











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
La región de alta montaña de la zona Andina Neotropical representa uno de los ecosistemas más especiales del planeta (Madriñán, Cortés, & Richardson, 2013). El Páramo,siendo un área tan pequeña comparada con otros ecosistemas como el Bosque Tropical, alberga gran cantidad de especies de las cuales una amplia proporción corresponde a especies únicas (Gonzalez-Duran, 2016). El Páramo es considerado como centro de diversificación para diferentes grupos taxonómicos (Madriñán et al., 2013; Rangel, 2000; Vuilleumier, & Monasterio, 1986), además de ser reconocido por su alta complejidad geológica; la cual está ligada directamente a la orogénesis andina del Bloque Norte de los Andes (Buytaert, Iniguez, & De Bievre, 2007). Sin embargo, no solo sus particularidades históricas y biológicas lo han hecho atractivo para las investigaciones en diferentes ramas, sino también su fragilidad a los cambios abruptos del ambiente, su papel en el desarrollo cultural de las poblaciones (Morales-Rivas et al., 2007) y su capacidad de actuar como reservorio de agua (Cortés-Duque & Sarmiento, 2017).
La palabra páramo proviene del latín parămus, que significa “terreno yermo, raso y desabrigado, lugar frío y desamparado” (RAE, 2014); este es un concepto europeo, aplicado por los exploradores españoles a un bioma tropical, debido a su semejanza con las llanuras áridas de la península Ibérica (Luteyn, 1999). Actualmente, Páramo es definido como un ecosistema húmedo tropical ubicado a partir del límite superior del bosque con vegetación propia, de tipo herbácea y arbustiva (Hofstede, Segarra, & Vásconez, 2003). Los Páramos se distribuyen en las regiones de alta montaña de Ecuador, Colombia, en Venezuela sobre la región de Mérida, Panamá y Costa Rica (Hofstede et al., 2014; Luteyn, 1999). En el Bloque Norte de los Andes, su origen data del Plioceno, época en la cual las cordilleras alcanzaron su altitud actual (Van der Hammen, 1997) y su vegetación ya era propia del Páramo contemporáneo (Wijninga, 1996), pero con su límite altitudinal por debajo (1 700 m.s.n.m.) del límite actual (2 800 - 3 000 m.s.n.m.) (Flantua & Hooghiemstra, 2018; Van der Hammen & Gonzáles, 1963). No obstante, dicho límite ha sido dinámico (Flantua, O’dea, Onstein, Giraldo, & Hooghiemstra, 2019), ya que durante el Cuaternario (2.4 Ma) y hasta el presente, han ocurrido entre 22 a 24 ciclos glacial-interglacial (Diazgranados & Barber, 2017), permitiendo que islas de páramos se conectaran y se separaran durante estos periodos de tiempo, impulsando el proceso de especiación y de generación de especies endémicas (Benham, Cuervo, McGuire, & Witt, 2015; Jørgensen & Ulloa, 1994; Quintero, Ribas, & Cracraft, 2012; Van der Hammen & Cleef, 1986).
Son diversos los acercamientos que se han tenido en cuenta para definir Páramo, lo cual ha resultado en una plétora de delimitaciones que son, o no, parcialmente congruentes entre sí. Dichas definiciones están sujetas al objeto de estudio del investigador y solo tienen en cuenta factores de fácil reconocimiento y/o medición. Por ejemplo, Cuatrecasas (1958) afirma que los páramos son regiones de gran extensión ubicadas arriba del Bosque Andino entre 3 800 m.s.n.m hasta 4 700 m.s.n.m, dividiéndose en Subpáramo, Páramo y Superpáramo de acuerdo a sus características florísticas. Otros autores han considerado en conjunto factores bióticos y abióticos, tales como la altitud y algunas comunidades de plantas; siendo estos últimos los más utilizados para delimitar esta región en términos ecológicos (Guhl, 1982; Monasterio, 1980; Rangel, 2000; Sarmiento et al., 2013; Vargas & Pedraza, 2004).
Morrone (2017) plantea Páramo como “... of the Andean cordillera of Venezuela, Colombia, and Ecuador, from the upper forest line at 3 000 - 3 500 meters upwards and below the permanent snowline at ca. 5 000 meters. This high-elevation biome has been influenced by glaciations, being the landscape irregular, from very rough to flat and stretches from 3 000 meters to the perennial snowline at altitudes of 4 800 - 5 000 meters” (p. 218). Este es uno de los primeros acercamientos que plantea Páramo como una unidad histórica, y en definirlo formalmente en términos de biogeografía (Morrone 2001, Morrone 2014, Morrone 2017). Sin embargo, dicha propuesta se basó en el compendio del conocimiento previo de múltiples autores y no en una hipótesis a testear y replicable bajo métodos cuantitativos. Otros trabajos han evaluado Páramo a nivel biogeográfico (Jiménez-Rivillas, García, Quijano-Abril, Daza, & Morrone, 2018), sin embargo se basan en métodos usualmente criticados, los cuales requieren definir áreas a priori y violan la premisa de no jerarquía en la definición de área de endemismo en biogeografía histórica (Szumik, Cuezzo, Goloboff, & Chalup, 2002), o simplemente no presentan una delimitación clara de Páramo, ni se basan en la definición de áreas de endemismo (Hazzi, Moreno, Ortiz-Movliav, & Palacio, 2018). Adicionalmente en estos trabajos, las zonas de alta montaña de Costa Rica y el norte de Perú siempre son excluidos de los análisis, probablemente, porque no son parte de la definición de Van der Hammen (1974), trabajo en el cual se han basado la mayoría de estudios de Páramo. Por tanto, un reanálisis, bajo métodos de optimización, de toda la extensión del área que históricamente se ha considerado como Páramo, es relevante para poder reconocerla como unidad biogeográfica.
Un área de endemismo se define como “the congruent distributional limits of two or more species” (Platnick, 1991). Sus límites están marcados generalmente por eventos vicariantes y podrían ser difusos por efecto de expansión del rango de distribución de las especies y su dinamismo a través del tiempo (Zunino & Zullini, 2003). Para el reconocimiento de unidades biogeográficas se ha implementado diferentes algoritmos como: el Análisis de Parsimonia de Endemismos (PAE) (Rosen & Smith, 1988), el Análisis de Parsimonia de Endemismos para cuadrículas (PAE) (Morrone, 1994), el criterio para el reconocimiento de áreas de endemismo (Harold & Mooi, 1994), el criterio de optimización para evaluar métodos de endemismo (Linder, 2001), el Análisis de Endemicidad (AE) (Szumik et al., 2002; Szumik & Goloboff, 2004), el Análisis de Co-presencia Significante de Taxones (Sigcot) (Mast & Nyffeler, 2003), Sigcot con PAE (Giokas & Sfenthourakis, 2008), el Análisis de Endemismos de Áreas Anidadas (NAEA) (Deo & DeSalle, 2006), el Método de Análisis de Redes (NAM) (Dos Santos, Fernández, Cuezzo, & Dominguez, 2008) y la Interpolación Geográfica de Endemismo (GIE) (Oliveira, Brescovit, & Santos, 2015).
De los anteriores, el análisis de endemicidad propuesto por Szumik et al. (2002) y Szumik y Goloboff (2004), ha sido el más aceptado para el análisis de endemismo en los últimos años, dadas sus ventajas sobre los otros métodos (Alvez-Valles et al., 2018; Martínez-Hernández et al., 2015; Sánchez, Garcia-Cruz, Espejo-Serna, & Lopez-Ortega, 2019; Zhuang et al., 2018). Este reconocimiento se debe a que tiene en cuenta de manera explícita el componente espacial, además de identificar áreas superpuestas, núcleos y límites difusos (Aagesen, Szumik, & Goloboff, 2013; Casagranda, Taher, & Szumik, 2012; Martínez-Hernández et al., 2015; Szumik et al., 2002). Pese a su importancia y amplio reconocimiento en la comunidad, hasta el momento no se ha utilizado como herramienta para evaluar exclusivamente las zonas de alta montaña del Bloque Norte de los Andes, Mérida y Talamanca, y es así que surge la idea de determinar si los páramos neotropicales son una o varias unidades biogeográficas.
Datos: A partir de listas de referencia, como Luteyn (1999) o Rangel (2000), identificamos las clases y divisiones taxonómicas con mayor número de especies presentes en lo que hasta el momento se ha definido como Páramo. Teniendo en cuenta los límites de distribución de páramo (11°N, 8°S, 84°W, 70°W) propuestos en Luteyn (1999) descargamos los datos disponibles de los diferentes taxones seleccionados (Aves, Amphibia, Mammalia, Reptilia, Marchantiophyta y Spermatophyta) desde la plataforma de información de biodiversidad global GBIF. Además, revisamos la taxonomía y validez del nombre de cada especie en bases de datos como The Integrated Taxonomic Information System [ITIS] (Shaw, 2004) y Tropicos (Tropicos, 2018). Posteriormente, de acuerdo a las recomendaciones de Chapman (2005), realizamos una limpieza exhaustiva de datos, siguiendo el protocolo propuesto por Romero-Alarcon (2016) e implementado en el paquete biogeoclean (Romero-Alarcon & Miranda-Esquivel, 2015) de R (R Core Team, 2019). Primero, obtuvimos una lista de especies con distribución sobre los 1 900 m.s.n.m (altura mínima de distribución de los páramos), y eliminamos las especies exóticas e invasoras (IBIS (Kells & Worswick, 1997) e ISSG (Lowe et al., 2000)). Luego, con esta nueva lista de especies, descargamos de GBIF todos los puntos georeferenciados para cada una de las especies, eliminamos puntos mal ubicados, especies ampliamente distribuidas, y especies con menos de tres presencias; para finalmente, obtener especies con presencias viables para nuestros análisis.
Búsqueda y visualización de áreas de endemismo: Usamos como partición cada grupo taxonómico previamente mencionado. Asimismo, estos taxones se agruparon para generar particiones adicionales como: plantas totales (Plantas-T: Marchantiophyta + Spermatophyta), animales totales (Animales-T: Aves + Amphibia + Mammalia + Reptilia) y evidencia total (Plantas-T + Animales-T). En total en nuestro estudio usamos nueve particiones, las cuales se analizaron de manera independiente para evidenciar el aporte diferencial de cada una de ellas al patrón general.
Para identificar áreas de endemismo sensuPlatnick (1991), utilizamos el criterio de optimalidad propuesto por Szumik et al. (2002) y Szumik y Goloboff (2004) e implementado en el algoritmo eNDeMism/ Visual of eNDeMism (NDM/VNDM) (Goloboff, 2005). Para ello, realizamos búsquedas heurísticas de 10 repeticiones, manteniendo áreas con dos o más especies endémicas y con índice de endemicidad mayor o igual a 2.0, además de conjuntos de subóptimos con 0.90 de ajuste mínimo. La semilla de aleatorización fue 1.0 y todas las áreas se mantuvieron independientemente del porcentaje de solapamiento entre especies únicas. Estos parámetros fueron seleccionados siguiendo el análisis de Escalante, Rodríguez-Tapia, Szumik, Morrone, & Rivas (2010) para la delimitación de áreas de endemismo en la región Neártica. Siguiendo las recomendaciones de Aagesen, Szumik, Zuloaga y Morrone (2009) y Szumik et al. (2012), cada partición la evaluamos independientemente bajo dos tamaños de cuadrícula diferentes 0.25° y 0.5°; no usamos tamaños como 1° o mayores ya que superan la extensión del páramo más grande a evaluar. Finalmente, resumimos la información obtenida usando el consenso estricto de similitud de especies bajo tres cortes distintos: 30, 50 y 75 % (Aagesen et al., 2013).
Comparación de áreas con propuestas de otros autores: Construimos un vector espacial tipo polígono con cada una de las definiciones previas de Páramos: Cuatrecasas (1958), Jiménez-Rivillas et al. (2018), Monasterio (1980), Morrone (2017), Rangel (2000), Van der Hammen (1997); y Vargas y Pedraza (2004). Cada polígono fue generado extrayendo las curvas de nivel a partir de un modelo de elevación digital (DEM, por sus siglas en inglés) y proyectando estas como mapas vectoriales con ayuda de QuantumGis (QGIS Development Team, 2019). Para comparar numéricamente las definiciones previas con los consensos obtenidos, calculamos el área de sobreposición entre ellas usando la función gArea del paquete de análisis espacial Rgeos (Bivand, Rundel, Pebesma, Stuetz, & Hufthammer, 2016). El valor de la sobreposición para cada una de las comparaciones se escaló sobre el área total de cada área de endemismo encontrada.
Resultados
Después de realizar la actualización taxonómica y depurar las presencias obtenidas desde GBIF, encontramos 7 025 especies (570 Amphibia, 119 Reptilia, 272 Aves, 70 Mammalia, 189 Marchantiophyta y 5 725 Spermatophyta) con 193 250 presencias viables para nuestros análisis. De estas, 954 especies (116 Amphibia, 30 Aves, 8 Mammalia, 29 Reptilia, 5 Marchantiophyta y 766 Spermatophyta) soportaron al menos un área del total de las áreas de endemismo recuperadas bajo el análisis de evidencia total (Apéndice Digital).
Los diferentes tamaños de cuadrícula nos permitieron encontrar grupos de áreas similares, sin embargo, con el tamaño de 0.25° los identificamos con mayor resolución. Lo mismo sucedió con los cortes de consenso; aunque los tres (30, 50 y 75 %) fueron aplicados para resumir cada análisis, el corte de 75 % proporcionó la mejor interpretación de los resultados; además, con este valor (75 %) diferenciamos áreas de evidencia total soportadas por especies pertenecientes a una misma clase o división taxonómica. Cuando comparamos los resultados obtenidos con ambos tamaños de cuadrícula, el 99.4 % de las áreas encontradas con tamaño de 0.25° estuvieron contenidas en las encontradas con tamaño de 0.5°. Así mismo, cuando resumimos los resultados en cada análisis, todos los tipos de corte del consenso estricto recuperaron los mismos conjuntos de áreas, pero cada uno con número diferente de áreas consenso.
El número de arreglos iniciales fue diferencial entre tamaños de cuadrícula; por ejemplo, el análisis de la partición de evidencia total para 0.25° generó 445 arreglos, en contraste a los 1 040 arreglos encontrados bajo 0.5°. De la misma forma, encontramos 281 áreas consenso al resumir los resultados de 0.25°, comparadas a las 541 encontradas para 0.5°, ambos, bajo un corte de 75 %. Esto mismo ocurrió con otras particiones como Animales-T con 100 arreglos iniciales y 58 áreas consenso, o Plantas-T con 328 arreglos iniciales y 208 áreas consenso, ambos bajo un corte de 75 % (Tabla 1). Por tanto, el tamaño de cuadrícula influyó directamente en la cantidad de arreglos iniciales y en el número de áreas consenso en todos los análisis realizados. Pese a este comportamiento, el patrón final o sectores del mismo fueron constantemente recuperados entre todas las particiones.
TABLA 1 Número de arreglos iniciales de cada partición por tamaño de cuadrícula
| Partición | Número de arreglos iniciales | |
| 0.25° | 0.5° | |
| Mammalia | 1 | 7 |
| Marchantiophyta | 1 | 8 |
| Reptilia | 6 | 16 |
| Aves | 19 | 28 |
| Amphibia | 60 | 180 |
| Animales-T | 100 | 209 |
| Spermatophyta | 352 | 889 |
| Plantas-T | 328 | 929 |
| Evidencia total | 445 | 1 041 |
Unidades biogeográficas dentro de Páramo: Nuestros análisis permitieron identificar dentro de lo que actualmente es denominado como Páramo, ocho unidades biogeográficas: Santa Marta-Perijá, Mérida, Santanderes-Boyacá, Cundinamarca, Cordillera Central-Occidental, Norte de Ecuador, Centro-Sur de Ecuador y Talamanca (Fig. 1). Dichas unidades se asignaron al nivel de subprovincia, según la taxonomía propuesta para áreas biogeográficas (Ebach, Morrone, Parenti, & Parenti, 2008; Escalante, 2009). Estas áreas fueron comparables a la zonificación propuesta por Jiménez-Rivillas et al. (2018), trabajo del cual extrajimos la nomenclatura para asignar los nombres de nuestras subprovincias.
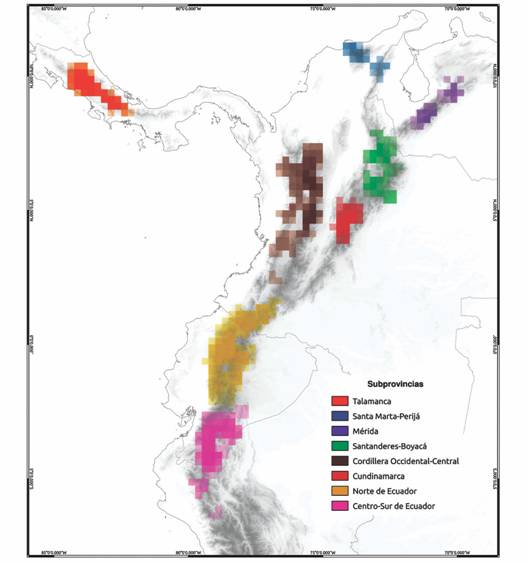
Fig. 1 Distribución geográfica de las áreas de endemismo (AE) en la provincia biogeográfica de Páramo para el tamaño de cuadrícula 0.25°. Consenso estricto 75 %, análisis de Evidencia total. La intensidad de color indica el nivel de superposición de las áreas consenso.
En términos de particiones, Plantas-T (Fig. 2A) y Animales-T (Fig. 2B) identificaron regiones semejantes a las subprovincias encontradas por la partición de evidencia total. Sin embargo, las regiones recuperadas de manera individual no fueron equivalentes entre ellas. Por ejemplo, Plantas-T recuperó, aunque no en su extensión completa, el patrón general de todas las subprovincias planteadas en este estudio (Fig. 1); dado principalmente por las especies de Spermatophyta (Fig. 2D) las cuales representaron el 97 % de la partición de Plantas-T, el 3 % restante correspondió a especies de Marchantiophyta (partición que solo participó en la identificación de una única área) (Fig. 2C). Algunas subprovincias como Talamanca, Santa Marta-Perijá, Mérida, Cordillera Central-Occidental, Norte de Ecuador y Centro-sur de Ecuador, fueron recuperadas por la partición Animales-T (Fig. 2B), pero a diferencia de Plantas-T (Fig. 2A), cada una de las particiones por clase (Aves, Amphibia, Mammalia y Reptilia) reconoció diferentes subprovincias de manera individual (Fig. 2E, Fig. 2F, Fig. 2G, Fig. 2H). Por ejemplo, la partición de Animales-T fue la única que recuperó la subprovincia de Mérida (Fig. 2B), en comparación a las encontradas por cada clase taxonómica de manera individual.
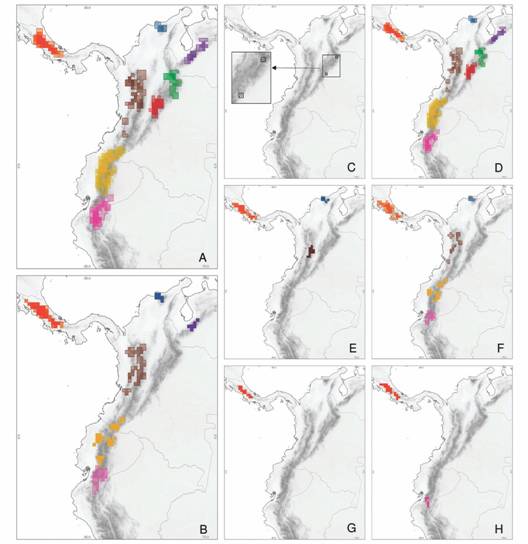
Fig. 2 Distribución geográfica de áreas de endemismo por partición (tamaño de cuadrícula 0.25°, consenso estricto 75 %): (A) Plantas-Total, (B) Animales-Total, (C) Marchantiophyta, (D) Spermatophyta, (E) Aves, (F) Amphibia, (G) Mammalia, (H) Reptilia. Los colores indican cada una de las subprovincias según la Fig. 1. La intensidad de color no representa el nivel de superposición de las áreas consenso, y el color del borde no representa los límites de las áreas.
Especies de Spermatophyta soportan el 98 % de las áreas consenso encontradas en el análisis de evidencia total, mientras que especies de Amphibia soportan el 61 % de las áreas. El resto de grupos aportaron en valores inferiores al índice de endemicidad de las áreas encontradas (Aves: 23 %, Mammalia: 14 %, Reptilia: 13 %), siendo Marchantiophyta el grupo con menor contribución (6 %) al soporte de las áreas consenso. Es así que cada grupo taxonómico contribuyó diferencialmente a la recuperación de cada una de las subprovincias, tanto de manera individual como colectiva (verEvidencia total).
Comparación de polígonos: Nuestra propuesta comparte un gran porcentaje de similitud (66 %) con las áreas planteadas por Jiménez-Rivillas et al. (2018). No obstante, dos de las propuestas más aceptadas: Morrone (2017) y Cuatrecasas (1958), utilizadas frecuentemente en trabajos de ecología, biogeografía y conservación, presentaron los menores valores de similitud con nuestros resultados (16 % y 4 % respectivamente). De igual forma, otras definiciones como las de Monasterio (1980), Van der Hammen (1997) y Rangel (2000) presentaron similitudes bajas respecto a nuestro trabajo (20 %, 26 %, y 15 %, respectivamente).
Discusión
Presencias a partir de bases de datos: Nuestro estudio es el primero en el cual se emplean alrededor de 7 000 especies de distintos taxa para la identificación de áreas de endemismo en zonas de alta montaña, en contraste a otros estudios como Jiménez-Rivillas et al. (2018) con 4 644 especies o Londoño, Cleef y Madriñán (2014) con 2 158 especies.
Trabajos con amplio muestreo de presencias, obtenidos a partir de bases de datos como GBIF, implican un reto en su manipulación, limpieza y análisis. Generalmente dichas presencias son propensas a diferentes tipos de errores, los cuales es nuestro deber detectar y corregir para minimizar el sesgo o la incertidumbre de nuestros datos finales (Antonelli et al., 2014; Chapman, 2005). Por ello, un componente esencial de nuestro trabajo fue depurar las descargas crudas de GBIF hasta obtener registros viables para nuestro análisis. Esta actividad es central, ya que una buena práctica de limpieza de datos debería ser el primer paso del diseño experimental, para cualquier estudio biogeográfico o que utilice datos distribucionales (Zizka et al., 2019). De esta forma, la información obtenida desde plataformas como GBIF podría ser mejor aprovechada para la obtención de patrones generales de biodiversidad (García-Roselló et al., 2015; Maldonado et al., 2015).
Tamaños de cuadrícula y particiones: Aunque no existe un tamaño de cuadrícula óptimo, otros estudios sobre áreas de endemismo, como Casagranda et al. (2009, 2012), recomiendan el uso de cuadrículas de diferentes tamaños para visualizar distintos patrones de áreas. Con el tamaño de 0.25° obtuvimos una mayor resolución de los límites de cada subprovincia, esto fue congruente con lo planteado por Szumik et al. (2012), quienes afirman que el tamaño de cuadrícula podría depender de la topografía del terreno, siendo las cuadrículas pequeñas las más adecuadas para el análisis en terrenos con pendiente alta, como por ejemplo en los Andes argentinos; idea que puede ser extendida fácilmente a nuestro trabajo.
De la misma forma que en Casagranda, Roig-Juñent y Szumik (2009) y Szumik et al. (2012), el número de arreglos iniciales fue proporcional al tamaño de cuadrícula, sin embargo, independientemente de este comportamiento, cada análisis nos permitió observar áreas equivalentes entre sí, indicándonos la consistencia del patrón general.
Trabajos como Escalante et al. (2010) proponen que el análisis de endemicidad se debe realizar con distintos taxa para que el estimador del índice de endemicidad sea robusto, con el cual se pueden hacer afirmaciones del patrón general de áreas. Por tanto, los resultados por clases taxonómicas son importantes para hacer inferencias y entender patrones individuales. Así, estos patrones individuales nos permiten ver el aporte proporcional de cada partición al patrón general. Por ejemplo, particiones como Amphibia y Spermatophyta contribuyeron en mayor proporción al patrón general, dado que contienen mayor cantidad de especies soporte. Sin embargo, esta contribución posiblemente está directamente relacionada a la riqueza de cada taxón evaluado, tal y como lo plantea Noguera-Urbano (2017).
Unidades biogeográficas: Encontramos que Páramo no es una sola unidad como se ha propuesto en estudios anteriores (Morrone, 2017); dichas unidades se identificaron parcialmente con las particiones de clases y divisiones; y el conjunto total de ellas (evidencia total) nos permitió identificar ocho subprovincias en el patrón general. Es importante aclarar que nuestras áreas de endemismo son unidades operacionales, que no son informativas sobre los límites reales de áreas, ya que nuestro objetivo implica la recuperación de núcleos/ patrones y no de delimitaciones altitudinales.
Nuestro patrón presenta mayor congruencia con la regionalización planteada por Jiménez-Rivillas et al. (2018), que con otras definiciones de Páramo; las regiones encontradas por las particiones de clase o división también son congruentes con los distritos propuestos por Jiménez-Rivillas et al. (2018), reconociendo la mayoría de estos a excepción del distrito Los Picachos. En general, la congruencia entre nuestro trabajo y Jiménez-Rivillas et al. (2018) nos muestra que subprovincias como Santa Marta-Perijá, Norte de Ecuador y Mérida fueron equivalentes en su extensión completa a los distritos de Jiménez-Rivillas et al. (2018) denominados como A (Venezuelan Páramo), D (Northern Ecuadorian Páramo) y F (Sierra Nevada). Otras subprovincias correspondieron a conjuntos de distritos, por ejemplo, la subprovincia Cordillera Central-Occidental fue equivalente a la suma de distritos G (Santa Inés-Sonsón), H (Paramillo del Sinú), I (Páramo de Los Farallones de Cali) y K (Páramos Quindío); de la misma forma que la subprovincia Centro-Sur de Ecuador que correspondió a la unión de los distritos B (Southern Ecuadorian Páramo) y C (Central Ecuadorian Páramo). El distrito J (Páramos de la Cordillera Oriental), en nuestro análisis lo encontramos como dos unidades independientes (Santanderes-Boyacá y Cundinamarca). En nuestro análisis el norte de Perú fue parte de la subprovincia Centro-Sur de Ecuador, la cual fue compatible con el distrito B. Finalmente, el distrito E (Los Picachos) nunca fue identificado bajo ninguna partición.
Tal similitud muestra que pese a las diferencias metodológicas (PAE en Jiménez-Rivillas et al., 2018, AE: en el presente estudio), ambas aproximaciones, independientemente, generan resultados equivalentes. Sin embargo, nuestro estudio tiene una ventaja sobre Jiménez-Rivillas et al. (2018) ya que la optimización de áreas de endemismo bajo el método de optimalidad (AE), puede ser más eficiente en la recuperación de unidades biogeográficas (Casagranda et al., 2009; Escalante, 2015). Esto, posiblemente, podría ser una de las explicaciones de que la similitud entre ambos trabajos solo fuera de un 66 %, a pesar de usar datos equivalentes de clases taxonómicas.
Es contraintuitivo que Morrone (1994), quien plantea el método de PAE en biogeografía y genera la regionalización del Neotrópico (Morrone, 2001, Morrone,2014, Morrone,2017), no logre reconocer las áreas identificadas en Jiménez-Rivillas et al. (2018) o en el presente estudio. Esto implica que es necesario hacer una revisión de la regionalización del Neotrópico. Un ejemplo de ello se puede encontrar en el estudio realizado por Romero-Alarcon (2016), quien evalúa, usando el método AE, la provincia del Chocó biogeográfico propuesta por Morrone (2014), encontrando que Chocó biogeográfico no corresponde a una única región como lo plantea Morrone (2014) sino que corresponde a tres regiones distintas, similares a las propuestas por Hernández-Camacho, Hurtado, Ortiz, & Walschburger (1992).
Cuando hablamos de Páramo en términos biogeográficos debemos referirnos a ocho subprovincias que no solo involucran las zonas de alta montaña de Colombia, Venezuela y Ecuador, sino que también deben incluir a Talamanca en Costa Rica. Esta última área ha sido considerada como Páramo en otras definiciones (Cleef & Chaverri, 1992; Luteyn, 1999; Luteyn & Balslev, 1992), basadas en su afinidad taxonómica con otros páramos colombianos. Por ejemplo, la presencia de algunos géneros de plantas (Myrrhidendron, Pterichis, Chaetolepis, entre otros) distribuidos en las regiones de alta montaña de Talamanca y los páramos en la Sierra Nevada de Santa Marta (Cleef & Chaverri, 1992). Esto mismo sucede con el norte de Perú, aunque algunos autores prefieren definirla como otra región denominada como Jalca, dada su estructura florística (Brack & Mendiola, 2000; Cabrera & Willink, 1980; de Mera et al., 2017; Van der Hammen, 1974), en nuestro caso, la encontramos como parte de la subprovincia Centro-Sur de Ecuador.
Nuestros resultados no identifican la sectorización tipo Subpáramo, Páramo y Superpáramo propuesta por Cuatrecasas (1958), ya que AE no permite identificar límites altitudinales, ni utilizar datos como comunidades de plantas o factores abióticos como el clima. Así mismo, nuestros resultados tampoco son comparables con la delimitación propuesta por Morales-Rivas et al. (2007), la cual se basa en criterios biofísicos, socioculturales e integridad ecológica. Podemos afirmar que nuestro trabajo permite observar el patrón general de unidades históricas de las zonas que han sido consideradas como Páramo, las cuales podrían ser la base para la resolución de preguntas en diferentes áreas como biogeografía histórica de patrón y eventos, diversificación, macroecología y conservación.
En conclusión, independientemente del tamaño de cuadrícula, AE permite identificar patrones generales consistentes. Adicionalmente, el uso de cuadrículas pequeñas o proporcionales al área de estudio, nos permite encontrar resultados más ajustados y de alta resolución. Páramo se ha considerado como diferentes áreas dadas algunas características como la altitud o el tipo de vegetación. En biogeografía estas áreas se han planteado como una sola unidad, mientras que nuestro trabajo define Páramo como ocho subprovincias biogeográficas las cuales podrían ser consideradas como la base para futuros análisis.
Recomendamos revisar las regionalizaciones del Neotrópico a escalas más pequeñas de región o provincia; además de explorar tamaños de cuadrícula más pequeños dentro de cada subprovincia, e involucrar métodos que incluyan directamente la elevación, de tal forma que se puedan plantear límites altitudinales de estas regiones de alta montaña.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.
Apéndice 1
