











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Los bosques lluviosos tropicales de América latina, poseen una gran diversidad de especies arbóreas (Gentry, 1988; Phillips, Hall, Gentry, Sawyer, & Vizquez, 1994; Valencia, Balslev, Paz, & Mino, 1994), y son considerados como los ecosistemas terrestres más biodiversos del planeta (Hartshorn, 2002; Myers, Mittermeier, Mittermeier, Da Fonseca, & Kent, 2000). Dentro de los cuales, se han registrado zonas con más de 300 especies por hectárea (Gentry, 1988; Valencia et al., 1994). Los factores que explican esta alta diversidad arbórea han sido de gran interés. Particularmente, Leigh et al. (2004), en una revisión de literatura consideraron que esta alta diversidad se encuentra explicada por factores como la limitación en dispersión, heterogeneidad de hábitats, suelos, topografía, disturbios, presión por patógenos, productividad, clima, y tasas de especiación y extinción basados en modelos neutrales y filtros ambientales. Asimismo Antonelli y Sanmartin (2011) consideraron que los factores que explican la alta diversidad de árboles en el Neotrópico se pueden clasificar en dos categorías: los factores bióticos (adaptaciones edáficas, interacciones con polinizadores, dispersores y herbívoros, nicho y limitación en dispersión), y los abióticos (tiempo, precipitación, temperatura, área geográfica, topografía e hidrología). Dichos factores explican en distintas proporciones la diversidad arbórea (Givnish, 1999).
Las condiciones edáficas han sido consideradas como un factor ambiental que explica significativamente la alta diversidad de especies arbóreas en bosques lluviosos tropicales (Huston, 1980; Gentry, 1981; Clark, Clark, & Read, 1998; Clark, 2002; Quinto & Moreno, 2014; Nadeau & Sullivan, 2015). Por ejemplo, Huston (1980, 1994) sintetizó varios trabajos que proponen que la diversidad de plantas es baja en suelos pobres en nutrientes, aumenta conforme incrementa el nivel de nutrientes, y finalmente vuelve a disminuir cuando el suelo es muy fértil. En consecuencia, se ha planteado la hipótesis de que en condiciones extremas de baja y/o alta fertilidad edáfica, la diversidad arbórea tropical tiende a disminuir (Clark, 2002; Huston, 1980, 1994, 2012).
Con el objetivo de poner a prueba dicha hipótesis, Huston (2012) estudió la relación existente entre la precipitación, la fertilidad del suelo, la productividad primaria neta y la diversidad arbórea de diferentes regiones del mundo. En los sitios con mayor precipitación, la productividad primaria neta y la fertilidad edáfica tienden a disminuir, pero la diversidad de especies es alta (Huston, 2012). Asimismo, Huston (1980) evaluó la riqueza de especies arbóreas y su relación con los nutrientes del suelo en boques lluviosos tropicales de Costa Rica y encontró que la diversidad tiende a disminuir con la fertilidad del suelo (sumatoria de los porcentajes de P, K y Ca). En suelos pobres en nutrientes (P, K y Ca) se registra la mayor riqueza de especies arbóreas (Huston, 1980; Nadeau & Sullivan, 2015). Esta tendencia en diversidad arbórea también ha sido reportada por Faber-Langendoen y Gentry (1991), Clinebell, Phillips, Gentry, Stark, y Zuuring (1995), Duque, Cavelier, y Posada (2003), y Quinto y Moreno (2014), quienes han encontrado altos valores de riqueza de especies arbóreas en suelos pobres en nutrientes de diferentes regiones tropicales. Particularmente, Clinebell et al., (1995) reportaron correlaciones negativas de la riqueza de especies arbóreas con el contenido de K, Mg, Ca y fertilidad del suelo; mientras que, con el Al y Fe la relación tiende a ser positiva. Mientras que, Condit, Engelbrecht, Pino, Pérez, y Turner (2013), evidenciaron una asociación entre la composición y riqueza de especies arbóreas y el contenido de humedad de la estación seca, el P disponible y el Ca. Sin embargo, algunos trabajos muestran que el efecto de las variables edáficas sobre la diversidad y composición florística es poco significativo (Barreto, Duque, Cárdenas, & Moreno, 2010; Duque et al., 2003; Quinto & Moreno, 2014; Valencia et al., 2004).
El Chocó biogeográfico es una región del neotrópico ubicada al noroeste de Sudamérica, considerado como un hotspot de conservación debido a su alta biodiversidad y endemismo (Myers et al., 2000). Allí se han registrado entre 100 y 300 especies arbóreas por hectárea (Faber-Langendoen & Gentry, 1991; Gentry, 1988; Quinto & Moreno, 2014). Gentry (1988) atribuyó esta inusual diversidad a los altos niveles de precipitación y ausencia de sequías, aunque este supuesto ha sido poco contrastado, pues en la región se han realizado pocos estudios.
En un estudio realizado en la localidad de Salero (Chocó biogeográfico), se encontró que la riqueza de especies arbóreas se relaciona positivamente con el Al del suelo y negativamente con la micro-topografía (Quinto & Moreno, 2014). Sin embargo, la escala de evaluación fue local, lo que posiblemente incidió en los resultados, porque no se evaluaron gradientes contrastables de condiciones edáficas. En consecuencia no se encontró evidencia que apoye la hipótesis de Huston (1980, 1994) y Clinebell et al., (1995) sobre diversidad arbórea y contenido de nutrientes del suelo en esta región. El objeto del presente estudio es evaluar como las condiciones edáficas (acidez, MO, Al, N total, P disponible, K, Mg, Ca, capacidad de intercambio catiónico, y textura) explican la diversidad y dominancia arbórea, a escala regional, empleando tres bosques pluviales tropicales ubicados en localidades independientes (Opogodó, Pacurita y Salero) en el Chocó biogeográfico. Para ello, se comparó la diversidad arbórea a nivel de unidades de muestreo (400 m2), y mediante curvas de diversidad se evaluó la acumulación de especies. Además, nos planteamos la hipótesis de que en suelos con altos contenidos de Al, y pobres en nutrientes como el N total, K, Mg, Ca, la riqueza de especies arbóreas podría ser alta. Asimismo, basados en Condit et al. (2013), se espera que el P disponible del suelo tenga una asociación positiva con la riqueza de especies arbóreas en bosques del Chocó biogeográfico.
Materiales y métodos
Área de estudio: El estudio se realizó en los bosques lluviosos tropicales de las localidades de Pacurita (municipio de Quibdó), Salero (municipio de Unión Panamericana) y Opogodó (municipio de Condoto), departamento del Chocó, Colombia. Estas localidades hacen parte de la subregión ecogeográfica Central Norte del Chocó biogeográfico que comprende la totalidad de las cuencas altas de los ríos Atrato y San Juan. Estos bosques presentan una altura de entre 70 y 150 m.s.n.m. y poseen una precipitación promedio anual de 7 500-10 000 mm (Quinto & Moreno, 2016). Los suelos son ultisoles, extremadamente ácidos, con alta saturación de Al y con muy bajas concentraciones de P, Mg, Ca y CICE. Los contenidos de K son intermedios. Mientras que la MO y el N Total son muy altos, especialmente en Opogodó. En términos de textura, los suelos de Pacurita presentan mayor contenido de arcilla, en Opogodó son arenosos, y en Salero poseen más limo (Quinto & Moreno, 2016). La composición arbórea de estas localidades está dominada por especies como: Mabea chocoensis, Pouteria sp., Oenocarpus bataua, Eschweilera pittieri, Croton jorgei, Wettinia quinaria, Eschweilera aff. integrifolia, Brosimum utile, entre otras. También están presentes las familias Sapotaceae, Euphorbiaceae, Lecythidaceae, Arecaceae, Myristicaceae, Vochysiaceae y Clusiaceae (García et al., 2003; Quinto & Moreno, 2014).
Establecimiento de parcelas: En la localidad de Salero se emplearon dos parcelas permanentes de investigación establecidas entre abril y agosto de 1998 mediante la metodología BIOTROP (García et al., 2003), las cuales hacen parte de la red de parcelas permanentes de Colombia (Vallejo-Joyas et al., 2005). Cada una de estas parcelas consiste de un cuadrado de 500 x 20 m dividido en 25 cuadrantes de 20 x 20 m, que se emplearon como unidades de muestreo de diversidad y dominancia arbórea. Por su parte, en las localidades de Opogodó y Pacurita, se emplearon parcelas de 100 x 100 m instaladas en el 2013, divididas en 25 cuadrantes de 20 x 20 m. En Opogodó, se establecieron tres parcelas de una hectárea, mientras que en Pacurita se instalaron dos parcelas de una ha de bosque. En cada una de los cuadrantes de 20 x 20 m se realizaron mediciones de los parámetros fisicoquímicos del suelo (acidez, MO, Al, N total, P disponible, K, Mg, Ca, capacidad de intercambio catiónico, y textura) que se detallan en Quinto y Moreno (2016).
Inventario de árboles en las parcelas: Se midió el diámetro a la altura del pecho (DAP) (1.3 m sobre el nivel del suelo) a todos los árboles con DAP ≥ 10 cm. A todos los árboles medidos se les determinó el hábito de crecimiento en las categorías de árbol, bejuco o liana y palma, se anotaron las observaciones particulares de cada individuo.
Identificación botánica: Se identificaron todas los individuos hasta el máximo nivel taxonómico posible (NN, especie, género, familia botánica) en el herbario de la Universidad Tecnológica del Chocó “D.L.C.” “Herbario Chocó”. Esta identificación se llevó a cabo utilizando las claves especializadas de Gentry (1993) y Mahecha (1997).
Análisis de datos: Los datos de los inventarios de los árboles de las parcelas se emplearon para determinar la abundancia de las especies, con lo cual se determinó la riqueza, diversidad y dominancia arbóreas en las parcelas y cuadrantes (Moreno, 2001). La diversidad alfa se determinó mediante los siguientes índices de diversidad: dominancia de Simpson y de Berger-Parker, equidad de Shannon-Wiener y diversidad de Margalef (Magurran, 1988; Moreno, 2001). Para hacer más robustas las evaluaciones de diversidad y dominancia arbórea, se emplearon los coeficientes o exponentes de Hill, para obtener la riqueza efectiva a partir de los índices de Shannon-Winner y Simpson (Hill, 1973; Moreno 2001, Jost, 2006). Específicamente, se determinó la serie de números de Hill (Hill, 1973; Moreno 2001), de siguiente forma:
N0 = número total de especies (S)
N1 = número de especies abundantes = eH’
N2 = número de especies muy abundantes = 1/λ
Donde H' es el índice de Shannon-Winner, λ es el índice de Simpson. Igualmente, se determinó el índice de equidad de Hill (Hill, 1973), de la siguiente forma: E' = N2/N1; donde N2 es el número de especies muy abundantes, y N1 es el número de especies abundantes (Hill, 1973; Moreno, 2001). Los valores de diversidad y dominancia arbórea de los cuadrantes se compararon mediante la prueba no paramétrica de Kruskal-Wallis, dado que no se cumplieron los supuestos de normalidad y homogeneidad de varianzas, evaluados con los estadísticos de Bartlett y Hartley (Hoshmand, 1998). Adicionalmente, se determinaron las curvas de acumulación de especies con los estimadores no paramétricos de diversidad y riqueza de especies, Jacknife 1, Chao 2 y Bootstrap (Chao, Chazdon, Colwell, & Shen, 2005; Colwell, Mao, & Chang, 2004). Estas curvas se determinaron con el programa EstimateS (Chao et al., 2005; Colwell et al., 2004, Colwell et al., 2012). La precisión de los estimadores se estimó evaluando el sesgo y exactitud por medio de la comparación de la riqueza estimada y la verdadera (López-Gómez & Williams-Linera 2006). Para este análisis se emplearon las fórmulas de Chiarucci, Enright, Perry, Miller, y Lamont (2003):
Sesgo = (Sestimada - Sverdadera) / Sverdadera
Exactitud = ((Sestimada - Sverdadera)/ Sverdadera)2
Las correlaciones entre las variables de diversidad y las condiciones edáficas se evaluaron mediante el coeficiente de correlación por rangos de Spearman y el análisis de componentes principales (ACP). Para la construcción del ACP y la matriz de correlación se tomaron los datos de diversidad, riqueza, equidad, dominancia arbórea, y los valores de las variables edáficas (acidez, MO, Al, nutrientes, CICE, y textura) registradas en cada una de las unidades de muestreo (400 m2), y se efectuaron las respectivas correlaciones.
Finalmente, se emplearon regresiones lineales múltiples para evaluar la relación conjunta existente entre la riqueza, equidad de Shannon y dominancia de Simpson con las variables edáficas. Las variables edáficas fueron transformadas con Log(+1) para reducir su anormalidad y las que fueron significativas en la regresión múltiple se seleccionaron con el método de Stepwise - Backward (gradualmente hacia atrás). En general, los análisis se realizaron con cuatro programas estadísticos, para comparar la diversidad entre unidades de muestreo se empleó el Statgraphics Centurion XV (Statistical Graphics Corp., 2002), los índices de diversidad se determinaron con el Past (Hammer, Harper, & Ryan, 2001) las curvas de acumulación de especies se generaron con el EstimateS 9 (Chao et al., 2005; Colwell et al., 2004, 2012) y los análisis de componentes principales se realizaron con el CANOCO 4.5 (ter Braak & Šmilauer, 2002).
Resultados
En los bosques del Chocó biogeográfico se registró la mayor riqueza total de especies (sp) por hectárea (ha) en la localidad de Salero, con 219; en Opogodó menor riqueza total de especies, con 75 (Tabla 1). La mayor abundancia de individuos se registró en Pacurita, con 979 árboles/ha; mientras que, en Opogodó se registró la menor abundancia con 636 árboles/ha (Tabla 1). La mayor dominancia de Simpson y de Berger-Parker se registró en Opogodó. Por su parte, la mayor equidad de Shannon-Weinner y diversidad de Margalef se registró en Salero (Tabla 1). Los números de especies abundantes de Hill (N1) mostraron un mayor valor en las localidades de Salero y Pacurita, mientras que el número de especies muy abundantes de Hill (N2) y la equidad de Hill fueron mayores en los bosques de Salero (Tabla 1).
TABLA 1 TABLE 1 Índices de diversidad alfa para las especies arbóreas de bosques tropicales del Chocó biogeográfico, Colombia Alpha diversity indices for tree species from tropical forests of the biogeographic Chocó, Colombia
| Salero | Opogodó | Pacurita | ||||||
| Índices de diversidad alfa | Parcela E | Parcela U | PI | PII | PIII | PIV | PV | |
| Riqueza de especies (esp./ha) | 219 | 179 | 90 | 75 | 78 | 97 | 93 | |
| Abundancia de individuos (ind./ha) | 712 | 644 | 955 | 708 | 636 | 913 | 979 | |
| Dominancia de Simpson (D) | 0.01 | 0.02 | 0.027 | 0.032 | 0.032 | 0.027 | 0.03 | |
| Equidad de Shannon-Weinner (H’) | 4.81 | 4.63 | 3.91 | 3.76 | 3.77 | 3.97 | 3.89 | |
| Diversidad de Margalef (I) | 33.19 | 27.52 | 12.97 | 11.28 | 11.93 | 14.09 | 13.36 | |
| Dominancia de Berger-Parker (D) | 0.05 | 0.07 | 0.057 | 0.08 | 0.073 | 0.071 | 0.077 | |
| N1 de Hill (número de especies abundantes) | 20.57 a | 16.46 b | 19.74 a | |||||
| N2 de Hill (número de especies muy abundantes) | 18.74 a | 14.10 c | 16.83 b | |||||
| Equidad de Hill | 0.90 a | 0.85 b | 0.85 b | |||||
A nivel de cuadrantes, la riqueza fue similar en Pacurita y Salero con promedios de 22.96 y 22.16 especies/cuadrante, respectivamente. Mientras que, en Opogodó fue significativamente menor con 19.1 especies/cuadrante (K 2, 172 = 23.22 P < 0.0001) (Fig. 1). Contrario a ello, la dominancia de Simpson fue significativamente mayor en Opogodó (Fig. 1). Las curvas de acumulación de especies mostraron que en Salero se alcanzó la mayor acumulación de especies, con un máximo de 301 especies en dos ha muestreadas. En Opogodó se registró un total de 119 especies en tres ha. Asimismo en Pacurita se registró un total de 113 especies arbóreas en dos ha (Fig. 2). En ambas localidades (Opogodó y Pacurita), los estimadores de diversidad, Jack 1, Chao 2 y Bootstrap, presentaron valores cercanos a los de la diversidad observada (Fig. 2). En consecuencia los valores de sesgos y exactitudes de los estimadores fueron muy bajos (Tabla 2). En Salero los estimadores presentaron valores muy altos y los sesgos de los estimadores Jacknife 1 y Chao 2 fueron igualmente altos. Aunque en esta localidad se presentó la mayor exactitud de estimación con el Chao 2 (Tabla 2).
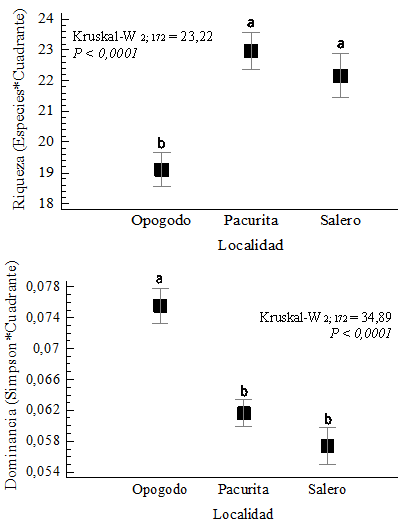
Fig. 1. Fig. 1 Riqueza y dominancia de especies arbóreas de los bosques lluviosos tropicales de las localidades de Opogodó, Pacurita y Salero en el Chocó biogeográfico, Colombia. Richness and dominance of arboreal species of the tropical rainforests of the localities of Opogodó, Pacurita and Salero in the biogeographical Chocó, Colombia.
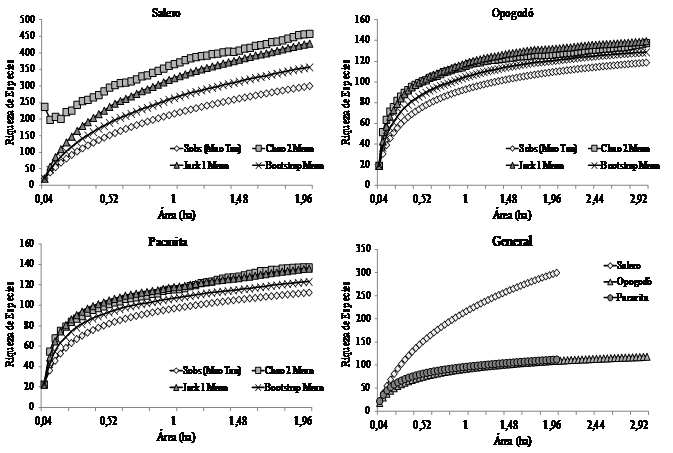
Fig. 2. Fig. 2 Curvas de acumulación de especies de arbóreas de los bosques pluviales tropicales de Opogodó, Pacurita y Salero en el Chocó biogeográfico, Colombia. Accumulation curves of tree species from the tropical rainforests of Opogodó, Pacurita and Salero in the biogeographical Chocó, Colombia.
TABLA 2 TABLE 2 Comparación de estimadores de diversidad (Jacknife 1, Chao 2, Bootstrap), sesgos y exactitudes de tres bosques tropicales del Chocó biogeográfico, Colombia Comparison of diversity estimators (Jacknife 1, Chao 2, Bootstrap), biases and accuracies of three tropical forests of the biogeographic Chocó, Colombia
| Estimadores de diversidad | Salero | Opogodó | Pacurita |
| Especies totales observadas | 301 | 119 | 113 |
| Jacknife 1 | 429.38 | 139.72 | 136.52 |
| Chao 2 | 458.45 | 137.84 | 137.59 |
| Bootstrap | 357.26 | 128.90 | 123.41 |
| Sesgo de Jacknife 1 | 0.427 | 0.174 | 0.208 |
| Exactitud de Jacknife 1 | 0.182 | 0.030 | 0.043 |
| Sesgo de Chao 2 | 0.523 | 0.158 | 0.218 |
| Exactitud de Chao 2 | 0.274 | 0.025 | 0.047 |
| Sesgo de Bootstrap | 0.187 | 0.083 | 0.092 |
| Exactitud de Bootstrap | 0.035 | 0.007 | 0.008 |
Mediante el coeficiente de correlación de Spearman se determinó que la riqueza y equidad arbórea se relacionaron de forma positiva con el P disponible, Al, limo y arcilla; mientras que, con el pH, MO, Ca, Mg, K y arena, la relación fue negativa (Tabla 3). Por su parte, las variables de dominancia se relacionaron de forma positiva con el pH, MO, Ca y arena, mientras que con el P y el Al la relación fue negativa (Tabla 3). El ACP presentó cuatro componentes significativos con Eigenvalue de 7.88 (43.8 %), 3.87 (21.53 %), 1.57 (8.7 %) y 1.15 (6.39 %) respectivamente que explican el 80.5 % de la variación. Los dos primeros componentes que explicaron el 65.3 % de la variación (Fig. 3). El ACP mostró una asociación entre la riqueza, equidad y abundancia de individuos, con el P disponible; así mismo, la dominancia mostró asociación con el Ca, K y Mg (Fig. 3).
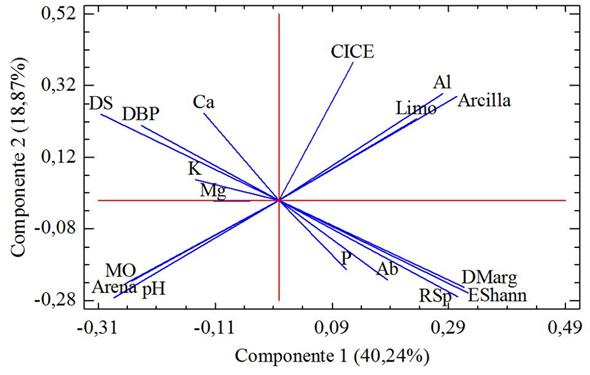
Fig. 3. Fig. 3 Principal component analysis of tree diversity and edaphic conditions (pH, Al, MO, P, K, Ca, Mg, CICE, Clay, Limo, Sand) of the rain forests of Chocó, Colombia. In the figure, DS is the Simpson dominance index, DBP is the Berger and Parker dominance index, Ab is the abundance of individuals, RSp is the species richness, DMarg is the Margalef index and EShann is the index of Shannon-Weiner equity. Análisis de componentes principales de la diversidad arbórea y condiciones edáficas (pH, Al, MO, P, K, Ca, Mg, CICE, Arcilla, Limo, Arena) de los bosques pluviales del Chocó, Colombia. En la figura, DS es el índice de dominancia de Simpson, DBP es el índice de dominancia de Berger y Parker, Ab es la abundancia de individuos, RSp es la riqueza de especies, DMarg es el índice de Margalef y EShann es el índice de equidad de Shannon- Weiner.
TABLA 3 Coeficiente de correlación por rangos de Spearman para variables de diversidad arbórea y condiciones edáficas de tres bosques lluviosos tropicales (Opogodó. Pacurita y Salero) del Chocó biogeográfico, Colombia TABLE 3 Spearman rank correlation coefficient for tree diversity variables and edaphic conditions of three tropical rainforests (Opogodó, Pacurita and Salero) of the biogeographical Chocó, Colombia
| Riqueza | Abundancia | Shannon | Margalef | Dominancia | Berger_Parker | |
| pH | -0.37** | -0.38** | -0.33** | -0.32** | 0.27** | 0.12ns |
| Materia orgánica | -0.37** | -0.29** | -0.35** | -0.35** | 0.30** | 0.13ns |
| Fósforo | 0.25** | 0.18* | 0.23** | 0.23** | -0.20** | -0.10ns |
| Aluminio | 0.36** | 0.22** | 0.38** | 0.38** | -0.36** | -0.22** |
| Calcio | -0.33** | -0.26** | -0.32** | -0.32** | 0.29** | 0.21** |
| Magnesio | -0.16* | -0.25** | -0.10ns | -0.10ns | 0.04ns | -0.05ns |
| Potasio | -0.17* | -0.14ns | -0.16* | -0.17* | 0.14ns | 0.05ns |
| CICE | 0.06ns | 0.01ns | 0.09ns | 0.08ns | -0.10ns | -0.06ns |
| Arena | -0.32** | -0.01ns | -0.39** | -0.41** | 0.40** | 0.26** |
| Limo | 0.27** | -0.02ns | 0.34** | 0.36** | -0.36** | -0.25** |
| Arcilla | 0.38** | 0.19** | 0.40** | 0.41** | -0.37** | -0.22** |
*: P < 0.05; **: P < 0.01 ns: No significativa.
Mediante la regresión lineal múltiple se determinó que la riqueza de especies y la equidad de Shannon fueron explicadas significativamente por el P disponible, Al Mg y CICE, con un porcentaje de explicación del 30.9 % (R2 = 0.31) para riqueza de especies, y un 32.7 % (R2 = 0.33) para la equidad. Además, tanto la riqueza de especies y la equidad de Shannon, presentaron una relación negativa con el CICE del suelo (Tabla 4). Por su parte, la equidad mostró también una relación significativa negativa con el porcentaje de arena (Tabla 4). Finalmente, la dominancia de Simpson presentó relación significativa negativa con la Al, Ca, Mg, y K; mientras que, con el CICE, arena y limo, la relación fue positiva, con un porcentaje de explicación del 30.4 % (R2 = 0.3; Tabla 4).
TABLA 4 TABLE 4 Regresiones lineales múltiples de las variables de diversidad arbórea y condiciones edáficas de los bosques pluviales tropicales (Opogodó. Pacurita y Salero) del Chocó biogeográfico, Colombia. El método de Stepwise - backward Multiple linear regressions of tree diversity variables and edaphic conditions of the tropical rain forests (Opogodó, Pacurita and Salero) of the biogeographical Chocó, Colombia. The Stepwise-backward method
| Riqueza de especies | ||||
|---|---|---|---|---|
| Parámetro | Estimación | Error Estándar | Estadístico T | Valor-P |
| CONSTANTE | 21.2777 | 2.47234 | 8.60632 | 0.0000 |
| LOG(Fósforo+1) | 4.65855 | 1.48164 | 3.14418 | 0.0020 |
| LOG(Aluminio+1) | 16.5579 | 2.73334 | 6.05775 | 0.0000 |
| LOG(Magnesio+1) | 12.3497 | 4.66154 | 2.64927 | 0.0089 |
| LOG(CICE+1) | -15.4497 | 3.66902 | -4.21084 | 0.0000 |
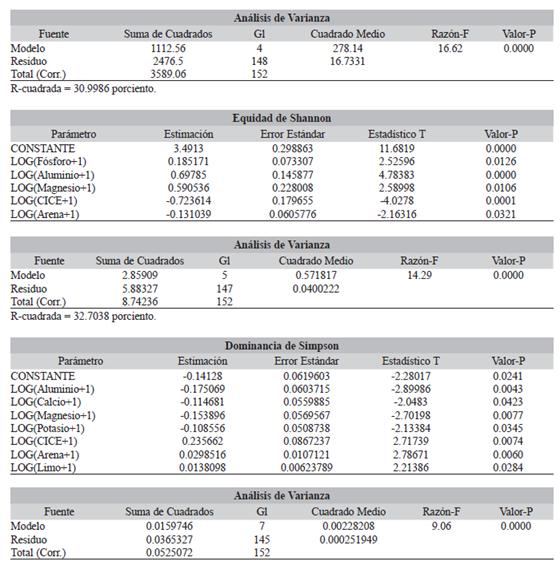
R-cuadrada = 30.42 por ciento.
Además mediante una revisión de artículo se determinó que la riqueza de especies arbóreas de bosques del Chocó pueden presentar valores muy variables (bajos y altos) de diversidad arbórea (Tabla 5). Entre los cuales se puede mencionar a Gentry (1993), Galeano (2001), Mosquera, Robledo, y Asprilla (2007), Quinto y Moreno (2014), y Álvarez-Dávila et al. (2016) que presentan valores diferentes de riqueza de especies por ha (Tabla 5). Particularmente, en este estudio en Opogodó, con suelos arenosos y ricos en N y MO, se presentó una diversidad baja; mientras que en los suelos pobres en nutrientes y de topografía quebrada de Pacurita y Salero se presentaron valores altos de riqueza de especies.
TABLA 5 TABLE 5 Diversidad y abundancia de especies arbóreas de bosques lluviosos tropicales del Chocó biogeográfico, Colombia Diversity and abundance of tree species from tropical rainforests of the biogeographical Chocó, Colombia
| Sitios | N° Individuos | N° de Especies | Margalef | Área (ha) | DAP (cm) | Referencia |
| Tutunendo 1. Colombia | 763 | 103 | 15.37 | 1.0 | ≥ 10 | Citado por Mosquera et al. (2007) |
| Tutunendo 2. Colombia | 717 | 92 | 13.84 | 1.0 | ≥ 10 | Citado por Mosquera et al. (2007) |
| Pacurita. Colombia | 158 | 66 | 12.84 | 0.3 | ≥ 10 | García et al., (2004) |
| San Martin. Colombia | 112 | 49 | 10.17 | 0.3 | ≥ 10 | García et al., (2004) |
| San José. Colombia | 298 | 124 | 21.59 | 0.3 | ≥ 10 | García et al., (2004) |
| Guadalupe. Colombia | 192 | 44 | 8.18 | 0.3 | ≥ 10 | García et al., (2004) |
| Pacurita. Colombia | 654 | 95 | 14.50 | 1.0 | ≥ 10 | Citado por Mosquera et al., (2007) |
| Coquí. Colombia | 471 | 158 | 25.51 | 0.4 | ≥ 5 | Galeano (2001) |
| Nuquí. Colombia | 414 | 151 | 24.89 | 0.4 | ≥ 5 | Galeano (2001) |
| Bajo Calima. Colombia | 675 | 258 | 39.45 | 1.0 | ≥ 10 | Faber-Langendoen y Gentry (1991) |
| Opogodó. Colombia | 955 | 90 | 12.97 | 1.0 | ≥ 10 | Este estudio |
| Opogodó. Colombia | 708 | 75 | 11.28 | 1.0 | ≥ 10 | Este estudio |
| Opogodó. Colombia | 636 | 78 | 11.93 | 1.0 | ≥ 10 | Este estudio |
| Pacurita. Colombia | 913 | 97 | 14.08 | 1.0 | ≥ 10 | Este estudio |
| Pacurita. Colombia | 979 | 93 | 13.36 | 1.0 | ≥ 10 | Este estudio |
| Salero. Colombia | 712 | 219 | 33.19 | 1.0 | ≥ 10 | Este estudio |
| Salero. Colombia | 644 | 179 | 27.52 | 1.0 | ≥ 10 | Este estudio |
Discusión
Posiblemente, la alta diversidad registrada en Salero está relacionada con la forma de las parcelas. Dado que, en Salero las parcelas fueron rectangulares (20 x 500 m); mientras que, en Opogodó y Pacurita, fueron cuadradas (100 x 100 m). El hecho de que en Salero las parcelas fueran rectangulares probablemente permitió evaluar una mayor variedad de hábitats y ambientes edáficos (Clark, 2002), lo que seguramente contribuyó a registrar una mayor riqueza de especies arbóreas; dado que, las parcelas de Salero atraviesan mayor variedad de microhábitats, como quebradas, pendientes, áreas pantanosas y tierra firme.
La influencia de la forma de las parcelas sobre la diversidad de especies arbóreas ha sido evaluada por varios investigadores previamente (Vallejo-joyas et al., 2005). Por ejemplo el efecto de la forma de la parcela sobre la riqueza de especies del bosque evaluando tres formas: rectangulares de 20 x 500 m, transectos de 10 x 1 000 m y cuadradas de 100 x 100 m, en bosques lluviosos tropicales de Pasoh (Malasia) y Barro Colorado (Panamá); y encontraron que en las parcelas rectangulares la diversidad de especies incrementa entre 5.3-11.3 % más que en las parcelas cuadradas (Condit et al., 1996); similar a lo encontrado en este estudio, al comparar las curvas de acumulación de especies entre localidades.
La alta diversidad arbórea registrada en algunos sitios del Chocó biogeográfico, corroboran la aseveración de Gentry (1988), quien postuló a los bosques de esta región como candidatos a ser uno de los más biodiversos del Neotrópico. Particularmente, la diversidad encontrada en Salero fue similar a la encontrada en sitios de la región geográfica como Tutunendo y Bajo Calima (Clinebell et al., 1995; Gentry, 1988). En este sentido, Faber-Langendoen y Gentry (1991) reportaron una riqueza de 258 especies arbóreas por hectárea en bosques del Bajo Calima, sobre suelos arcillosos, pobres en nutrientes, con muy alto contenido de Al y de topografía quebrada (Faber-Langendoen & Gentry, 1991). Similar a lo registrado en Salero, con 219 especies arbóreas por hectárea e igualmente con suelos pobres en nutrientes, con alto contenido de Al y de topografía quebrada. Aunque en el Bajo Calima, las parcelas fueron cuadradas y en Salero fueron rectangulares. Asimismo, Galeano (2001), registró 294 especies arbóreas en 1.8 ha en bosques del Golfo de Tribugá (Chocó biogeográfico) con suelos arenosos y ricos en nutrientes.
En este estudio se observó que la riqueza y equidad de especies presentan una correlación negativa con algunas variables de fertilidad del suelo, específicamente con nutrientes del suelo como Ca, Mg, K y MO. Estos resultados son similares a los reportados por Huston (1980) quien evidenció una relación negativa entre la disponibilidad de nutrientes (P, K, Ca, Na, bases totales y CICE) del suelo y la riqueza de especies en bosques lluviosos de Costa Rica. Asimismo, Clinebell et al., (1995) reportó que la riqueza se relaciona negativamente con la disponibilidad de nutrientes (K, Ca, Mg y P) en un meta-análisis a nivel de bosques Neotropicales. Basados en lo anterior, Huston (1980, 2012) consideró que la mayor riqueza ocurre bajo condiciones que restringen el crecimiento arbóreo.
Por otra parte, la relación entre algunos nutrientes como P disponible y Mg con la diversidad y equidad arbórea fue positiva, estas fueron las variables edáficas que mejor explicaron la riqueza y equidad. Esto se relaciona con lo encontrado por Condit et al., (2013) que evidenciaron una asociación entre la composición y riqueza de especies arbóreas y el contenido de humedad de la estación seca, el P disponible y el Ca. Al parecer, bajo condiciones de baja fertilidad edáfica, la concentración de P disponible puede tener un efecto significativo y positivo sobre la diversidad arbórea.
Huston (1980) y Clinebell et al., (1995) encontraron que la riqueza de especies presenta correlaciones positivas con la precipitación, pero negativa con la fertilidad del suelo. Así mismo, la fertilidad del suelo presenta correlación negativa con la precipitación (Clinebell et al., 1995; Huston, 1980), lo cual es consistente con que en condiciones de alta precipitación y baja fertilidad edáfica la riqueza de especies tiende a ser mayor (Huston, 2012). Sin embargo, Gentry (1982, 1988) y Clinebell et al. (1995) consideraron que la precipitación anual y los meses de sequias son las variables que mejor explican la riqueza de especies y no las variables edáficas como disponibilidad de nutrientes, contrario a lo planteado por Huston (1980, 2012). Particularmente, los estudios realizados en bosques del Chocó muestran que las condiciones edáficas explican poco la diversidad arbórea, como se evidenció en Quinto y Moreno (2014) y en el presente estudio; los nutrientes aunque muestran correlaciones significativas con las variables de diversidad arbórea su porcentaje de explicación es muy bajo (< 40 %). La posible razón de este bajo porcentaje de explicación puede estar relacionada con la influencia de otras variables ambientales y biológicas como la limitación en dispersión, clima, entre otros (Duque et al., 2003; Valencia et al., 2004), que al parecer están teniendo mayor influencia sobre la diversidad arbórea.
Aunque una correlación no necesariamente representa causalidad, la pregunta que surge es: ¿por qué bajo condiciones de alta precipitación y baja disponibilidad de nutrientes, la riqueza de especies arbóreas tiende a ser alta en bosques tropicales? Según Huston (1980) la alta precipitación genera reducción en los niveles de nutrientes del suelo por lixiviación (Austin & Vitousek, 1998; Quinto & Moreno, 2016; Santiago, Schuur, & Silvera, 2005); lo cual, produce una mayor limitación del crecimiento arbóreo (Lambers, Chapin III, Pons, & Thijs, 2008; Marschner, 1991; Salisbury & Ross, 1994), pero conduce a una mayor abundancia de árboles pequeños. La alta precipitación no solo reduce la cantidad de nutrientes por lixiviación, sino que también produce mayor acidez y concentración edáfica de Al (Jenny, 1941; Austin & Vitousek, 1998). Por consiguiente, con alta precipitación tienden a presentarse altos contenidos de Al edáfico que reducen las tasas de crecimiento arbóreo (Marschner, 1991) y ocasiona una mayor riqueza de especies (Huston, 1980). Esto explicaría porque se encontró una relación positiva entre la riqueza de especies arbóreas y el contenido de Al edáfico en bosques del Chocó biogeográfico (Quinto & Moreno, 2014) y en otros bosques lluviosos tropicales (Clinebell et al., 1995). Asimismo, investigaciones realizadas en el Chocó y la Amazonia han mostrado una alta diversidad arbórea sobre suelos con alto contenido de Al (Duivenvoorden, 1996; Faber-Langendoen & Gentry, 1991).
Phillips et al., (1994) concluyeron que los bosques lluviosos tropicales con mayor dinámica (mortalidad y reclutamiento) tienden a presentar alta riqueza de especies. Asimismo, los pequeños disturbios (claros naturales) ayudan a mantener la diversidad. Probablemente, factores como la toxicidad por Al, topografía quebrada, acidez del suelo y baja disponibilidad de nutrientes, podrían generar mayor dinámica del bosque (Asquith, 2002; Laurance et al., 2010; Marschner, 1991); y por ende, una mayor diversidad arbórea (Phillips et al., 1994). Posiblemente esta interacción de factores es la que mejor explica porque se presenta una alta diversidad arbórea en bosques tropicales de alta precipitación, con suelos infértiles y con alta toxicidad por Al (Clinebell et al., 1995; Duivenvoorden, 1996). En síntesis, la alta diversidad arbórea registrada en estos bosques del Chocó está determinada considerablemente por el contenido edáfico de nutrientes. Asimismo, la baja fertilidad edáfica, acidez y toxicidad por Al, generados por la precipitación, están determinando la dinámica y la diversidad arbórea.

