











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
Among various abiotic stresses, light stress is emerging as one of the important factors that limits the biomass yield. This is important in the context of shrinking land area under cultivation and increasing demand for cropping systems to enhance productivity per unit area. Combining pastures with plantation crops or with forest trees is likely to increase the availability of forage and use the land more efficiently. The area under agro-forestry and silvipasture systems is fast increasing in India and around the world. The extent of agroforestry in India was estimated to be 14.46 million ha (excluding fallow land), whereas the potential area under agroforestry was estimated to be about 17.45 million ha (Newaj et al., 2017). Panicum maximum cultivars, particularly Petrie (Green Panic) and Embu (Creeping Guinea), have been used for grazing under coconuts (Stfir & Shelton, 1991; Plucknett, 1979). Shading intensity varies significantly in various agro-forestry, silvipasture, and social forestry systems. Shade intensity also varies according to the age of plantation. Hence, suitable understory crop for varying degree of shade tolerance is required.
Among many perennial grasses, Megathyrsus maximus (guinea grass), formerly known as Panicum maximum Jacq., has proven to be a high biomass producing grass in agroforestry and silvipasture systems. Singh and Chaturvedi (2011) identified the Alnus nepalensis, Stylosanthesis guyensis and P. maximum system as the best for high rainfall hilly regions with the potential of 13.5 tha-1(DM) fodder per year. The grass is known to be tolerant to shading conditions (Wong & Wilson, 1980; Kaushal, Malaviya, & Singh, 2000) and is recommended for cultivation in agroforestry and forestry systems.
This grass is a widely adapted perennial grass for utilization as fodder because of its ease of propagation, fast re-growth, high biomass and nutritional quality. Under rain-fed condition (400-1 500 mm rainfall per year), it yields 40-60 t/ha dry matter with over 14 % crude protein content and 41-72 % dry matter digestibility (Bogdan, 1977; Sukhchain & Sidhu, 1992). In vitro dry matter digestibility of organic matter from first harvest to third harvest is reported to vary from 56.9 to 87.7 % among different varieties (Aganga & Tshwenyane, 2004). It is also responsive to higher input conditions and also suitable for cultivated/irrigated conditions. The species is highly responsive to fertilizers, which results in an increased number of cuts of the grass (Pereira et al., 2012). Despite the latter information and being considered to be shade tolerant, some studies had reported different results. For instance, in some of the studies, shading decreased the yield in guinea grass with a significant increase in the N content under shade (Norton, Wilson, Shelton, & Hiu, 1991; Deinum et al., 1996). On the other hand, Lowry, Lowry and Jones (1988) reported an increase up to 250 %. Hence, there is particular need of studies that take into account the available genetic diversity to identify what genotype is more for shaded condition.
The crops’ primary gene pool exhibits high degree of variation for several traits including morphology, adaptation and biomass yield. Therefore, the morphological, biochemical, reproductive and physiological parameters were characterized and the distinct morphotypes were identified in the sets of germplasm used at the Indian Grassland and Fodder Research Institute in Jhansi, India (Jain, Roy, Kaushal, Malaviya, & Zadoo, 2003, Jain, Roy, Kaushal, Malaviya, & Zadoo,2006; Kaushal, Malaviya, & Singh, 1999; Kaushal et al., 2008; Malaviya, 1996, Malaviya,1998). This gene pool was considered ideal for studies that evaluate this factor under abiotic stress because such diverse lines may respond differently to the stress condition. Hence, in view of the expanding area under agroforestry/silvipastures and the necessity of identifying the shade adapted forage grasses, the present investigation tried to understand how the morphologically distinct genotypes of guinea grass respond to various shaded environments. Additionally, the correlation between morphology and the biomass yield under abiotic stress was also envisaged to be established.
Materials and methods
Site of study: The analysis was carried out at the experimental farm of ICAR - Indian Grassland and Fodder Research Institute (IGFRI) in Jhansi, India (25.27’ N & 78.35 E), 271 m.a.s.l. under rain-fed conditions. The annual rain-fall during the first and the second year of the experiment was 477 mm and 1 147 mm respectively. The rain-fall distribution was mainly during July to September. Minimum and maximum temperatures during the productive months (July to September) in the first and second year also ranged between 21-28°C and 29-38°C, respectively. The bright sunlight hours during the growth period (July, August and September) in the first year were 5.3, 4.0 and 8.0 hours/day, whereas during the second year they were 5.2, 5.1 and 2.7 hours/day, respectively. The sunlight intensity in open conditions at 12 pm in September was observed to be 582.3 mmol/m2/s.
Experimental design: The experiment was conducted by planting the genotypes in open as and shaded conditions. Three shaded conditions were created by using a shade net which intercepted 25 %, 50 % and 75 % of the sunlight. Shade nets were put at a height of 2 m and the crop was also covered from the sides. A total of 44 genotypes representing the collections from Ethiopia and India were studied. This material included exotic genotypes from Ethiopia (IG 97-6, IG 97-5, IG 97-14, IG 96-240, IG 97-35), North India (IG 01-83, IG 01-134, IG 01-180, IG 01-183, IG 01-181, IG 01-187, IG 01-141), Central India (IG 01-207-1, IG 01-207-2, IG 01-207-3, IG 01-207-4, IG 01-204-1, IG 01-204-2, IG 01-200, IG 01-87) and South India (IG 01-155, IG 01-212, IG 01-171, IG 01-221, IG 01-205, IG 01-191, IG 01-152, IG 01-98, IG 01-96, IG 01-93, IG 01-92, IG 01-89, IG 01-206, IG 01-201, IG 01-185, IG 01-176, IG 01-220, IG 01-198, IG 01-197, IG 01-190, IG 01-160, IG 01-122, IG 01-116, IG 01-85). The seeds containing those genotypes were procured from the gene bank of IGFRI, Jhansi, India. Nursery of the genotypes was raised and six week old seedlings were transplanted.
Raising of plants and observations: Genotypes were transplanted in 3 x 1 m plots that had 3 m long paired rows, spaced 50 cm apart and accommodating six tussocks spaced at 50 cm in the row. The plants were harvested above ground at 15 cm of height once during the first year of establishment and thrice during the second year in the rainy season. For identification of shade tolerant genotypes, cumulative yield over four cuttings was considered. Dry matter percent, at each harvest, was estimated by drying 500 g green sample of each accession at 60°C in hot air oven until obtaining a constant sample weight. Data on morphological observations such as plant height, leaf blade length and leaf blade width were recorded on three plants in each plot during the second year, just before the second harvest. Plant height was recorded as the total length of the longest tiller from the ground to the top (the tip of the last emerged leaf). The leaf width and length were recorded on the third leaf from the top of the three randomly selected plants. The first and the last plants in the row were not considered for this data recording. Chlorophyll content among genotypes in ambient light condition and that at three shade levels was recorded on three plants in each plot using CCM-200 by placing the third leaf from the top between the sensor points provided in the instrument. This data was recorded between 9 to 11 am.
Statistical analysis: Paired t-test, quadratic regression and correlation analysis were performed following standard statistical methods using MS Excel program. Ranking of genotypes was done based on their score for green and dry biomass. The analysis of variance and the interaction effect was calculated following Kharrati-Kopaei and Sadooghi-Alvandi (2007). The effect of ante-dependence was judged following Kenwood (1987). The correlation matrix and variance was generated on the residual basis. The t-test was performed on yield data within treatments over different cuttings and between the cumulative yield over the cuts with the yield in individual cuts. It was also performed between the two groups of the genotypes: the genotypes with an increase in yield up to 50 % shade and the remaining genotypes showing yield increase even at 75 % of shade.
Results
Biomass production under open vis-à-vis shaded condition: The total green forage yield (GFY), in four harvests for two years, under open conditions ranged from 14.9 to 77.5 MT/haacross the genotypes with a mean of 43.5 MT/ha. The mean green biomass increased to 73.8 MT/haand 100.7 MT/haunder 25 % and 50 % shade, respectively, and declined to 91.8 MT/ha under 75 % shading (Digital Appendix 1, Fig. 1). The paired t-test revealed significant difference (< 0.01 % level) only at 25 % and 50 % shading when comparing with the open and shading at 25 % respectively (Digital Appendix 1). Since the fresh and the dry biomass declined at 75 % shading, further correlations and the identification of genotypes was done considering yield in open vis-à-vis at 50 % shading conditions. There was a positive significant correlation between green biomass yield under open vis-à-vis at 50 % shading conditions. The quadratic trend shown in Fig. 1 and the equation in the Digital Appendix 1 also show the trend of decline at 75 % shade.
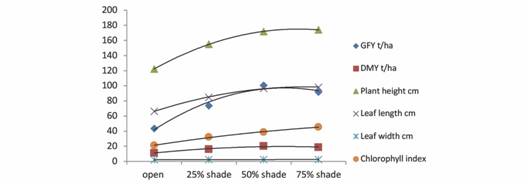
Fig. 1 Fig. 1 Yield and yield attributes of guinea grass grown under open and shaded conditions. (Lines connecting the data point are quadratic trend lines). Componentes de rendimiento y rendimiento del zacate guinea cultivados en condiciones abiertas y sombreadas (Las líneas que conectan el punto de datos son líneas de tendencia cuadráticas).
The dry matter yield (DMY), in the four harvests during two years, ranged from 3.9 to 19.46 MT/haunder ambient sunlight condition with a mean of 11.31 MT/ha across the genotypes. The mean dry matter production was 16.09 MT/ha, 20.42 MT/ha and 18.78 MT/ha, respectively, under the three shaded conditions (25 %, 50 % and 75 %) (Digital Appendix 1). However, there was no significant difference between the yields at 50 % shade and 75 % shade. The positive correlation was established between dry matter yields under open conditions with that under 50 % shaded condition. The quadratic trend shown in Fig. 1 and the equation in Digital Appendix 1 also show the trend of nominal decline at 75 % shade.
The interaction effect calculated (F = 2.78, d.f. = 78, 48) was highly significant (P = 0.000114) (Digital Appendix 2). Since the p-value was too low, the highly significant interaction effects for green forage yield among treatments and the genotypes were confirmed, as suggested by Kharrati-Kopaei and Sadooghi-Alvandi (2007). Similarly, for the dry forage yield, it was highly significant (F = 2.097, P = 0.003, d.f. = 78, 48).
In the repeated observation, the relations were likely to be serially correlated, hence, the residual correlation matrix was generated from the yield data observed at different cuttings. In the present study, the correlation pattern was not the typical for repeated measurements over different cuttings in open and shaded conditions (Digital Appendix 3). Since they did not follow a trend, it suggests that there were other factors influencing the regeneration such as seasonal variation, its regeneration ability and the genotypic differences. Similar trends were also observed for the correlation of dry matter productivity.
The t-test between yield at different cuttings with their preceding cutting yield were found to be significant in open as well as shaded conditions. The trend was similar for the GFY and dry matter yield (DMY) (Digital Appendix 4). On the other hand, the t-test between the first and second cutting for green fodder yield (GFY) and that between the third and fourth cutting for DMY were the only values with non-significant differences. For the ante-dependence test, the genotypes were grouped on the basis of biomass yield: genotypes that showed decline in yield under 50 % shade and those that showed yield increase even at 75 % shade. The t-test showed non-significant differences between the groups in open, 25 % shade and 50 % shaded condition. However, at 75 % shade, significant differences between the groups were observed (Digital Appendix 5). Nevertheless, the increase of variance over time was typical of cumulative yield data over the cuttings, which indicated that the covariance structure was not stationary.
Chlorophyll content andmorphological adaptation in shaded condition: The average plant height observed among genotypes in open condition was 122.3 cm, which increased to 154.3 cm and 172.0 cm respectively at 25 % and 50 % shading (Digital Appendix 1). Marginal numerical and non-significant increase of plant height to 174.0 cm was noticed at 75 % shading. The plant height showed positive correlation when the shading was increased. The average leaf length when the shading was increased to more than 50 %. There was no significant difference between leaf length at 50 % shade and 75 %. Additionally, the leaf length had a positive correlation and regression with increasing shade. The leaf width also had similar trends as the leaf length (Digital Appendix 1).
The genotypes grown under open and shaded conditions showed that the chlorophyll content, as indicated by CCI, increased with the shading intensity. Average CCI in the open environments was 20.9. It increased to 32.1, 38.7 and 45.4 respectively under 25 %, 50 % and 75 % shading conditions (Digital Appendix 1). On the other hand, there was no correlation between chlorophyll content in open vis-à-vis shaded conditions; however, there was linear increase in chlorophyll content with increasing shade.
Discussion
The mean green biomass increased under 25 and 50 % shading conditions, but declined under 75 % shading. However, there was no significant difference between yield at 50 and 75 % shade. Guinea grass under dense shade generally gives 27 % of light, according to previous stuides, and have 11 % higher biomass than under full sunlight conditions (Eriksen & Whitney, 1981). In this investigation, the increase in yield under shade was significantly higher, which follows the results of Wong and Wilson (1980) who found a 27 % increase in shoot yield and a 76 % increase in nitrogen yield of 8-week cut green panic (P. maximum var. trichoglume)grown under 40 % of light transmission, compared to full sunlight conditions. On the other hand, there was an increase in yield up to 250 % in guinea grass under the shade of the canopy trees Albizia lebbek, compared to those in full sun outside the canopy, according to Lowry and collaborators (1988).
In regards to experiments conducted under artificially created shaded condition with a 50 % cloth, Wilson, Catchpoole and Weier (1986) showed an increase in dry weight and nitrogen yield on green panic, similar to the one obtained in this study. Such an increase in biomass yield under shade is also reported for other tropical grasses. Wilson, Hill, Cameron and Shelton (1990) also reported a 35 % increase in growth for bahia grass (Paspalum notatum) under approximately 55 % light transmission within a plantation of Eucalyptus grandis trees, in addition to a 67 % increase in nitrogen yield of the herbage.
Most importantly, the genotypes that give high yield under open conditions were also more abundant under shaded condition, although it was not a consistent pattern. For instance, the genotypes IG 97-5 and IG 01-93, which ranked 8th and 15th respectively under open conditions, ranked first and second respectively under 50 % shade (Table 1). The high value of the regression coefficient observed for green biomass yields under open and the three shaded conditions suggests the involvement of additional factors affecting its yield under different light intensities. The top three genotypes (IG 01-92, IG 01-98 and IG 97-5), on the basis of overall score, also performed well under 50 % shaded condition. Dry matter accumulation also increased significantly to more than 50 % shade and showed a positive correlation between dry matter yields under open conditions and those under shade.
TABLE 1 TABLA 1 Top ranking genotypes based on their rank in green and dry matter yield in 50 % shade and open condition Genotipos de alto rango basados en su rendimiento de materia verde y seca en 50 % de sombra y la condición abierta
| Genotype | Rank in Green fodder yield | Rank in Dry matter yield | Total | Overall Rank | ||
| open | 50 % shade | open | 50 % shade | |||
| IG 01-98 | 3 | 3 | 2 | 3 | 11 | 1 |
| IG 01-92 | 2 | 6 | 3 | 6 | 17 | 2 |
| IG 97-5 | 8 | 1 | 8 | 1 | 18 | 3 |
| IG 97-6 | 5 | 4 | 5 | 8 | 22 | 4 |
| IG 01-89 | 4 | 13 | 4 | 10 | 31 | 5 |
| IG01-220 | 1 | 17 | 1 | 17 | 36 | 6 |
| IG 01-93 | 15 | 2 | 17 | 2 | 36 | 6 |
| IG 01-155 | 6 | 15 | 10 | 11 | 42 | 7 |
| IG 01-212 | 9 | 19 | 6 | 18 | 52 | 8 |
| IG 01-187 | 16 | 12 | 11 | 13 | 52 | 8 |
| IG 01-171 | 13 | 11 | 20 | 16 | 60 | 9 |
| IG 01-116 | 7 | 24 | 7 | 26 | 64 | 10 |
| IG 01-207-2 | 12 | 28 | 12 | 29 | 81 | 11 |
| IG 01-96 | 11 | 30 | 16 | 25 | 82 | 12 |
| IG 01-191 | 10 | 34 | 9 | 32 | 85 | 13 |
| IG 01-221 | 14 | 38 | 21 | 37 | 110 | 14 |
| IG 96-240 | 17 | 36 | 28 | 40 | 121 | 15 |
The higher herbage yields obtained by Odedire and Babayemi (2008) for P. maximum, when cropped with Leucaena leucocephala,was attributed to the ability of the grass to withstand the shading effect, since the dry matter (DM) production of tropical forage grasses is influenced by photoperiods by stimulating floral induction (Taiz, Zeiger, Møller, & Murphy, 2015). This might be the reason why there is a lower yield in open conditions compared to those in shaded conditions. The top three genotypes for dry matter yield under open conditions were identified as IG 01-220, IG 01-98 and IG 01-92, whereas, under 50 % shade condition, the top three genotypes were IG 97-5, IG 01-93 and IG 01-98. Thus, IG 01-98 performed well under open and shaded conditions.
In regards to the genotypic response to shaded conditions, the genotypes IG 97-5, IG 01-93 and IG 01-98 ranked first, second and third respectively for the green and dry biomass yield under 50 % shading conditions (Table 1). However, considering the overall ranking - based on the green and dry matter yield under open and shaded condition - IG 01-98 was the best followed by IG 01-92, IG 97-5, IG 97-6 and IG 01-89 in decreasing order. Of these genotypes, excepting IG 97-5 (which ranked 8th under open conditions), the rest of the genotypes were good both under open and 50 % shade conditions. This shows that the genotypes, irrespective of the percentage point reduction under shade, were a high yielder under shade intensity over 50 %. In contrast, the genotype IG 01-93, which ranked 15th and 17th respectively for green and dry matter yield in open conditions, ranked second for both traits under shaded conditions. This shows that IG 01-93 recorded a minimum percentage point reduction under shaded conditions. Such genotypes can be treated as lines to be specifically grown under shades, in addition to their inclusion in breeding programmes for development of shade tolerant lines.
The microclimate created by the shade has a direct impact on the wind speed, temperature regime, humidity, evapo-transpiration and soil moisture levels. Hence, these conditions are likely to impact the biomass production of any crop growing under shade. If the light intensity is not reduced below the threshold level, other factors will increase biomass production. In the present study, green and dry biomass of many genotypes increased under shading. It appears that the less light intensity under shade was compensated with higher moisture availability compared to open conditions. Araujo, Santos, Rodriguez, and Pezzopane (2018) have also identified the water availability index and the day length as key seasonal factor for dry matter production in guinea grass. Increase in yield under moderate light reduction has also been reported by Samarakoon, Wilson and Shelton (1990). Genotypic variation in guinea grass, involving a few genotypes, for shade tolerance was observed in earlier reports also (Kaushal et al., 2000).
Most of the genotypes were responsible for the increase in the plant height under shade over 50 % compared to open conditions. It was observed that most of the genotypes, which ranked in the top five for yield parameters under open conditions and shade, were also characterized by higher values of plant height, leaf length and leaf width than the general mean for their respective traits (Table 1). Increase in plant height of guinea grass under shade was also reported by Anita and Lakshmi (2014). However, the genotype IG 01-207-2, which showed maximum plant height in open conditions, was associated with highly reduced height under shade. On the other hand, the genotype IG 01-93, which ranked second for green forage yield (GFY) and dry matter yield (DMY) under shade, was not tall and exhibited further reduction in height under shade. However, as per visual observation, this genotype was marked for increased number of tillers under shade. Additionally, under shading conditions over 50 %, leaf length and leaf width showed an increasing trend. Leaves of the three genotypes among the top five ranking were longer and wider than most of the other genotypes, thus indicating that long and broad leaved genotypes may perform better in shaded environment.
Chlorophyll content is a crucial factor in plant growth. Non-destructive, in situ, optical techniques are widely used to provide a relative indication of leaf chlorophyll concentration (Parry, Blonquist, & Bugbee, 2014). We used CCM-200 for comparing chlorophyll content. It gives the ratio of transmission of radiation at 931 nm to that at 653 nm, which is defined as the chlorophyll content index (CCI) and requires chlorophyll prediction equations for getting chlorophyll content (Cerovic, Masdoumier, Ghozlen, & Latouche, 2012). However, as per requirement of the present study, CCI were directly used for comparison among genotypes as well as the change in CCI affected by shading conditions. According to our results, the average chlorophyll content increased with shading intensity, but there was no correlation between chlorophyll content in open vis-à-vis shaded conditions. The top ranking genotypes for yield parameters were not among the top ranking genotypes for CCI either. Also, no clear trend for increase or decrease of CCI in open vs shaded condition could be established as the genotypes behaved differently for this trait. Therefore, we were unable to make generalizations.
Among the top five efficient (better performing) genotypes, IG 01-98, IG 01-92, IG 01-89, IG 97-5 and IG 97-6, the first three belonged to collections from south Indian humid tropical environments, whereas the latter two were from an exotic collection from Ethiopia. For instance, the genotype IG 01-93, which ranked second for green and dry matter yield under shaded conditions, also was in the collection from South India. This genotype possessed high shade tolerance because, in open conditions, it ranked 15th and 17th respectively for green and dry matter production. All these south Indian genotypes possessed early flowering nature. Thus, it appears that, with early flowering nature, they are more adapted to shading tolerance. Genotype IG01-220 was considered highly susceptible to shade because it ranked first in open conditions for dry and green matter production; however, they ranked 17th under 50 % shading conditions. Similarly, many other genotypes, which ranked moderately high in open conditions, showed a high reduction in yield under shade (Table 1).
In general, P. maximum is reported to have high degree of shade adaptability (Wong, Sharudin, & Rahim, 1985). The species shows linear increase in leaf area with increasing shade (Durr & Rangel, 2000). Our results also confirm that, with increasing shade intensity, leaf-length and leaf-width augmented, but with differential genotypic response. P. maximum’s genotypes represent C3-C4 intermediate types but some of them are close to C3, while others are close to C4 anatomy (our unpublished anatomical study on these genotypes). Hence, a differential response to shade is expected.
In conclusion, the study clearly established the trend of some accession of genotypes being more suitable under shade than other. Thus, genotypes IG 01-98, IG 01-92, IG 97-5, IG 97-6 and IG 01-89 were successfully identified suitable both under shaded and open conditions, whereas IG 01-93 was identified as suitable only in 50 % shade conditions. Morphological features such as tall nature, long and broad leaves were associated with shade tolerance. The study also established appreciable variability for shade tolerance among genotypes under study which can be utilized further in breeding programmes. The genotypes that were identified can be valuable genetic resources to be utilized in India, Africa and any other place with iso-climatic conditions.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.
Digital Appendix 1
