











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
El género Montanoa Cerv. (Asteraceae) representa una fuente importante de extracción, síntesis y biosíntesis de metabolitos secundarios de interés farmacológico, con más de 70 productos naturales registrados (Villa-Ruano & Lozoya-Gloria, 2014). Montanoa tomentosa Cerv., comúnmente conocida como “zoapatle” por las raíces cihutal (mujer) y pathli (medicina), es una planta neotropical utilizada en la medicina tradicional mexicana como auxiliar en partos, regulador del ciclo menstrual, analgésico y anticonceptivo (Hahnn, Ericson, Lay & Probst, 1981; Lozoya-Legorreta, Velázquez-Díaz, & Flores-Alvarado, 1988; Carro-Juárez, Franco & Rodríguez-Peña, 2014). Es considerada recurso fitogenético no alimenticio por la Food and Agriculture Organization (FAO) (Molina-Moreno & Córdoba-Téllez, 2006). Su uso medicinal se atribuye a la actividad uterotónica y uterocontractora de los compuestos mayoritarios ácidos kaurenoico, monogenoico y grandiflorénico (Robles-Zepeda et al., 2006; Villa-Ruano, Betancourt-Jiménez & Lozoya-Gloria, 2009).
Estudios sistemáticos fundamentados en caracteres morfológicos de estructuras vegetativas (hojas y brácteas) y reproductivas (capítulos, flores y aquenios) agruparon a la especie en cuatro taxa infraespecíficos[M. tomentosa subsp. microcephala (Sch. Bip. ex K. Koch) V.A. Funk; M. tomentosa subsp. rosei (Rose ex B.L. Rob. & Greenm.) V.A. Funk; M. tomentosa subsp. tomentosa y M. tomentosa subsp. xanthiifolia (Sch. Bip. ex K. Koch) V.A. Funk] (Funk, 1982). Trabajos filogenéticos posteriores no mostraron diferencias entre las subespecies tomentosa,microcephala y xanthiifolia y no contemplaron la subespecie rosei (Plovanich & Paner, 2004), la cual actualmente se reclasificó a nivel de especie (Martin et al., 1998; van Devender, Sanders, Wilson & Meyer, 2000; Panero & Villaseñor, 2008).
En general, las variantes infraespecíficas de los organismos son resultado de procesos evolutivos (Manel, Schwartz, Luikart, & Taberlet, 2003), influenciados entre otros factores por la estructura geográfica y ambiental del paisaje en que se distribuyen (Paiaro, Oliva, Cocucci, & Sérsic, 2012). Esta interacción genera cambios en la formación y concentración de los productos naturales (Külheim et al., 2011), como ha sido registrado en otras plantas (Zhao et al., 2003; Chen et al., 2013; Chen et al., 2015).
La distribución de las especies puede calcularse mediante modelamientos predictivos con Sistemas de Información Geográfica (SIG) (Guisan & Thuiller, 2005). Estos modelos permiten generar estimaciones de nicho ecológico e indirectamente de distribución, basados en localidades de recolecta y observación georreferenciadas y variables ambientales como predictores (Phillips, Dudík & Schapire, 2019). Por lo anterior, este trabajo tuvo como objetivo estimar la distribución geográfica de M. tomentosa subsp. tomentosa y M. tomentosa subsp. xanthiifolia, así como identificar las variables ambientales asociadas a la formación de sus nichos ecológicos, dentro del territorio mexicano.
Materiales y métodos
Base de datos: Se elaboró una matriz de datos de registros de M. tomentosa subsp. tomentosa y M. tomentosa subsp. xanthiifolia a partir de la base del Herbario Nacional del Instituto de Biología de la Universidad Nacional Autónoma de México (IBUNAM: MEXU) (Apéndice Digital 1), complementada con registros utilizados en la revisión más extensa del género Montanoa (Funk, 1982) y otros obtenidos durante el desarrollo de este trabajo. Para estos últimos, las coordenadas se tomaron con un receptor de posicionamiento satelital GPS Garmin ETrex (Datum ITRF92). La georreferenciación, para casos de registros previos sin ubicación exacta, fue generada mediante Google Earth. Se realizaron recorridos de campo en diferentes localidades de los estados de Guanajuato, Querétaro, Hidalgo, Ciudad de México, Estado de México, Puebla, Oaxaca y Chiapas durante la etapa de floración y fructificación (octubre 2015 a marzo 2016). Las exploraciones fueron planeadas para cubrir la mayor parte de las regiones donde se han documentado registros de esta especie. Se lograron obtener 181 nuevos registros (Apéndice Digital 2). Esta información complementó la base de datos históricos inicial, con lo que se obtuvo un total de 314 registros.
Modelado de áreas potenciales de distribución: Se generó un modelo de idoneidad del hábitat con el software MaxEnt versión 3.4.1, el cual estima la probabilidad de distribución de las especies basada en el conjunto de localidades de recolecta y observación georreferenciadas y sus variables ambientales como predictores, mediante el método de máxima entropía y el método bayesiano (Phillips et al., 2019). Se utilizaron 22 variables (Tabla 1), 19 bioclimáticas con resolución espacial de 0.5 min de arco, obtenidas de la base de datos WorldClim (www.worldclim.org). Los datos digitales de elevación (DEM, 90 m de resolución) se obtuvieron de CGIAR-CSI (http://srtm.csi.cgiar.org), mientras que las capas en formato vectorial de uso de suelo y vegetación se tomaron de la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (INEGI, 2013). El régimen de humedad del suelo también se adquirió de la misma comisión (Maples-Vermeersch, 1992).
TABLA 1 TABLE 1 Variables ambientales y bioclimáticas utilizadas para determinar la distribución geográfica de Montanoa tomentosa subsp. tomentosa y Montanoa tomentosa subsp. xanthiifolia en México Environmental and bioclimatic variables used to determine the geographic distribution of Montanoa tomentosa subsp. tomentosa and Montanoa tomentosa subsp. xanthiifolia in Mexico
| Descripción de la variable | Unidades * |
| Temperatura media anual Ámbito de temperatura media diurna Isotermalidad Estacionalidad de la temperatura Temperatura máxima del mes más cálido Temperatura mínima del mes más frío Ámbito anual de temperatura Temperatura media del trimestre más lluvioso Temperatura media del trimestre más seco Temperatura media del trimestre más cálido Temperatura media del trimestre más frío Precipitación anual Precipitación del mes más húmedo Precipitación del mes más seco Estacionalidad de la precipitación Precipitación del trimestre más lluvioso Precipitación del trimestre más seco Precipitación del trimestre más cálido Precipitación del trimestre más frío Altitud Régimen de humedad Vegetación | ° C °C Adimensional CV ° C ° C ° C ° C ° C ° C ° C mm mm mm CV mm mm mm mm m ° 23 tipos |
* ° C = grados centígrados, CV = coeficiente de variación, m = metros, mm = milímetros.
Resultados
Análisis del nicho ecológico de Montanoa tomentosa en México: Se realizaron tres modelos de distribución con el programa MaxEnt 3.4.1. El primer modelo abarcó las dos subespecies estudiadas (Fig. 1A), el segundo se elaboró solo para M. tomentosa subsp. tomentosa (Fig. 1B) y el tercero para M. tomentosa subsp. xanthiifolia (Fig. 1C). Los valores del área bajo la curva del umbral acumulativo (AUC) para los datos de prueba de cada modelo fueron 0.988, 0.990 y 0.991, respectivamente. Asimismo, los valores de AUC para la comprobación de datos en cada modelo fueron 0.992, 0.996 y 0.998. Se obtuvo el valor máximo para la predicción al azar en todos los casos (0.5).
El área de máxima entropía calculada para las dos subespecies fue de aproximadamente 77 688 km2, que representa 4 % del territorio de la República Mexicana (Fig. 2A, 2B y 2C).
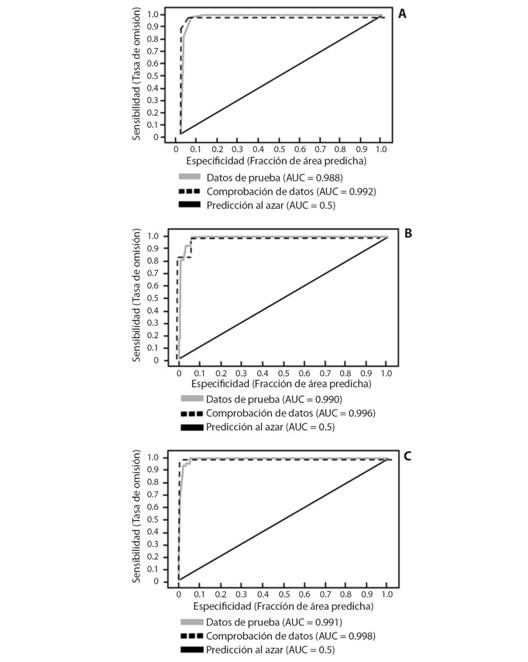
Fig. 1 Fig. 1 Sensibilidad y especificidad para los modelos de Montanoa tomentosa. La línea gris muestra el ajuste del modelo a los datos de información de la especie. La línea discontinua indica el ajuste del modelo a los datos de prueba. La sensibilidad frente a especificidad se muestra para A. dos subespecies estudiadas, B. M. tomentosa subsp. tomentosa y C. M. tomentosa subsp.xanthiifolia. Sensitivity and specificity for Montanoa tomentosa models. The gray line shows the fit of the model to the information data of the species. The dashed line indicates the fit of the model to the test data. Sensitivity to specificity is shown for A. two subspecies studied, B. M. tomentosa subsp. tomentosa and C. M. tomentosa subsp. xanthiifolia.
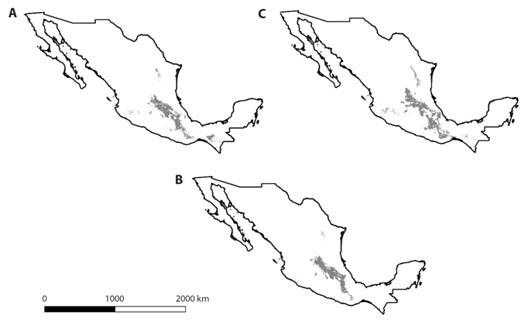
Fig. 2 Fig. 2 Distribución potencial de Montanoa tomentosa:A. dos subespecies, B. subsp. tomentosa y C. subsp. xanthiifolia, determinada a través del modelo de máxima entropía. En negro se observan las zonas de mayor potencial de distribución. Potential distribution of Montanoa tomentosa: A. two subspecies, B. subsp. tomentosa and C. subsp. xanthiifolia, determined through the maximum entropy model. In black, the areas of greatest distribution potential are observed.
Variables climáticas: De las 19 variables bioclimáticas utilizadas como predictores para el complejo de M. tomentosa, cinco contribuyeron con 70.2 % de la estimación total (Tabla 2).
TABLA 2 TABLE 2 Contribuciones porcentuales de las principales cinco variables bioclimáticas en el modelo MaxEnt para Montanoa tomentosa subsp. tomentosa y Montanoa tomentosa subsp. xanthiifolia en México Percentage contributions of the main five bioclimatic variables in the MaxEnt model for Montanoa tomentosa subsp. tomentosa and Montanoa tomentosa subsp. xanthiifolia in Mexico
| Variable | Contribución % |
| Estacionalidad de la temperatura Isotermalidad Temperatura máxima del mes más cálido Precipitación del trimestre más cálido Precipitación del trimestre más lluvioso Total | 21.8 17.7 13.2 9.6 7.9 70.2 |
En regiones del país con variación estacional de la temperatura de 14 a 26 % y precipitación media en el trimestre más cálido de 200 mm y de 300 mm en el trimestre más lluvioso, existe mayor probabilidad de presencia de la especie (por arriba de 0.6) (Fig. 3A, 3B y 3C). Además, en zonas donde la temperatura es superior a 30 °C durante el mes más cálido, la probabilidad disminuye hasta 0.3 (Fig. 3D). Los valores de isotermalidad (variación de la temperatura) óptimos para el desarrollo de ambas subespecies son de 63 a 72 unidades (Fig. 3E).
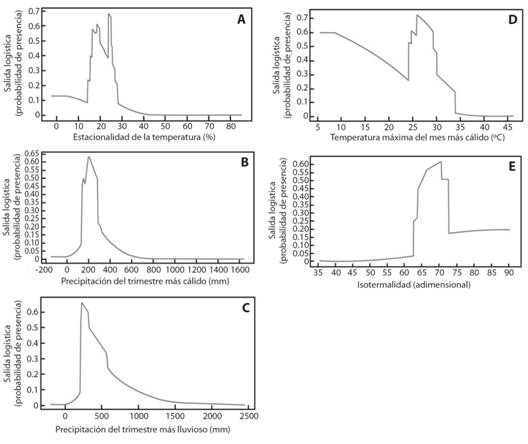
Fig. 3 Fig. 3 Variables bioclimáticas de mayor contribución en el modelamiento de máxima entropía para el conjunto M. tomentosa subsp. tomentosa y M. tomentosa subsp. xanthiifolia. A. Estacionalidad de la temperatura, B. Precipitación del trimestre más cálido, C. Precipitación del trimestre más lluvioso, D. Temperatura máxima del mes más cálido, E.Isotermalidad. Bioclimatic variables of greater contribution in the modeling of maximum entropy for the set M. tomentosa subsp. tomentosa and M. tomentosa subsp. xanthiifolia A. Seasonality of temperature, B. Precipitation of the warmest quarter, C. Precipitation of the wettest quarter, D. Maximum temperature of the warmest month, E.Isothermality.
Análisis por subespecie: Las variables de mayor impacto en el modelo de distribución de M. tomentosa subsp. tomentosa fueron precipitación en el trimestre más frío, isotermalidad y temperatura estacional. En zonas sin precipitación en el trimestre más frío, la probabilidad de presencia de la subespecie es superior a 0.9 y disminuye hasta 0.0 en regiones con precipitaciones superiores a 50 mm (Fig. 4A). Zonas con valores isotérmicos de 85 unidades tienen una probabilidad de presencia superior a 0.9, la cual disminuye hasta 0.2 en zonas con 60 unidades (Fig. 4B). En regiones donde la temperatura estacional es menor o igual a 10 % se estima una probabilidad de presencia de 1.0 de la subespecie tomentosa, la cual disminuye hasta 0.1 en regiones donde la estacionalidad de la temperatura es superior a 30 % (Fig. 4C).
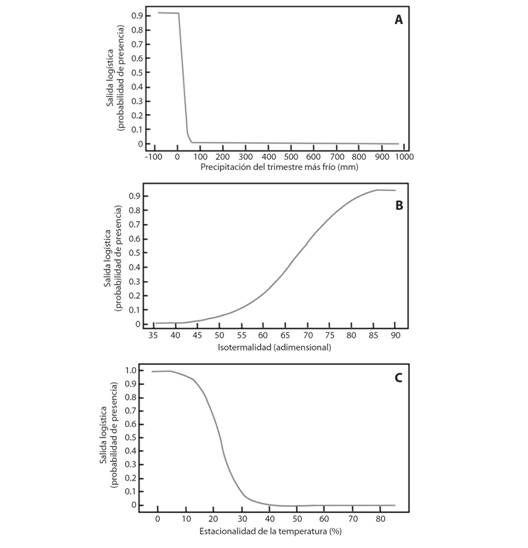
Fig. 4 Fig. 4 Variables bioclimáticas de mayor contribución en el modelamiento de máxima entropía para M. tomentosa subsp. tomentosa. A. Precipitación del trimestre más frío, B. Isotermalidad, C. Estacionalidad de la temperatura. Bioclimatic variables of greater contribution in the modeling of maximum entropy for M. tomentosa subsp. tomentosa A. Precipitation of the coldest quarter, B. Isothermality, C. Seasonality of temperature.
Las variables de mayor impacto en el modelo de distribución de M. tomentosa subsp. xanthiifolia fueron la estacionalidad de la temperatura, la isotermalidad y el ámbito anual de temperatura. En zonas con 20 % de variación en la temperatura estacional, la probabilidad de presencia de la subespecie es superior a 0.75 y disminuye a 0.2 en zonas con variación inferior o igual a 10 % y superior o igual a 30 % (Fig. 5A). En regiones con isotermalidad de 65 unidades existe una probabilidad de distribución de 0.6, la cual se reduce hasta 0.05 en zonas con valores por debajo de 55 o superiores a 85 unidades (Fig. 5B). En regiones con temperaturas que oscilan 23 °C, la probabilidad de presencia de la especie es de 0.6 y disminuye hasta 0.1 en regiones donde el ámbito supera 28 °C (Fig. 5C).
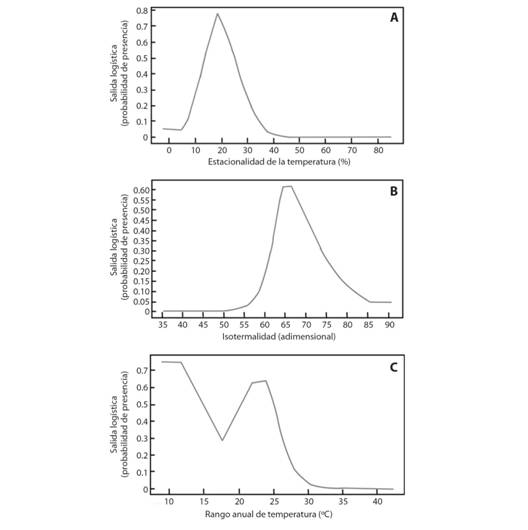
Fig. 5 Fig. 5 Variables bioclimáticas de mayor contribución en el modelamiento de máxima entropía para M. tomentosa subsp. xanthiifolia. A. Estacionalidad de la temperatura, B. Isotermalidad, C. Ámbito anual de temperatura. Bioclimatic variables of greater contribution in the modeling of maximum entropy for M. tomentosa subsp. xanthiifolia A. Temperature seasonality, B. Isothermality, C. Annual temperature range.
Otras variables: El complejo de M. tomentosa estudiado se ubica a una altitud que va de 500 a 2 500 m, pero con mayor frecuencia en un recorrido de 1 500 a 2 500 m. Los tipos de vegetación a los que mejor se asocia son la vegetación xerófila (matorrales desérticos y crasicaules), zonas con clima subhúmedo, ocupando la periferia y los claros de bosque tropical perennifolio y subcaducifolio, bosques de coníferas, bosques de latifoliadas y bosque mesófilo de montaña.
El régimen de humedad del suelo (RHS) predominante en los hábitats de esta especie es xérico y ústico (90 a 180 y 180 a 270 días de suelo húmedo, respectivamente), con precipitaciones de 400 a 1 200 mm y en menor probabilidad con RHS arídico y údico (menos de 90 y 270 a 330 días de suelo húmedo, respectivamente), con precipitaciones de 125 a 400 mm y de 1 200 a 2 000 mm.
Discusión
La base de datos de M. tomentosa, utilizada en los modelos de distribución, se generó a partir de reportes de presencia históricos y de visitas recientes a los sitios registrados de recolecta y observación. Las nuevas poblaciones georreferenciadas fueron seleccionadas de acuerdo con las claves sistemáticas utilizadas por Funk (1982). Esta información representa el registro más actualizado para ambas subespecies. El ámbito geográfico que abarcan los datos presentados cubre las condiciones ecológicas del área geográfica en que las subespecies pueden desarrollarse. Por esta razón, los resultados coinciden en gran medida con la distribución documentada previamente por Funk (1982).
El área bajo la curva de los tres modelamientos registró valores de AUC cercanos a la unidad, tanto para los datos de prueba como para la comprobación de datos. Los valores de AUC varían de 0 a 1, de tal modo que los superiores a 0.9 se clasifican como excelentes y los menores o iguales a 0.6 como inválidos (Hoffman, Narumalani, Mishra, Merani, & Wilson, 2008). Por esta razón, se considera que la capacidad predictiva de los modelamientos generados es apropiada. En este sentido, Elham, Mostafa, Mehdi y Mohammad (2015) mencionan que los valores de AUC tienden a ser menores para las especies con una amplia distribución geográfica.
Generalmente, los patrones de distribución son resultado de interacciones bióticas y abióticas, además de factores históricos ambientales y orográficos (Maciel-Mata, Manríquez-Morán, Octavio-Aguilar & Sánchez-Rojas, 2015), los cuales no son considerados por los algoritmos de modelado, pero pueden proporcionar información valiosa sobre orígenes históricos, relaciones con otras especies y capacidad potencial de colonización (Peterson, Ball, & Cohoon, 2002). Por ejemplo, el hábitat de M. tomentosa suele ubicarse en espacios abiertos, perturbados por el hombre, como asentamientos humanos, caminos o veredas y zonas de producción agrícola con remoción regular del suelo. Estas características son propias de especies con hábito de crecimiento ruderal y arvense (Martínez-De la Cruz et al., 2015), predominantemente heliófilas.
Lo anterior se observa en la enorme cantidad de especímenes de M. tomentosa recolectados en hábitats perturbados, de tal modo que el modelado de nichos ecológicos debe interpretarse como la representación geográfica de las condiciones ambientales requeridas por la especie, como han sugerido Peterson (2003) y Martínez-Meyer (2005) para otros organismos. Asimismo, los resultados mostraron zonas potenciales para el desarrollo de las dos subespecies, aunque no existe información biológica que respalde dicha distribución. Esto podría deberse a la ausencia de recolectas en estos sitios, como ha ocurrido con otras especies (Luna-Vega, Alcántara-Ayala, Contreras-Medina, & Ríos-Muñoz, 2012). Además, los modelos tanto de nicho ecológico idóneo como de amplitud de distribución de especies son de tipo probabilístico (Broennimann et al., 2006) y las especies no están presentes en todas las áreas potenciales de distribución, como lo destacan Soberón y Peterson (2005).
En este trabajo se amplía la información presentada por Funk (1982) sobre el tipo de vegetación y el gradiente altitudinal de la zona en que se desarrolla M. tomentosa y se establecen las características ambientales que favorecen la idoneidad de su hábitat.Funk (1982) menciona poblaciones atípicas de M. tomentosa subsp. xanthiifolia localizadas a 230 m y registros de M. tomentosa subsp. tomentosa a elevaciones mayores de 2 800 m. Estos datos difieren de los resultados obtenidos en el presente trabajo. El registro a muy baja elevación parece corresponder a un taxón diferente, por lo que los especímenes probablemente requieren de una revisión adicional.
En México, la mayor riqueza de especies de la familia Asteraceae se localiza en los bosques mesófilo, templado y tropical (Cruz-Cárdenas, Villaseñor, López-Mata, & Ortiz, 2013). En el caso particular de M. tomentosa, se observó que la presencia de ambas subespecies se limita a zonas de clima seco con matorral xerófilo desértico y crasicaule en el centro y sur del país, así como a comunidades abiertas dentro de zonas con predominancia templada, tropical y del bosque mesófilo de montaña (claros desprovistos de vegetación y periferia de esos bosques).
Los resultados obtenidos en el presente trabajo aclaran las principales características ambientales que delimitan el hábitat de M. tomentosa. La estacionalidad de la temperatura, isotermalidad y temperatura máxima del mes más cálido determinan la distribución de la especie. La precipitación del trimestre más cálido y la precipitación del trimestre más lluvioso definen el hábitat entre ambas subespecies al observarse un efecto negativo en la distribución de M. tomentosa subsp. tomentosa respecto a regiones con mayor precipitación.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

