











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
En Costa Rica, el banano (Musa AAA) constituye en términos de divisas, el primer producto de exportación dentro del sector agrícola (Alpízar, 2017). En el 2017, el área bananera estaba compuesta por 42 857 hectáreas, mientras que la productividad nacional alcanzó 54.2 toneladas métricas. El manejo agronómico actual del cultivo ha permitido obtener un rendimiento alto, sin embargo, se ha observado un incremento en plagas como el picudo negro Cosmopolites sordidus (Gemar), (Coleoptera: Dryophthoridae). Este aumento podría estar relacionado al establecimiento de coberturas naturales en los cultivos, que si bien reducen la escorrentía superficial, favorecen el reciclaje de nutrientes en el suelo y el desarrollo radical, benefician la presencia de la plaga ya que encuentra un nicho más adecuado para su persistencia (CORBANA, 2011). Otro factor es la aplicación en Costa Rica de un nuevo nematicida denominado Verango (Bayer®), catalogado como banda verde debido a su bajo impacto ambiental, y que, contrario a insecticidas-nematicidas como terbufos y oxamil utilizados anteriormente, (Gutiérrez, 2014; CORBANA, 2017) no presenta ningún efecto sobre el picudo.
El picudo negro afecta el establecimiento y la producción del cultivo de banano (Rukazambuga, Gold, & Gowen, 1998; Gold, Pena, & Karamura, 2001); las hembras realizan cavidades en el cormo donde depositan los huevos, y una vez que eclosionan, las larvas continúan la perforación alimentándose del tejido vegetal (Cuillé, 1950), dañando el sistema radical, reduciendo la absorción de agua y nutrientes y aumentando la susceptibilidad de la planta a la infección por microorganismos (Rukazambuga et al., 1998; Gold et al., 2001). Dentro de los efectos de este insecto en el cultivo, se encuentra la reducción en el desarrollo de las yemas vegetativas acortando la vida de la planta, la pérdida de racimos (Cerda, López, Sanoja, Sánchez, & Jaffé, 1996), así como la maduración tardía y reducción del peso de las vainas (Carballo, 2001; Rukazambuga, Gold, Gowen, & Ragama, 2002). En casos severos, el insecto puede provocar pérdidas significativas en la plantación (Gómez & Soto, 2008).
Debido a las graves consecuencias de esta plaga, se han estudiado diferentes alternativas para su control. Los métodos utilizados para contrarrestar el picudo en plantaciones de banano se basan comúnmente en el control químico, que consiste en la aplicación de los plaguicidas organofosforados, carbamatos y piretroides (Gold et al., 2001). Una posible herramienta para el manejo de C. sordidus es el control biológico (Gold & Messiaen, 2000; Lopes, Laumann, Moore, Oliveira, & Faria, 2014), mediante la aplicación de los nemátodos entomopatógenos (NEPs) Heterorhabditis y Steinernema (Amador et al., 2015) y los hongos Cordyceps bassiana, Metarhizium anisopliae y Beauveria bassiana, (Maharaj & Khan, 2016; Gonzálezet al., 2018). La implementación de estos métodos de control en el sistema del cultivo, reduciría el uso de agroquímicos y los riesgos ambientales causados por estos.
Los NEPs de las familias Steinernematidae y Heterorhabditidae poseen una relación mutualista con bacterias de los géneros Xenorhabdus y Photorhabdus, respectivamente. Este complejo nemátodo-bacteria funciona para el control biológico de plagas, ya que son parásitos obligados de insectos que habitan el suelo, lo cual les confiere el potencial como agentes controladores de una variedad de especies de importancia económica (Stock, 1995; Shapiro-Ilan, Gouge, Piggott, & Fife, 2006; Marianelli et al., 2018).
La diversidad de cepas aisladas presentan un amplio rango de hospederos. La capacidad de los NEPs para eliminar a los insectos plaga en 24-48 h y el desarrollo de formulaciones comerciales (Kaya, 1990; Hiltpold, 2015), los convierten en una alternativa para el control de C. sordidus en el cultivo del banano. Al respecto, se han utilizado in vitro ambos géneros y especies como H. bacteriophora, S. fetilae y H. atacamensis. (Mwaitulo, Haukeland, Sæthre, Laudisoit, & Maerere, 2011; Padilla-Cubas, Carnero-Hernández, & Garcia del Pino, 2010; Amador et al., 2015); lo que demuestra el potencial de estos organismos en el control del picudo.
Sin embargo, debido a que los NEPs pueden ser afectados por factores y condiciones ambientales, tales como temperatura, radiación solar (rayos UV) y características propias del suelo como acidez, materia orgánica y textura (Bedding, 1990; Andaló et al., 2018), se requiere de estrategias que permitan su sobrevivencia en el campo, como el uso de distintos métodos de encapsulamiento (Peters, 2016; Bogantes, Flores, Castellón, & Uribe, 2018) o el aislamiento de cepas adaptadas a las condiciones del agroecosistema.
Así, el aislamiento de los NEPs a partir de suelos bananeros y el estudio de los factores que influyen en su presencia y persistencia, permitiría realizar aplicaciones en el mismo hábitat de donde fueron extraídos o en sitios que presentan condiciones ambientales similares, con el propósito de favorecer el establecimiento de estos organismos.
El objetivo de este trabajo fue aislar NEPs a partir de muestras de suelo de fincas bananeras de la región Caribe y caracterizar su hábitat para predecir presencia y ausencia de éstos según características físico-químicas y microbiológicas del suelo; y a la vez, obtener un modelo a partir de la función discriminante cuadrática que explique su ocurrencia mediante la caracterización de los hábitats seleccionados.
Materiales y métodos
Muestreo de suelos: Se tomaron muestras de suelo de nueve fincas bananeras comerciales de la región Caribe, de las provincias de Limón y Heredia (Fig. 1). Dentro de las fincas estudiadas, se seleccionaron dos hábitats predominantes según el tipo de vegetación: plantación y zona aledaña (parche boscoso, pastizal o terreno sin cultivar cubierto de maleza espesa). En cada hábitat, se eligieron tres puntos de muestreo, separados por una distancia mínima de 100 m entre sí. En cada punto se tomó una muestra de aproximadamente 1 kg de suelo, compuesta de tres submuestras recolectadas en puntos cercanos, a una profundidad de 10 a 20 cm, eliminando la capa superficial, generando un total de 54 muestras. Se seleccionaron sitios que presentaban condiciones de humedad a capacidad de campo, con sombra, presencia de hojarasca y abundancia de artrópodos según lo recomendado por Stock, Pryor y Kaya (1999). Los muestreos se llevaron a cabo entre agosto y abril.
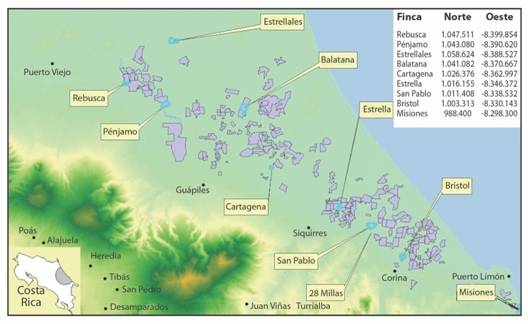
Fig. 1 Fig. 1 Distribución regional de las fincas bananeras muestreadas. Recuperado y modificado de CORBANA (2011). Regional distribution of sampled banana farms. Recovered and modified from CORBANA (2011).
Las muestras se homogenizaron, se dividieron en dos partes y se transportaron en hieleras a la Corporación Bananera Nacional (CORBANA) para realizar los análisis físico-químicos; y al Centro de Investigaciones Agronómicas (CIA) de la Universidad de Costa Rica (UCR), para el aislamiento de nemátodos y análisis microbiológicos. Los análisis y el aislamiento de los nemátodos se realizó en un período menor a 24 h de la recolecta.
Aislamiento de NEPs: Se empleó la técnica de aislamiento mediante el uso de insecto cebo (Bedding & Akhusrt, 1975) donde se utilizaron larvas de Galleria mellonella (Lepidóptera: Pyralidae), de aproximadamente 1.5 cm de longitud, o del coleóptero Tenebrio mollitor (Coleóptero: Tenebrionidae) procedentes del pie de cría establecido en el CIA (Rodríguez, Torres, Uribe, & Flores, 2009). Las larvas se colocaron en recipientes con 500 g de suelo de cada punto de muestreo. Se extrajeron aquellas que murieron en el proceso y en las que se observó signos de infección típica por NEPs: En el caso de la infección por nemátodos del género Steinernema, los cadáveres presentaron una coloración ocre a marrón, y del género Heterorhabditis, una coloración rojiza. En ambos procesos, los insectos no mostraron signos de putrefacción (Poinar, Thomas, Haygood, & Nealson, 1980). Los cadáveres se colocaron en trampas White modificada (Kaya & Stock, 1997), y aproximadamente siete días después, emergieron los NEPs, los cuales se recolectaron, se lavaron tres veces con agua destilada y se almacenaron en botellas de cultivo celular a temperatura ambiente.
A partir de cada aislamiento, se realizó el procedimiento de cría, inoculando con 1 mL de la suspensión de NEPs tres larvas de G. mellonella o T. mollitor colocadas sobre un papel filtro Whatman 1 con porosidad de 11 μm contenido en una placa Petri. Después de tres días, los cadáveres se colocaron en trampas White y los NEPs recolectados se almacenaron en solución Ringer en botellas de medio de cultivo celular. Se corroboró el género mediante microscopía de luz a una magnificación de 1 000 X y claves de identificación (Nguyen, 2007).
Análisis fisico-químico y microbiológico del suelo: Se analizó acidez, contenido de macro y micro elementos, pH, acidez intercambiable y contenido de materia orgánica (Nelson & Sommers, 1982; Thomas, 1982; Mehlich, 1984; Horwitz, 2000). Las variables Ca, Mg, y K se analizaron en cmol+/L y las variables P, Fe, Cu, Zn y Mn en mg/L y materia orgánica (MO) de manera porcentual. El análisis textural se realizó mediante la medición del peso en gramos de suelo en una suspensión a 20 °C con un hidrómetro de Bouyoucos y el nombre textural se asignó de acuerdo con el Triángulo de Texturas (Bouyoucos, 1962). Se determinó la respiración microbiana (RM) utilizando la metodología de Jenkinson y Powlson (1976) y la biomasa microbiana de carbono (CBM) mediante el método de fumigación extracción (Vance, Brookes, & Jenkinson, 1987).
Análisis estadísticos y modelo de presencia y ausencia: Se utilizó el programa SAS versión 8.0 (SAS Institute, Inc., Cary, N. C.) para realizar un análisis de varianza con el propósito de determinar diferencias entre fincas y sitios de muestreo. Se descartaron dos muestras que estadísticamente presentaron valores extremos de las mediciones para un total de 52. Se llevó a cabo una comparación mediante una prueba de T entre muestras con presencia y ausencia de NEPs para determinar la influencia de cada variable en ambas categorías, y posteriormente, un análisis de componentes principales (ACP) para encontrar qué valores de éstas explicaban la presencia y sintetizar la información obtenida reduciendo el número de datos (Shlens, 2014). Finalmente, se efectuó un análisis discriminante para determinar la eficiencia de las variables seleccionadas en el diagnóstico de ocurrencia NEPs y determinar la homogeneidad de la covarianza de ambos grupos, presencia y ausencia.
Resultados
De las 52 muestras evaluadas, se aislaron NEPs a partir de 20 (38.4 % del total); de éstas, 11 (55.0 %) se tomaron en las zonas aledañas a las plantaciones bananeras y nueve propiamente en ellas (45.0 %). Del total de muestras positivas, únicamente en una (5.0 %) proveniente de un parche boscoso de la finca Balatana (Cariari de Pococí, Limón), se comprobó la presencia del género Heterorhabditis; mientras que en las restantes 19 (95.0 %) de Steinernema. En todas las fincas con excepción de Misiones (cantón central Limón, Limón) y Bristol (Matina, Limón), se identificó la presencia de NEPs en al menos una muestra, presentando las fincas San Pablo (Siquirres, Limón), La Estrella (Siquirres, Limón) y Cartagena (Guácimo; Limón), el mayor porcentaje de puntos positivos (cuatro de seis). Por otra parte, no fue evidente que el tipo de sistema (plantación o zona aledaña), tuviese alguna relación en cuanto al número de muestras positivas (P = 0.4296).
La mayoría de las características físico-químicas y microbiológicas del suelo difirieron significativamente entre fincas y sitios de muestreo (P < 0.05). Únicamente el Zn y el K no presentaron diferencias significativas entre fincas (P = 0.5162 y P = 0.1148, respectivamente). Por otra parte, la MO (P = 0.3571), la arcilla (P = 0.2236) y la RM (P = 0.9169) no fueron significativamente distintas entre sitios de muestreo (Tabla 1).
TABLA 1 TABLE 1 Significancia estadística obtenida a partir del análisis de varianza, de la comparación de variables entre fincas y hábitats Statistical significance obtained from the analysis of variance, from the comparison of variables between farms and habitats
| Variable | Pr> F | |
| Finca | Hábitat | |
| MO | 0.0001 | 0.3571 |
| pH | > 0.0001 | 0.0102 |
| Arena | > 0.0001 | 0.0116 |
| Arcilla | 0.0001 | 0.2236 |
| Limo | 0.0027 | 0.0028 |
| BMC | 0.0003 | 0.0496 |
| RM | 0.0064 | 0.9169 |
| Acidez | 0.0003 | 0.0010 |
| Ca | > 0.0001 | 0.0005 |
| Mg | > 0.0001 | 0.0066 |
| K | 0.1148 | 0.0004 |
| P | > 0.0007 | > 0.0001 |
| Fe | > 0.0007 | 0.0002 |
| Cu | > 0.0001 | 0.0411 |
| Zn | 0.5162 | 0.1240 |
| Mn | > 0.0001 | 0.0137 |
Mediante la prueba de T se determinó que las variables MO, pH, CBM, RM y Mn, presentaron diferencias estadísticas significativas (P < 0.05) entre muestras con presencia y ausencia de NEPs. Con base en los análisis de la prueba de T, se seleccionaron las variables MO, pH, CBM, RM y acidez para realizar el ACP. Debido a que MO presentó valores elevados en comparación con los obtenidos para las demás variables, se utilizaron valores normalizados (Shlens, 2014). Según los resultados obtenidos, se observó que los dos primeros componentes principales explican un 40 y 23 % de la varianza total, respectivamente, para un total de 64 %; además presentaron “eigenvalores” superiores a uno (Tabla 2).
TABLA 2 TABLE 2 Eigenvalores obtenidos a partir de la matriz de correlación para cada uno de los componentes principales y valores de diferencia, proporción y acumulativos Eigenvalues obtained from the correlation matrix for each of the main components and values of difference, proportion and cumulative
| Componentes | Eigenvalores | Diferencia | Proporción | Acumulativo |
| 1 | 2.0258 | 0.8476 | 0.4052 | 0.4052 |
| 2 | 1.1781 | 0.3211 | 0.2356 | 0.6408 |
| 3 | 0.8570 | 0.3128 | 0.1714 | 0.8122 |
| 4 | 0.5441 | 0.1492 | 0.1088 | 0.9210 |
| 5 | 0.3948 | - | 0.0790 | 1.0000 |
El primer componente principal (CP1) presentó una correlación fuerte con MO y acidez, mostrando valores de 0.5243 y 0.4976 respectivamente y una correlación negativa con pH (-0.5706); para CBM y RM se obtuvo una correlación positiva baja de 0.0400 y 0.3867. En el caso del componente principal dos (CP2), se observó una correlación positiva alta con CBM, para la cual se obtuvo 0.6900, y RM, con una correlación de 0.4705. Así mismo, la acidez presentó una correlación negativa (-0.4607) mientras que MO y pH mostraron una correlación positiva baja con valores de 0.2291 y 0.1860. (Tabla 3).
TABLA 3 TABLE 3 Eigenvectores obtenidos mediante el ACP y correlaciones (C) de las variables originales con los componentes Eigenvectors obtained by the ACP and correlations (C) of the original variables with the components
| Variable | CP1 | CP2 | CP3 | CP4 | |||||
| E.V | C | E.V | C | E.V | C | E.V | C | ||
| MO | 0.5243 | -0.7500 | 0.2291 | 0.2500 | 0.2293 | -0.2100 | 0.7822 | 0.5800 | |
| pH | -0.5706 | 0.8100 | 0.1860 | 0.2000 | 0.3063 | -0.2800 | 0.1548 | 0.1100 | |
| CBM | 0.0481 | -0.0700 | 0.6922 | 0.7500 | -0.6987 | 0.6500 | -0.0493 | -0.0400 | |
| RM | 0.3867 | -0.5500 | 0.4705 | 0.5100 | 0.5501 | -0.5100 | -0.5666 | -0.4200 | |
| Acidez | 0.4976 | -0.7100 | -0.4607 | -0.5000 | -0.2502 | 0.2300 | -0.2014 | -0.1500 | |
Según las correlaciones originales obtenidas de las variables para cada componente principal, se obtuvo que MO, CBM, RM y acidez, poseen una correlación negativa en el componente principal uno, con valores de -0.7500, -0.0700, -0.5500 y -0.7100 respectivamente; mientras que el pH presentó una correlación positiva con un valor de 0.8100. La variable CBM tuvo un valor bajo de correlación. Para el caso del CP2, se observó una correlación negativa de -0.5000 en la acidez, mientras que el resto de las variables correlacionaron positivamente, siendo CBM la variable con un valor alto de 0.7500 (Tabla 3).
En el gráfico de dos dimensiones (Fig. 2) del CP1 y CP2, se observó que nueve muestras con presencia se ubicaron en el cuadrante cuatro, mientras que 15 con ausencia en el dos. En los cuadrantes restantes se distribuyó aproximadamente la misma proporción de muestras de ambas categorías. Estos resultados indican que las variables elegidas son más eficientes para determinar la ausencia de NEPs, es decir, de las 15 muestras ubicadas en el cuadrante cuatro, nueve correspondieron a muestras con presencia, mientras que en el cuadrante dos, de las 16 muestras observadas, 15 correspondieron a muestras con ausencia.
Como se observa en la figura dos, la mayoría de muestras con NEPs presentaron valores de RM, CBM y MO mayores a sus promedios respectivos, por el contrario, en las muestras con ausencia se observaron valores bajos. Por otro lado, tanto las muestras “1” como las muestras “0” se distribuyeron a lo largo del rango de variación de acidez y pH. Asimismo, de un total de 21 muestras evaluadas, el número de muestras “1” con valores de RM, CBM y MO superiores a sus respectivos promedios fue de 14, 16 y 16, respectivamente.
Dentro de las variables que se relacionan con presencia de NEPs se observa que MO, CBM y RM fueron las más influyentes. Valores mayores al promedio de MO y RM se asocian con presencia; por lo tanto, valores bajos de estas variables indican una disminución de los organismos, así como un pH con un valor mayor a 6.35 (Fig. 2). Por otro lado, la acidez del suelo presentó poca influencia con respecto a presencia y ausencia. De los resultados obtenidos se puede concluir que las muestras con valores altos de MO y RM, tenían la probabilidad de presentar NEPs, contrario a aquellas con valores más altos al promedio de pH evaluado (6.6).
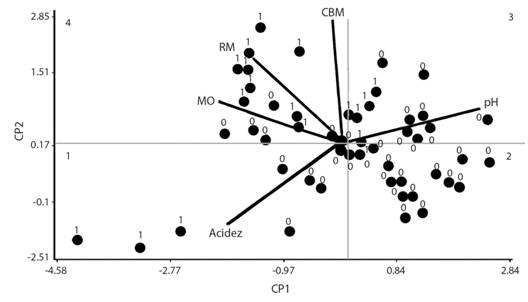
Fig. 2 Fig. 2 Bioplot de los componentes principales uno y dos, que indica si las muestras presentan (1) o no (0) NEPs. Bioplot of the main components one and two, which indicates whether the samples present (1) or not (0) EPN.
Mediante el análisis discriminante se encontró que las matrices de covarianza de ambos grupos no eran homogéneas (P < 0.0001), por lo que se utilizó una función discriminante cuadrática. Con base en esto, se determinó que un 81.2 % de la totalidad de las muestras sin NEPs (32 muestras), seis deberían pertenecer a la categoría presencia, mientras que, de las muestras positivas, un 55.0 % (nueve de 20) deberían presentar ausencia. Por lo tanto, las variables seleccionadas son más eficientes para determinar ausencia de nemátodos que presencia (Tabla 4).
TABLA 4 TABLE 4 Número y porcentaje de muestras clasificadas por la ecuación discriminante cuadrática en los grupos de presencia y ausencia de NEPs Number and percentage of samples classified by the quadratic discriminant equation in the presence and absence groups of EPN
| Ecuación cuadrática | Clasificación según ecuación | Total | |||
| 0 | 1 | ||||
| Datos observados vs. datos predichos por la ecuación discriminante | 0 | 26 (81.2 %) | 6 (18.8 %) | 32(100 %) | |
| 1 | 9 (45.0 %) | 11(55.0 %) | 20 (100 %) | ||
| Total | 35(67.3 %) | 17(67.3 %) | 52(100 %) | ||
Discusión
Los resultados obtenidos no concuerdan con lo expuesto por Mráček, Bečvář, Kindlmann y Jersáková (2005) y Lawrence, Hoy y Grewal (2006), quienes encontraron una mayor abundancia de NEPs en bordes y en bosques, que en plantaciones, debido a que estos suelos son generalmente más ricos en nutrientes y presentan una diversidad de organismos importante.
En este estudio, se observó que no hay diferencia con respecto a la presencia de NEPs entre zona aledaña y la plantación; es decir, que un ambiente alterado como las plantaciones dentro de las cuales se aplican agroquímicos, no fue en este caso, un factor determinante. Cabe destacar que los ambientes seleccionados son cercanos entre sí, y diversos estudios realizados en Costa Rica han demostrado la presencia de agroquímicos en sitios cercanos a plantaciones (Castillo, Ruepert, & Solis, 2000; Shunthirasingham et al., 2011).
Sobre este tema, Méndez, Castillo, Ruepert, Hungerbuehler y Ng (2018) estimaron las concentraciones químicas de plaguicidas en diversos ambientes como agua, aire, suelo, sedimentos y plantas a lo largo del tiempo, tomando en cuenta aspectos como propiedades de los agroquímicos, patrones de emisión, y condiciones ambientales, y encontraron altas concentraciones en agua, aire y sedimento, los cuales son medio de transporte para residuos químicos.
Se han llevado a cabo diversas investigaciones acerca el efecto de plaguicidas sobre los NEPs (De Nardo & Grewal, 2003; Laznik, Vidrih, & Trdan, 2012). Navarro, McMullen y Stock (2014) evaluaron en condiciones in vitro el efecto de los ingredientes activos presentes en los insecticidas dinotefuran [ScorpionTM 35SL] (Gowan, Yuma, AZ), indozcarb [Avaunt®] (Dupont, Wilmington, DE) e imidacloprid [Merit® 75 WP] (Bayer, NC), éste último utilizado en Costa Rica (Unidad de Registro de Agroquímicos y Equipos de Aplicación, 2019), sobre H. sonorensis y S. riobrave y determinaron que ninguna de las concentraciones analizadas: imidacloprid y dinotefuran (1, 2, 4, 8, 12 g ingrediente activo/ha) y indoxacarb (1, 2.5, 25, 250, 2 500 g ingrediente activo/ha) afectó significativamente la sobrevivencia, sugiriendo de esta manera que es posible aplicar NEPS en combinación con insecticidas (Laznik et al., 2012).
Los porcentajes de arcilla y arena de los suelos muestreados se encontraron dentro de los valores reportados para suelos bananeros costarricenses, con mínimos de 6 % y máximos de 71 % de arcilla, y 9 % y 82 % respectivamente de arena (Sandoval & Mata, 2014), estos porcentajes no afectan la sobrevivencia y movilidad del nemátodo. En el presente estudio, se observó que la textura del suelo no fue un factor determinante en la presencia de NEPs, al encontrarse todos los puntos de muestreo dentro del rango adecuado (Portillo-Aguilar, Villani, Tauber, Tauber, & Nyrop, 1999; Koppenhöfer & Fuzy, 2006). Autores como Kung (1990) mencionan que los IJ del género Steinernema dependen de sus reservas energéticas para llevar a cabo actividades como la sobrevivencia, patogenicidad y movilidad, y ésta puede ser afectada por la textura del suelo.
Los factores más importantes que determinaron la presencia de NEPs fueron MO y CBM; valores altos de MO y CBM indican condiciones más favorables para la presencia de organismos en el suelo. En el caso de los nemátodos bacteriófagos, se ha demostrado que se conglomeran debido a la atracción que presentan por la biomasa microbiana (Young, Griffiths, Robertson, & McNicol, 1998); es decir, las adiciones de materia orgánica en cultivos pueden inducir cambios con respecto a las comunidades de nemátodos. Rossi y Quénéherve (1998) encontraron que el enriquecimiento de materia orgánica en plantaciones bananeras afectó positivamente la estructura de la comunidad de nemátodos del suelo y de otros microorganismos.
Los resultados concuerdan con lo encontrado por Briar, Grewal, Somasekhar, Stinner y Miller (2007) quienes relacionan un alto contenido de materia orgánica con la presencia de nemátodos bacteriófagos y de vida libre en plantaciones de maíz, soya, avena y heno, en las cuales algunas parcelas se encontraban en transición de manejo convencional a orgánico, o bien se trabajaban con algún tipo de cobertura del suelo o insumos orgánicos. Los autores demostraron que la abundancia de nemátodos de vida libre representados en su mayoría por bacteriófagos, era mayor cuando existía un contenido alto de materia orgánica (3 %), pues su aporte al suelo en forma de abono, residuos animales o plantas de cobertura, aumentaba la disponibilidad de energía para la microbiota del suelo incrementando la actividad. Además, se ha demostrado que la abundancia de nemátodos bacteriófagos se puede atribuir directamente a la biomasa microbiana (Ferris, Venette, & Lau, 1996).
Powers et al. (2009) estudiaron utilizando técnicas moleculares, la diversidad de nemátodos de un bosque lluvioso tropical en Costa Rica, con el propósito de determinar la relación entre biomasa y diversidad tomando en cuenta la distribución vertical de los nemátodos y los hospederos con los que mostraban asociación. Dentro de sus resultados observaron la presencia de seis especies de NEPs, de los cuales el 50 % correspondía al género Heterorhabditis y el porcentaje restante a Steinernema, lo cual representa una diversidad importante de nemátodos de este tipo.
En este estudio, se observó que, de 20 muestras positivas, sólo en una se determinó la presencia del género Heterorhabditis, lo cual concuerda con lo descrito por otros autores que aislaron en mayor porcentaje (aproximadamente 80 %) a nemátodos del género Steinernema, (García & Palomo, 1996; Seenivasan, Prabhu, Makesh, & Sivakumar, 2012; Zepeda-Jazo, Molina-Ochoa, Lezama-Gutiérrez, Skoda, & Foster, 2014). Esto sugiere que la capacidad para adaptarse a un mayor rango de hábitats, factores bióticos y abióticos es más amplia en Steinernema que en Heterorhabditis (Zepeda et al., 2014). Se ha observado además que especies del género Heterorhabditis son más comunes en áreas litorales y costeras relacionando su presencia a las características del suelo y las condiciones climáticas (Griffin & Downes, 1991; Hara, Gaugler, Kaya, & Lebeck, 1991; Kanga, Waeyenberge, Hauser, & Moen, 2012).
Se ha determinado que la composición química del suelo y el pH son factores que, presentando valores extremos, pueden afectar la presencia y persistencia de los NEPs. Kung (1990), demostró que a un pH 10 se redujo la sobrevivencia de Steinernema sp.; sin embargo, no hubo diferencias entre un pH de 4 a 8. Los suelos bananeros de Costa Rica presentan un promedio de 5.5 con un valor mínimo de 4.0 y máximo de 8.0 (Sandoval & Mata, 2014); los suelos de este estudio se encontraron dentro del rango 5.6 - 7.6.
El modelo de presencia o ausencia de NEPs obtenido en este estudio concuerda con lo descrito por autores que relacionan la calidad del suelo, con la abundancia de nemátodos de vida libre (Nahar et al., 2006). Cabe destacar que el modelo explica mejor la ausencia de los nemátodos; es decir, en suelos con valores bajos de respiración (< 0.20 CO2 g-1 día1), MO (< 4.7 %) y CBM (156 C Kg -1 suelo -1), existe una probabilidad del 80 % de no encontrar nemátodos de los géneros Heterorhabditis ySteinernema.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

