, P= 0.744 y P= 0.489 respectivamente) entre 2017 y 2018, hecho que también se evidenció al comparar la densidad y biomasa de moluscos megazoobentónicos (U de Mann-Whitney, P= 0.681 y P= 0.285 respectivamente).
Tabla 1 Table 1 Biomasa y densidad del megazoobentos (≥ 4 mm) y de los moluscos con concha que forman parte de la comunidad megazoobentónica en los fondos blandos del golfo de Batabanó, Cuba Biomass and density of megazoobenthos (≥ 4 mm) and shell mollusks that are part of the megazoobenthic community in the soft bottoms of the Batabanó Gulf, Cuba
| 2017 | ||||||
| Media | Mediana | Mínimo | Máximo | V | D.E. | |
| Biomasa bentos (g/m2) | 5.49 | 1.84 | 0.16 | 34.70 | 59.04 | 7.68 |
| Densidad bentos (ind/m2) | 4.99 | 4.29 | 0.33 | 12.05 | 8.60 | 2.93 |
| Biomasa moluscos (g/m2) | 0.73 | 0.26 | 0.00 | 9.74 | 2.16 | 1.47 |
| Densidad moluscos (ind/m2) | 1.86 | 1.31 | 0.00 | 6.52 | 2.51 | 1.58 |
| 2018 | ||||||
| Biomasa bentos (g/m2) | 4.21 | 1.35 | 0.20 | 32.87 | 47.97 | 6.93 |
| Densidad bentos (ind/m2) | 6.12 | 4.10 | 0.48 | 24.95 | 27.16 | 5.21 |
| Biomasa moluscos (g/m2) | 0.91 | 0.35 | 0.01 | 5.46 | 1.65 | 1.29 |
| Densidad moluscos (ind/m2) | 2.52 | 1.41 | 0.10 | 22.00 | 12.09 | 3.48 |
V: varianza; D.E.: desviación estándar. / V: variance; D.E.: standard deviation.
El análisis espacial de la densidad promedio del 2017 y 2018, reveló que los sitios con mayores valores, tanto de megazoobentos como de moluscos megazoobentónicos, fueron 2, 3 y 4. Similar comportamiento se obtuvo también para la biomasa (Fig. 2).
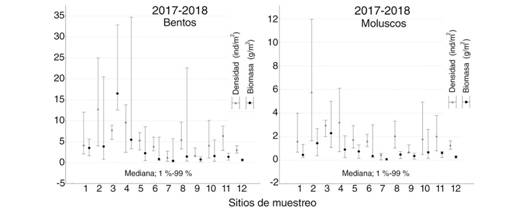
Fig. 2 Fig. 2 Densidad y Biomasa de megazoobentos (≥ 4 mm) y de moluscos megazoobentónicos en cada sitio muestreado en los fondos blandos del golfo de Batabanó, Cuba (2017 y 2018). Density and Biomass of megazoobenthos (≥ 4 mm) and megazoobenthic mollusks at each site sampled in the soft bottoms of the Batabanó Gulf, Cuba (2017 and 2018).
Los cambios en la densidad y biomasa de megazoobentos influyeron respectiva y significativamente en la densidad y biomasa de moluscos megazoobentónicos, tanto en la escala espacial como en la temporal. Se comprobó que las variaciones espacio-temporales de la comunidad de moluscos y del bentos fueron semejantes y mostraron una significativa relación de dependencia fundamentada en el resultado de las regresiones (Tabla 2).
Tabla 2 Table 2 Análisis de regresión entre la densidad y biomasa de megazoobentos (variables independientes) y la densidad y biomasa de moluscos megazoobentónicos (variables dependientes) en los 12 sitios muestreados en 2017 y 2018 (variación espacial) y como promedios anuales de los años estudiados (1983; 2007-2018) durante todo el período de muestreo (variación temporal) Regression analysis between the density and biomass of megazoobenthos (independent variables) and the density and biomass of megazoobenthic mollusks (dependent variables) at twelve sampled sites in 2017 and 2018 (spatial variation), and like annual average for each sampled year (1983; 2007-2018) during all study (temporal variation)
| 2017-2018 | ||||
| R | R2 | R2 ajustado | P | |
| Densidad bentos-Densidad moluscos | 0.977 | 0.954 | 0.954 | 0.001 |
| Biomasa bentos-Biomasa moluscos | 0.394 | 0.155 | 0.143 | 0.001 |
| 1983-2018 | ||||
| Densidad bentos-Densidad moluscos | 0.799 | 0.638 | 0.577 | 0.017 |
| Biomasa bentos-Biomasa moluscos | 0.807 | 0.652 | 0.594 | 0.015 |
El promedio anual de la densidad de organismos se incrementó en 2007 respecto a 1983 pero presentó tendencia a la disminución entre 2007 y 2018. Particularmente en moluscos (y= 12.3229-1.2095*x) esta tendencia resultó 54 % menos pronunciada que para el bentos (y= 22.7796-2.2346*x). En ambos grupos el mayor decrecimiento sucedió entre 2010 y 2015. La biomasa también mostró una tendencia a la disminución, con una pendiente 44 % mayor para el bentos (y= 12.9575-1.25*x) que para los moluscos (y= 7.2996-0.7038*x). Los valores de biomasa en 2007 fueron mayores a los existentes en 1983 excepto para la biomasa de bentos, caso en el que la tendencia al decrecimiento se observó a partir de 2010 (Fig. 3). Respecto a 1983, en 2018 la densidad de bentos mostró una significativa (P < 0.001) disminución del 73 %, sin embargo, la biomasa experimentó una disminución aún mayor (86 %). La densidad y biomasa de moluscos también mostraron un significativo decrecimiento (P < 0.001 en ambos casos), cuantificado en 55 y 88 % respectivamente. Con base en estos valores, entre 1983 y 2018 la disminución en el potencial alimentario disponible para las langostas se estimó en un: [(55 % + 88 %) / 2]- 0.2 [(73 % + 86 %) / 2]= 55.6 %.
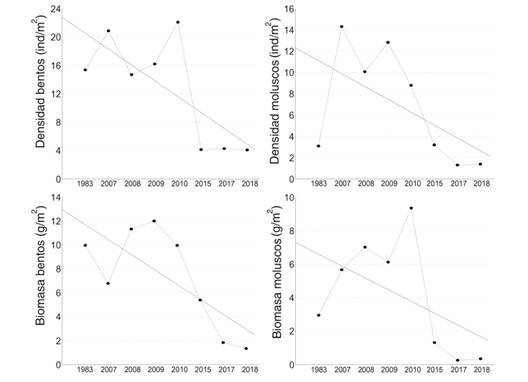
Fig. 3 Fig. 3 Densidad y biomasa de megazoobentos y de moluscos megazoobentónicos de fondos blandos en el golfo de Batabanó, Cuba. Density and biomass of megazoobenthos and megazoobenthic soft-bottom mollusks in the Gulf of Batabanó, Cuba.
En 2017 la densidad y biomasa de bivalvos fueron superiores (P= 0.001; P= 0.006 respectivamente) a las de gastrópodos, comportamiento que también se evidenció en 2018 (P= 0.001 y P= 0.011 respectivamente; Fig. 4).
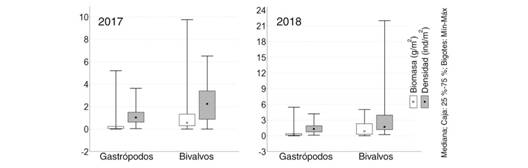
Fig. 4 Fig. 4 Densidad y Biomasa de los bivalvos y gastrópodos encontrados como parte de la comunidad megazoobentónica (≥ 4mm) en los fondos blandos del golfo de Batabanó, Cuba. Density and Biomass of bivalves and gastropods found as part of the megazoobenthic community (≥ 4mm) in the soft bottoms of the Gulf of Batabanó, Cuba.
Discusión
Según reportó Alcolado (1990), en el período 1981-1985, los sitios con mayor densidad y biomasa de megazoobentos (excluyendo esponjas, corales y gorgonáceos), fueron 5, 6, 8, 9 y 11. Según los resultados obtenidos en el presente estudio, tales sitios no destacan entre las de mejor condición, encontrándose mayores densidades y biomasas hacia la porción central del golfo (fundamentalmente sitios 2, 3 y 4). Esto puede ser consecuencia de los cambios que, a lo largo de más de tres décadas, han tenido lugar en esta región. Como ha sido reportado, tales cambios se traducen en el deterioro de las condiciones tanto bióticas como abióticas en las zonas pesqueras (Arias-Scheriber et al., 2008; Cerdeira-Estrada et al., 2008; Capetillo-Piñar, Villalejo-Fuerte, & Tripp-Quezada, 2015; Betanzos-Vega, Capetillo-Piñar, Lopeztegui-Castillo, Garcés-Rodríguez, & Tripp-Quezada, 2019). También se ha reportado deterioro en áreas próximas a la costa, que constituyen zonas de crianza para juveniles de varias especies de interés comercial y/o ecológico (Lopeztegui-Castillo & Capetillo-Piñar, 2008).
Las variaciones espaciales de la densidad y la biomasa corroboraron, además, una condición de deterioro generalizada en esta región. Entre los sitios de menores valores se encontraron algunos (sitios 6 y 12) próximos al borde de la plataforma, donde el mayor intercambio con aguas profundas (del Mar Caribe) debiera favorecer la existencia de mejores condiciones para el desarrollo la fauna bentónica (Martínez-Daranas, Betanzos-Vega, Lopeztegui-Castillo, & Capetillo-Piñar, 2018). No obstante, la condición más deteriorada se obtuvo en el sitio 7, más próximo a la costa y a la desembocadura del río Las Casas. Esto demuestra que el impacto antrópico en las zonas más próximas a las costas, muchas de las cuáles constituyen áreas de cría de especies comerciales como P. argus, es de elevada magnitud (Lopeztegui-Castillo & Capetillo-Piñar, 2008). Las variaciones temporales confirmaron una tendencia gradual al decrecimiento tanto de la abundancia como de la biomasa de megazoobentos. Los valores obtenidos en 2017 y 2018 fueron inferiores a los reportados por Alcolado (1990).
Además de implicar afectaciones a la biodiversidad, el deterioro de la comunidad megazoobentónica en el golfo de Batabanó puede traer como consecuencia una disminución en la productividad pesquera (Gómez, Ibarzábal, & Silva, 1980). La energía que ingresa a la trama trófica mediante organismos bentónicos disminuye con el decrecimiento de la densidad y la biomasa del megazoobentos. Esto afecta primera y directamente a organismos estrictamente bentófagos, como P. argus. Sin embargo, otros organismos de eslabones tróficos subsecuentes, como los depredadores de langosta, podrían llegar a ser impactados. La langosta P. argus es considerada un mesodepredador omnívoro que forma parte de la dieta de varias especies de interés económico o ecológico como pargos, chernas, pulpos, morenas y tiburones (Cruz et al., 1990; Briones-Fourzán et al., 2019). Estas especies podrían presentar variaciones en su crecimiento y hasta en sus patrones de abundancia y distribución, a causa de variaciones significativas en el megazoobentos que constituye alimento para las langostas. En los ecosistemas marinos, los mesodepredadores omnívoros juegan un papel determinante en la transmisión de la materia orgánica a través de la trama trófica (Long, Bruno, & Duffy, 2011).
Desde el año 1985 las capturas de langosta en Cuba, y particularmente en el golfo de Batabanó, han presentado una tendencia gradual al decrecimiento (Puga-Millán et al., 2013; Alzugaray-Martínez et al., 2018; Baisre, 2018). Aunque ya ha sido demostrado que tal decline tiene un origen multifactorial, sólo algunas de las causas han sido identificadas (Piñeiro-Soto, Arsenio-Areces, Puga-Milián, Cobas-Gómez, & de León-González, 2017; Alzugaray-Martínez et al., 2018). Las investigaciones existentes permiten concluir que desde mediados de la década de 1980, las comunidades bentónicas de fondos blandos en el golfo de Batabanó han experimentado un gradual y significativo deterioro. El presente estudio cuantifica este deterioro basado fundamentalmente en organismos que constituyen alimento para las langostas, haciendo evidente que otro de los factores que influye en la disminución de las capturas, es la reducción en la disponibilidad de alimento. Según Lozano-Álvarez, & Aramoni-Serrano (1996), el alimento puede influir determinantemente en la abundancia, distribución y condición nutricional de las langostas. Lopeztegui-Castillo, Capetillo-Piñar y Betanzos-Vega (2012), sugieren que las variaciones encontradas en la condición nutricional de P. argus en el golfo de Batabanó, se relacionaron con las afectaciones de la comunidad bentónica y la disminución de la disponibilidad de alimento en esta región.
El análisis de regresión practicado a los datos de 2017 y 2018, demostró que la densidad y biomasa de moluscos megazoobentónicos estuvieron relacionadas, respectivamente, con la densidad y biomasa de megazoobentos. Esta relación, estadísticamente significativa, se comprobó también al analizar los datos anuales del período en estudio (1983-2018). Consecuentemente, la densidad o biomasa de moluscos (> de 4 mm) de fondos blandos, podría ser utilizada para inferir la densidad o biomasa de megazoobentos, con un grado de certidumbre que varía entre 80 y 98 %. Similar resultado fue obtenido por Alcolado y Espinosa (1996), quienes encontraron una relación de r= 0.892 entre el número de especies de moluscos bentónicos y el número total de especies del bentos. Estas relaciones posibilitan estimar el estado de la comunidad bentónica a partir del estudio de sólo uno de sus componentes, los moluscos, relevantes además por su importancia como grupo bioindicador y como componentes de la trama trófica (alimento esencial para las langostas).
Dentro de los moluscos, los bivalvos ocuparon el mayor porcentaje, con mayores valores de densidad y biomasa en 2017 y 2018. Esta tendencia ya había sido reportada por Alcolado (1990) y parece ser una de las características de la comunidad de moluscos que no ha sufrido considerables variaciones, sugiriendo que los factores que determinan el deterioro del bentos en el golfo de Batabanó, impactan de manera similar a ambos grupos de moluscos. Ecológicamente, esto pudiera ser expresión de resiliencia y significar que la encontrada proporción favorece el equilibrio dinámico con el ambiente y resulta positiva para esta comunidad. Sin embargo, los gastrópodos han resultado ser más importantes que los bivalvos en la dieta de las langostas (Espinosa et al., 1990; Herrera et al., 1991; Martínez-Coello, Lopeztegui-Castillo, & Amador-Marrero, 2015), lo cual podría deberse a que se encuentran más accesibles dado que no se entierran en el sedimento.
Varias investigaciones, empleando diferentes grupos de organismos tanto de la meiofauna como de la macrofauna, han reportado afectaciones en la comunidad bentónica del golfo de Batabanó, particularmente sobre fondos blandos (Arias-Scheirber et al., 2008; Armenteros et al., 2008; Lopeztegui-Castillo, & Capetillo-Piñar, 2008; Armenteros et al., 2012; Hidalgo, Toledo, & Granados-Barba, 2015). Otros estudios han mostrado resultados similares basados principalmente en cambios de biotopo y las características de la vegetación submarina (Cerdeira-Estrada et al., 2008; Martínez-Daranas, Cano-Mallo, & Clero-Alonso, 2009; Martínez-Daranas, & Suárez, 2018; Martínez-Daranas et al., 2018). Por primera vez basado en atributos de la comunidad megazoobentónica, esencialmente densidad y biomasa, los presentes resultados corroboran los signos de deterioro y la degradación del bentos, lo que resulta novedoso y aporta relevante información al entendimiento de la dinámica de los ecosistemas de esta región.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

