











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
The life history of California sea lion (Zalophus californianus) in the Gulf of California (henceforth, the Gulf) is marked by a series of important events influencing and modifying its population growth, distribution, and evolution. The ancient settlement of local California sea lion demes in the Gulf dates to three million years ago, during the late Pleistocene or earlier (Maldonado, Orta-Dávila, Stewart, Geffen, & Wayne, 1995). Sea lions started to occupy the Gulf due to high primary productivity and large abundance of sardine (Sardinops sagax), a typical California sea lion prey, particularly around the Midriff island region (Fig. 1). The Gulf of California sea lion population progressively became isolated from the US California and Pacific Baja California populations, as demonstrated by several genetic studies (Fig. 1, Maldonado et al., 1995; Bowen et al., 2006; Schramm, 2002).
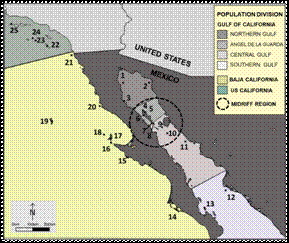
Fig. 1 Genetic division of California sea lion (Zalophus californianus) population in Mexico and California (US) (Maldonado et al. 1995): US California, Baja California and Gulf of California; Gulf of California eco-region subdivision (Szteren & Aurioles-Gamboa 2011): Northern, Angel De La Guarda, Central And Southern Gulf; rookeries distribution: 1. Rocas Consag, 2. San Jorge Island, 3. Lobos Island , 4. Granito Island, 5. Los Cantiles, 6. Los Machos, 7. El Partido, 8. El Rasito, 9. San Esteban, 10. San Pedro Mártir, 11. San Pedro Nolasco, 12. Farallón de San Ignacio, 13. Los Islotes, 14. Santa Margarita Island, 15. Asunción Island, 16. Navidad Island, 17. Cedros Island, 18. San Benito Islands, 19. Guadalupe Island, 20. San Jerónimo Island, 21. Los Coronados Island, 22. San Clemente Island, 23. San Nicolas Island, 24. Santa Barbara Island, 25. San Miguel Island.
Over the past several centuries, the Gulf of California sea lion population stabilized into 13 breeding colonies distributed along the Gulf (Fig. 1). Genetic analysis identified three groups in the same number of regions: North, Center and South (Schramm, 2002), seemingly related with the Gulf’s physical environmental conditions. Females are confined near specific reproductive rookeries (high philopatry) (Maldonado et al., 1995). However, this division is not completely clear: only a few colonies have been genetically examined for each region. Using a multivariate model including genetics, diet and osteoarthritis data and other variables, Szteren and Aurioles-Gamboa (2011) divided the Gulf population into four demes located in the same number of eco-regions: North, Ángel de la Guarda (A.G.), Central and Southern Gulf. González-Suárez, Aurioles-Gamboa, and Gerber (2010) and Ward et al. (2009) obtained similar results yet considered that the distance between colonies is the main cause for their division. Nevertheless, uncertainties remain about the reasons why the sea lion population in the Gulf is comprised of distinct rookeries and where are the true limits. In the present study we use Szteren and Aurioles-Gamboa (2011) eco-regional distribution (ca., Fig. 1).
Even though the population has been studied since the 1950s, research has been rather punctual and fragmentary, particularly focused in the A.G. colonies, followed by Los Islotes. Over the past two decades there has been increased concern about the entire Gulf of California sea lion population dynamics and trend, particularly in the period 2010-2016 with the PROMOBI (Program for Biological Monitoring) and PROMANP (Program for Species at Risk) projects promoted by CONANP (Comisión Nacional de Áreas Naturales Protegidas). Before this period, there were very few censuses conducted simultaneously in all rookeries, which precluded gaining insight regarding key aspects of life cycle, population trend and potential threats in general or at specific sites.
A life table produced by Hernández-Camacho (2001) for California sea lions in the Gulf, with further adjustments in survival rates (Hernández-Camacho, Aurioles-Gamboa, Laake, & Gerber, 2008) was used as a reference for many studies. This life table has some problems because it uses the growth rate found for Los Islotes, the Southern-most rookery (ca., Fig. 1), with environmental conditions very different compared to other sites. This was confirmed in 2015 when the same authors intended to apply surrogate data to estimate the growth rate of two colonies located at a different sites with doubtful results attributed to differences in survival rates between rookeries (see below). In preceding years, Inclán (1999) recognized the need to use a dynamic life table and different survival rates for the rookeries in the Gulf population, given the significant differences between ecoregions. In fact, Ward et al. (2009) used a multivariate state-space model and found synchrony in growth rates and variability between colonies from the same region and high temporal correlation among the three most Northern ecoregions.
Scarcity of census data precluded a long-term evaluation of population patterns, which could contribute to mislead results of population viability analyses (Chirakkal and Gerber, 2010) and our perception about the Gulf of California sea lion population status. Szteren, Aurioles-Gamboa and Gerber (2006) published a study about survival rates and found a 20 % decrease during 1976 and 2004; this alarmed the scientific community. Despite that negative trend, another study considered the population to be stable over 37 years (Aurioles-Gamboa and Hernández-Camacho, 2015). This leads one to conclude that the results of a given study might vary depending on the method and time period considered. In the present work we conducted a thorough review encompassing the last 37 years of California sea lion life history and environment in the Gulf of California. This constituted an opportunity to focus on short- and long-term processes acting on the population and hopefully improve knowledge about the population trend and status and unveil some latent aspects hardly recognizable using a partial vision.
Materials and Methods
Data from different sources since the 1970s to 2018 were collected and analyzed: published papers, master’s and doctoral thesis, and technical reports. Particularly, for the last eight years PROMOBI and PROMANP reports (CONANP, unpublished data, available upon request) were used. Data were organized in sections according to crucial population life history events for better understanding the evolution of environmental and anthropogenic factors.
Results
Hunting Period (1533-1970). First population collapse: California sea lions were hunted historically 1 000-2 000 years before European colonization (Zavala-González & Mellink, 2000). Some 500 years ago, the Comcáac (Seri) people settled along the coast of Sonora, where they hunted sea lions for livelihood around Ángel de la Guarda and San Esteban, the largest islands in the Gulf of California. In 1872, sea lion hunting turned into a commercial activity to extract a low-price oil from the blubber and for the leather industry. Between 1860 and 1888 there was an intensive exploitation (Ronald, Selley, Healey, 1982 cited in Zavala-González & Mellink, 2000), and sea lions started to decrease significantly in the late 1870s. In the late XIX Century, the federal government of Mexico granted sea lion hunting permits to Mexican citizens, which were sold to foreigners. During President Álvaro Obregon’s government (1920-24) hunting was declared open in small amounts per permit.
In 1930, the hunting was banned, but in 1937 the sea lion flesh started to be used as bait for shark fishing. The sea lion exploitation was extended until the 1970s, and came to an end in 1982, yet in the 1990s the sea lion flesh was still used as bait (Zavala-González & Mellink, 2000). The Midriff island region was the most exploited area for sea lions, and adult males were the preferred target. The Mexican federal fisheries agency (Dirección General de Pesca) estimated that an average of 400 adult males were killed annually in the Gulf of California during the mid XX Century (Zavala-González & Mellink, 2000).
In 1966, 10 366 adult sea lions were counted in seven colonies, and in 1991 the count raised to 17 486 animals in the same colonies (Zavala-González, 1993). Despite the large population decline in a 25-year period, apparently no genetic bottle neck took place (González, Aurioles-Gamboa, Gerber, 2010). In the early 1990s, California sea lions were still subject to capture in small anumbers to sell them to dolphinaria and aquaria in Mexico and abroad. To date only stranded individuals treated for illness or malnutrition are collected via special permits in displays at dolphinaria and aquaria.
Post -hunting period (1970-1990). Population recovery: After the hunting period, between 1970 and 1990, an important California sea lion recovery took place both in number of individuals and rookeries. From 1942-53, the population rose from 6 273-12 902 individuals; then there were 10 366 sea lions in 1965 (Lluch-Belda,1969; Zavala-González, 1999); 15 140 (14 colonies) in 1979; and 14 389 (in 12 colonies) in 1981 (Le Boeuf et al., 1983). According to Aurioles-Gamboa (1988), at the end of the 1970s the actual number of sea lions were 20 000 adjusted with correction factors to 25 000 animals. The population numbers continued to growth constantly to the point that some rookeries increased above 30 % from 1965-79 (Le Boeuf et al., 1983). During this period, the highest occupation index (total number of occupied island x 100/total number of islands), and pup production was observed in the Northern and Central Gulf up to San Esteban Island.
The highest presence of sea lions in these regions was probably related to food availability since the Monterrey sardine (Sardinops sagax), Pacific mackerel (Scomber japonicus), herring (Opisthonema spp.) and Northern anchovy (Engraulis mordax) were concentrated in the summer around Ángel de la Guarda and Tiburón islands (Midriff area or Central region) (Aurioles-Gamboa, 1988). The increase of sea lion population was particularly considered to be associated with the dramatic rise of Monterrey sardine biomass during the 1970s until 1988-89 (Cisneros-Mata, Nevárez, Hammann, 1995; Zavala-González, 1999). During 1982-83, one of the most intense El Niño events occurred in recorded history, causing a drastic decrease of US California and Pacific Baja California sea lion populations. The pup count decreased 35 %, and did not return to pre-El Niño levels until four years later (Lowry & Maravilla-Chávez, 2005).
This decrease was related with longer female foraging trips due to lower prey availability caused by El Niño (Trillmich & Ono, 1991), which probably resulted in the worsening of pups' health and their death. Conversely, there were no apparent or significant negative effects in the Gulf’s California sea lion population. Hernández-Camacho et al. (2008) documented at Los Islotes a low pup survival rate during 1980-81. However, during the El Niño the rate was very high, therefore they assumed a null influence of El Niño in the Gulf population. Other authors sustain that the environmental conditions inside the Gulf were not influenced by El Niño and the sea lion population was not affected, at least female fecundity, pup body index, juvenile survival rate or female foraging trip duration that remained similar (Lara-Lara, Holgin-Valdéz, Jiménez-Pérez, 1984; Aurioles-Gamboa & Le Boeuf, 1991; Samaniego-Herrera & Aurioles-Gamboa, 2000; Hernández-Camacho, 2001). Nevertheless, this hipothesis may not be completely certain, because there is evidence that there was a decrease in the number of adult males during El Niño associated with a long phase of warm temperature anomaly during 1979 and 1983 (Shirasago-Germán, Pérez-Lezama, Chávez and García-Morales, 2015). Hernández-Camacho (2001), recorded an increased male mortality in 1983 and assumed that, due to their higher metabolic rate and precocious weaning, males were more prone to environmental factor influence than females. This was confirmed by Aurioles-Gamboa and Zavala-González (1994) in 1982-83 in US Southern California when they found an increase in 6-to-12-month age juvenile deaths. Numbers in the Northern and Central Gulf had increased from the 1960s and during 1966-91 the population rose at an annual rate of 2.1 % (Zavala-González, 1993; 1999). After the strong El Niño event, in 1984 the California sea lion population began to decline (Fig. 2) throughout most of the 1990s.
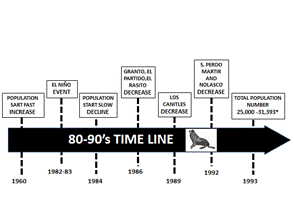
Fig. 2 Most important events occurred during 80-90’s.In 60´s the population fast growing until 1984-89 when the population sarted to decline (Zavala-González, 1993).
At the end of 1980s the sea lion population size was between 25 000 and 28 000 (Aurioles-Gamboa, 1988; Zavala-González, 1990).). In 1990-91, the estimated populations was of 30 800 (Zavala-González, 1993) and for 1993 the total population counted at different times during 1979-1993 in 29 colonies was 17 963 individuals with 5 262 pups, adjusted to 31 393 individuals (Aurioles-Gamboa & Zavala-González, 1994). In those counts, the numbers for the 13 reproductive rookeries was 15 807 individuals and 5 262 pups. Zavala-González (1999) mentioned that such an important increase and variability in population numbers recorded could have been due to an enhanced monitoring effort.
Unfortunately, the data reported are rather imprecise due to differences in the number of colonies surveyed, method of error estimation, and monitoring protocol. Furthermore, to obtain several total annual sea lion population sizes, some missing survey values were replaced using data corresponding to other years (Aurioles-Gamboa & Zavala-González, 1994; Szteren et al. 2006). However, this could result in an erroneous conclusion regarding the actual population size due to high interannual variability of enviromental conditions. Therefore, it is difficult to determine annual population size and trend particularly because the raw data are difficult to access or unavailable. The increase in the Gulf’s California sea lion population during the 1980s was attributed to a concomitant recovery of the Monterrey sardine. Further, a decrease in sardine recruitment in 1984-86 (Cisneros-Mata et al., 1995; Zavala-González, 1999), due to El Niño event in 1982-84 and positive sea surface temperature anomaly in 1989-90, and excess sardine fishing effort during 1990-91 resulted in a collapse of sardine biomass at the onset of the 1990s (Cisneros-Mata et al., 1995). At the same time, the sea lion population began to decline. Some authors hypothesized that the Gulf’s California sea lion abundance trend is directly related to sardine biomass rather than climatic events per se (Aurioles-Gamboa & Zavala-González , 1994; Szteren et al. 2006).
Global warming and fishing pressure (1990-2000). Second population collapse: As already mentioned, from 1989 to 1992 the Monterey sardine catch in the Gulf dropped from 292 000 metric tons (mt) in 1988-89 to 7 500 mt in 1991-1993 (Cisneros-Mata et al., 1995) and did not recover until 1995. Consequently, sardine predators declined during 1990-94 (Zavala-González, 1999), and simultaneously the Ángel de la Guarda Island (A.G.) California sea lion colonies started to decrease. Different to other colonies, A.G. colonies showed a dominance of sardine in their diet (García-Rodríguez & Aurioles-Gamboa, 2004) which leads to the conclusion that this sea lion colony is more dependent on prey availability (Sterzen & Aurioles-Gamboa, 2006, 2011). Aurioles-Gamboa and García-Rodríguez (1999) found that during the 1980s, pup abundance in Los Cantiles considerably increased in conjunction with the Monterrey sardine biomass peak and rapidly decreased with the collapse of sardine during the early 1990s; therefore they assumed the existence of some reletion between the two events. The latter is not totally supported by Zavala-González (1999), who did not relate the sea lion abundance drop in Los Cantiles with the sardine biomass because the pups’ nutritional condition remained constant during 1988-1993.
The decline of the rookery was most likely related to emigration of sea lions out of the area rather than individual mortality (Heat et al., 1994 cited in Zavala-González, 1999); either way, the pup production had begun to decline years before (Zavala-González, 1999). To summarize, a probable cause of California sea lion decrease in the Gulf was due to the increase of sardine fishing and a reduction of the sardine’s natural productivity due to changing oceanographic conditions (Zavala-González, 1999). In support of this hipothesis, in 1993 the female foraging trips at Los Cantiles was longer than during 1988 probably related to prey availability (Heat et al., 1994 cited in Zavala-González, 1999). This might explain the reduction in sea lion female numbers during the onset of the 1990s in the central Gulf rookeries (Zavala-González, 1999). Despite that El Niño 1992-93 has been considered as a moderate event, it coincided with a long period of positive sea surface temperature anomalies (Zavala-González, 1999) which resulted in a major Gulf ecosystem effect.
During 1960 to 1990, in the Central Gulf and Aangel de la Guarda (A.G.) the sea lion population increased 2.1 % per year (Zavala-González, 1993, 1999). From 1984 to 1991 in eight colonies of this area the population was 12 600, and in 1990-1993 dropped 34.8 % (Maravilla-Chávez et al. 1997 cited in Zavala-González, 1999). As already mentioned, in these two areas the population had begun to decline since the 1980s but while colonies in the San Pedro Nolasco island and Los Cantiles ceased to decline in 1993 (58.6 and 29.9 %, respectively), the other colonies continued to show decreased sea lion numbers until 1997 (Zavala-González, 1999). Zavala-González (1999) assumed that since 1984, environmental changes had occurred in the Midriff island region which increased in 1990-93 and affected the sea lion population. This explains the 34.8 % decrease in total numbers, 37.1 % of females in 1991-92, 38 % of males in 1992-9 and 46.7 % of pups in 1990-93. In contrast, during 1985-1992 Ward et al. (2009) found a population drop greater than expected according to Zavala-González’s hypothesis.
Conversely, the US and Pacific Baja California sea lion populations did not experience the same process and, although decreased during El Niño 1992-93, the pup production recovered rapidly than in 1983. After having declined to just 8 902 individuals in 1993, the central and A.G. sea lion population remained constant around 9 237 individuals until 1997. Surprisingly, during the strong 1997-98 El Niño event the population did not declined compared to the US population where the pup production decreased significantly (Lowry & Maravilla-Chávez, 2005). In the Central Gulf region, during 1993-1997 the pup production increased by 54.1 % (Zavala-González 1999). In Los Islotes, pup numbers increased, and their body condition was similar to that in other years (Szteren et al., 2006; Hernández-Camacho et al., 2008). Neither of both strong El Niño events in 1983 and 1997 impacted the population, and it has been proposed that the Gulf’s environmental conditions during 1997 favored enough prey availability (Hernández-Camacho et al., 2008). This has not been substantiated with data. For 1998 in San Jorge Island (Northern Gulf), Mellink (2001) found an anomalous increase of adult individuals (6 717) without variation in pup numbers (793) probably due to immigration of individuals from the central and Southern rookeries. Later, during the 1999 La Niña, the author recorded more pups than in previous years (1 053). Yet the number of adults (2 953) varied slightly with respect to previous years (before El Niño), probably because the return of individuals to their original rookeries. This hypothesis of sea lion population movements to more favorable sites resembles the shift observed in seabirds nesting from Rasa Island south of A.G. to San Jorge Island during 1997-98 (Velarde, Ezcurra, Horn, & Patton, 2015).
Zavala-González (1999) interpreted the increase in sea lion pup numbers as a strategy developed over short time periods to confront strong environmental pressures, such as what occurred in the US population (Lowry et al., 1992 cited in Zavala-González, 1999). A decrease of pups and juveniles was not seen at Los Islotes during1997-98, Hernández-Camacho (2001) found fewer adult males and females. Although this author interpreted the latter as being due to extended feeding trips and not to mortality, Samaniego-Herrera (1999) did not report changes in food searching times but did find a fecundity increase for the same time period. This means that a greater number of pups than females were observed in the rookery, which seems to confirm Hernández-Camacho’s theory.
Despite pup recruitment being high, Samaniego-Herrera (1999) found changes in juvenile numbers which suggests movements to more favorable areas rather than a mortality increase. Moreover, during the El Niño 1997-1998, Samaniego-Herrera (1999) found a diet shift from grunts (Haemulopsis spp.) and groupers (Serranidae) in 1990-93 to flagfin (Aulopus bajacali), hairtail (Trichiurus lepturus), lantern fishes (Myctophidae) and mollusks (Cephalopoda). These findings seem to support an adaptive strategy of California sea lion in response to environmental conditions (Samaniego-Herrera, 1999).
The 35 % population decrease during the 1980s and 1990s should not be attributed neither to pups -that showed an increase despite intense interannual variability, nor to adult males -that also increased- but to females abundance which had been decreasing over 14 years. This process generated in the Central Gulf rookeries a change in sex ratio from 1:10.9 (male:female) in 1984, to 1:4.3 in 1997, probably related to female mortality and a sharp drop in reproduction activity (Zavala-González, 1999). However, in San Pedro Nolasco rookery in 1999, relatively older females gave birth which suggested an increase in juvenile mortality (Aurioles-Gamboa, Godínez-Reyes, Hernández-Camacho, & Santos del-Prado-Gasca, 2011).
During warm sea surface temperature periods (1983, 1987, 1992 and 1998) each age class and sex behaves in different way (Shirasago-Germán et al., 2015). These authors found that during El Niño events adult female numbers decreased more than all male age classes and that male were more likely to decline during prolonged warm periods for the reasons previously discussed. They found a strong negative correlation between El Niño and total adult males and females and interpreted this as evidence of the impact of environmental factors on breeding California sea lion colonies.
In addition to climatic and ecological phenomena, the intensification of fishing effort could have contributed to the population drop of both, small pelagic fishes and California sea lions. During the 1980s and 1990s an increase of sardine fishing occurred, particularly in the A.G. region during summer, which coincides with the California sea lion reproductive season. Furthermore, in the same period, long line and entanglement net shark artisanal fishing notably increased, generating direct and indirect problems for the California sea lion population (Zavala-González, 1999). Sea lions were often found entangled in fishing gears and provoked an increase in fishermen unrest and sea lion shooting occurrences (Gallo-Reynoso, unpublished data). Also, fishers started to use sea lion flesh as bait for shark fishing (Gallo-Reynoso, 1986; Delgado-Estrella, Ortega-Ortíz, & Sanchez-Ríos, 1994).
The first research on the above-mentioned topic was conducted in 1981 in Guaymas Sonora (Fleischer & Cervantes, 1990 cited in Zavala-González, 1999) and at the end of 1980s (Aguayo, 1989 cited in Zavala-González, 1999). Delgado et al. (1994) reported an increase of sea lion entanglement when, due to the decrease of shrimp productivity, fishers began to use monofilament nets for sardine and shark fishing in the A.G. region and near Los Islotes colonies. That same year, sea lions were declared under special protection by Mexican government due to the increased anthropogenic threats (Zavala-González, 1999). In 1997, the situation worsened because the artisanal fishing effort further increased during El Niño. That year in A.G. and the Central Gulf, the entanglement index (percentage of entangled animals/total population) increased to 1.2 % (Zavala-González & Mellink, 1997). The rookery with a major risk for juveniles and females was Los Islotes where the entanglement index oscillated between 3.9 and 8.0 % until 2004 (Aurioles-Gamboa et al., 2011); interistingly, the sea lion population continued to increase.
In some colonies of Northern and Midriff regions, a relationship was found with croacker (Pisces, Scianidae) commercial fishing and a high index of entanglement (Aurioles-Gamboa & Porras, 2006 cited in Aurioles-Gamboa et al., 2011). Aurioles-Gamboa, García-Rodríguez, Ramírez-Rodríguez, and Hernández-Camacho (2003) found in Los Islotes sea lion diets an low overlap of 5 %. The commercially important fish species was spotted sand bass (Paralabrax maculatofasciatus). Finally, during 1998-99 in San Pedro Mártir, Aurioles-Gamboa et al. (2011) registered a 10 % entanglement during the peak of shark fishing and assumed that shark overexploitation had generated an increase of giant squid (Dosidicus gigas) that began to compete with sea lions for sardine. These authors concluded that sea lions migrated to other areas which resulted in a 61 % population drop at the San Pedro Nolasco sea lion colony in 2000. After the shark fishing decreased, in 2003 the rookery recovered considerably in numbers (Aurioles-Gamboa at al., 2011). Interestingly, Zavala-González (1999) did not report important sea lion population changes over a 55-year time period analyzed: a stable age distribution and pup growth rate was interpreted by this author to reflect a constant population size.
Climatic moderate variability (2000-2010). Population stability: in 2004 the Gulf’s California sea lion population was 13 185 individuals with a pup production of 4 299; the total number ranged between 24 062 and 31 159 with 8 598 pups (Szteren et al., 2006). The total population decreased 20.05 % compared to the last survey (Aurioles-Gamboa & Zavala-González, 1994) but each colony varied differently: Los islotes 50.6 %, Rocas consag 42.5 %, San Jorge island 12.3 %. San Esteban island sea lions increased in number and fecundity, while Granito island, Los cantiles, Los machos, El partido decreased in different amounts and Farallón de San Ignacio decreased significantly (around 45 %) in both indicators; Lobos island, El Rasito and San Pedro Mártir showed strong fluctuations.
The colony of San Pedro Nolasco Island recovered during 1992 and 1997 and stabilized in 2003. Colonies in San Esteban and Los Islotes showed the highest fecundity rate (up to75 %) and Lobos Island and Rasito the lowest (down to 26 %). Colonies at Angel de la Guarda (A.G.), Los Cantiles and Los Machos were the most affected because the net population growth rate was less than one (Szteren et al., 2006). Total population growth decreased during the period of 1994 to 2004. Szteren et al. (2006) attributed the population decline to low prey availability, particularly sardine, as showed in Central Gulf and A.G. trends (García-Rodríguez & Aurioles-Gamboa, 2004) (Fig. 3).
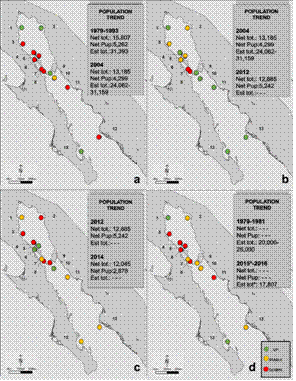
Fig. 3 Spatio-temporal trends variations of California sea lion rookeries in the Gulf of California during different time frames: A. 1979-1993 (Aurioles-Gamboa & Zavala-González 1994) and 2004 (Szteren et al. 2006); B. 2004 and 2012 (Aurioles-Gamboa & Gallo-Reynoso 2012); C. 2012 and 2014 (Gallo-Reynoso et al. 2014); D. 1979-81(Aurioles-Gamboa 1988 and Zavala-González1990) and 2015 -2016 (Gallo-Reynoso et al. 2015 and Pelayo-González 2018). See Fig. 1 for location name codes. NET TOT AND PUP: Net value of total number of individuals and pups. EST TOT: total population adjusted value. GREEN DOT: population number increase, YELLOW DOT: stable number and RED DOT: number decrease.
According to Pérez-Lezama (2010), Szteren‘s (2006) data represented a segment of the cyclic population abundance and not a true decrease. Furthermore, Aurioles-Gamboa & Zavala-González‘s (1994) survey included more colonies censed in different periods. Abundance numbers in 2004 as compared to 1979 or 1997 showed a decrease of 4.8 and 7.9 %, respectively. Including the corrections due to lack of colonies in Szteren’s (2006) data (they used other years’ numbers for two colonies not censused) the average varies from -2.9 to 0.65 %.
Pérez-Lezama (2010) agreed with Szteren et al. (2006) regarding a relation between a drop of sea lion numbers and chlorophyll a concentration and prey availability but attributed the decline to local feeding emigration of some sea lion age-classes, particularly juveniles, to more productive areas. This explains the need to consider not only short-time frames but also the long-term patterns and differences within regions and between age classes. This last aspect was widely demonstrated by Hernández-Camacho et al. (2015) who applied birth and adult survival rate surrogate data (obtained from Los Islotes) to estimate population trend of different colonies: survival rate fitted well between observed and surrogate data for San Jorge Island but not for Granito. This error yielded an estimate of adult female survival rate lower than that for juveniles which is unusual for large mammals.
High energy demand for gestation and the need to remain near the reproductive site for lactation could result in a major adult female mortality rate in an area with environmental stress such as the Midriff region islands. The mechanisms for the latter are unknown, and future investigations should focus on estimating adult survival rates and mortality. Neglecting differences in age-class survival rates could mislead interpretations in large-scale population trend determination; surrogate data should be used only with colonies that show a similar trend and belong to the same area (Hernández-Camacho, Bakker, Aurioles-Gamboa, Laake, & Gerber, 2015). Ward et al. (2009) proposed a multivariate state-space model that consider the different trends based on rookery’s distribution in the Gulf. This approach allows to appreciate spatial patterns of synchrony and correlation. These authors found that all northern colonies have similar trends and the southern ones show a negative correlation with all other colonies. Due to the independence of the Southern Gulf region, Ward et al. (2009) concluded that there is a low risk of decline for the whole Gulf California sea lion population yet in the future, the center of abundance would probably shift southwards with an increasing contribution to the total population from 20 % in 2006 to 33 % in 2030.
The Blob (2010-2016). Fast recovery and third population collapse: during 2010 a monitoring project took place by CONANP in collaboration with Mexican research institutions such as Centro Interdisciplinario de Ciencias Marinas (CICIMAR), and Centro de Investigación en Alimentación y Desarrollo (CIAD). The goal was to obtain data over a period of five consecutive years in all rookeries to better understand the state of the sea lion populations. In 2011 12 157 individuals and 4 089 pups were counted; the fecundity index was 60 %, with good pup body index. The majority of the 13 colonies were stable, recovering (San Esteban Island, San Pedro Mártir Island), or increasing (Los Islotes); in San Jorge island the colony was declining albeit a high fecundity rate was also observed.
Compared to 2004, 2011 showed a slight decrease of population (13 185), with 4 299 pups (Aurioles-Gamboa et al., 2011). In 2012 the population increased to 12 885 adults and 5 242 pups 1 000 pups more than in year 2011, numbers similar to those dating back to the 1980s. In both years (2011 and 2012) the highest index of fecundity was found in San Jorge Island, Los Cantiles, Los Machos, Farallón de San Ignacio and Los Islotes, although only the last two colonies increased in number (Aurioles-Gamboa & Gallo-Reynoso, 2012). Some colonies experienced an important change in trend: Farallón de San Ignacio showed an increase, after a period of important decline (around 45 %), starting in 2004; after a long constant decrease for 30 years (Szteren et al., 2006; Ward et al., 2009) in 2013 Los Cantiles and Los Machos reached the highest number of the past 15 and 20 years, respectively. That same year, Granito island, El Partido and El Rasito slowly increased in sea lion numbers; conversely, in the latter years Los Islotes continued to increase but at a lower rate (Aurioles-Gamboa, Gallo-Reynoso, & Hernández-Camacho, 2013) (Fig. 4).
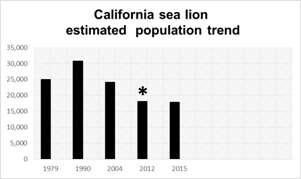
Fig. 4 Gulf of California sea lion (Zalophus californianus) total population trend (estimated value) during 36 years of investigation (1979-2015).*net value*. Total population number estimated during the period 1990-93 (Aurioles-Gamboa, 1988; Zavala-Gonzalez, 1990and 1993; Aurioles-Gamboa & Zavala-González, 1994).
The entanglement problems registered some changes as well. Before 2004 Los Islotes colony had the highest rate but in recent years the northern and central regions showed the highest entanglement rate, in particular, females (San Jorge) and yearlings. In contrast with 1997 the entanglement rate decreased with respect to the 1990s with a further decline during El Niño 2015. Although apparently this did not affect the growth rate, there is a great level of concern among researchers particularly due to a recent worsening again of the problem in risk areas as Midriff region (Aurioles-Gamboa & Gallo-Reynoso, 2012; Gallo-Reynoso, 2015). Until 2013 the growth and fecundity rates remained stable over the last 20 years (Szteren et al., 2006; Ward et al., 2009; Aurioles-Gamboa et al., 2013). The problems faced by the California sea lion population began in 2014 when the total number (12 045) and pup production (2 878) decreased 18 and 45 %, respectively as compared with 2012. Even though the pup production continued within range, exclusively San Pedro Mártir registered one of the lowest values in the last 20 years prior to 2012.
In 2014 the fecundity rate between the colonies was more variable and lower than in 2012 (30 to 100 %). In 2015 in 11 colonies the total number of individuals was 7 966 (17 807 estimated) 16 % less than 2014 but the pup production increased 19 % (3 070 adjusted to 6.748) (Gallo-Reynoso, Aurioles-Gamboa, & Hernández-Camacho, 2015; Hernández-Camacho et al. unpublished data cited in Pelayo-González, 2018). In 2016 the joint monitoring project came to an end and institutions did not share their information. Consequently, it is impossible to know each rookery’s population trend for that year. Partial data from another CONANP monitoring project shows a slight improvement of the sea lion population in five colonies distributed in different areas of the Gulf, except the A.G. Region (Gallo-Reynoso, Aurioles-Gamboa, & Hernández-Camacho, 2016).
To summarize, between 1979 and 2016 an important total Gulf’s California sea lion population decrease was estimated at 44 %, and pup production at 36 %, under historical average in the Northern and Central Gulf during 2015-16 (Fig. 4) (Hernández-Camacho et al. unpublished data cited in Pelayo-González, 2018). Each colony shows different growth rate and risk of extinction or extirpation: increasing at Rocas Consag and Los Islotes; decreasing in Lobos Island, El Granito Island, Los Cantiles, Los Machos, El Rasito and San Pedro Nolasco; stable at San Jorge Island, El Partido, San Esteban, San Pedro Mártir and El Farallón de San Ignacio (Pelayo-González, 2018) (Fig. 3). This result could reflect a state of the Gulf’s environment ecosystem during 2011-2016. After two consecutive La Niña events in 2013 and later in 2014 in the northern Pacific Ocean, an important sea surface anomaly around 1-4 °C, named “The blob” (NASA) which generated a decrease in primary productivity and of California sea lion prey availability (Elorriaga-Verplancken, Ferretto, & Angell, 2015; Kintisch, 2015). After that, in 2015-2016 a strong El Niño event also developed (NASA).
This series of climatic events determined profound ecosystem changes with important consequences. The sardine catch decreased from half a million mt in 2008-2009 to 3 500 mt in 2013-14 (Velarde & Ezcurra, 2015); migratory birds in 2015-2016 (different to El Niño 1997) did not nest in Rasa Island but in the Southern California Bight (US) (Velarde et al., 2015). US California sea lion abundance during 1975 to 2014 dramatically increased from 50 000 to 340 000 (McClatchie et al. 2016; Laake, Lowry, Delong, Melin, & Carretta, 2018). In 2011 the number of pups were the highest ever registered (61 943; Carretta et al., 2016) but during 2013-2016 the population declined significantly. In February and March of 2013 and 2015, a massive stranding of starving sea lion pups named “unusual mortality event” was observed in the Pacific US westcoast (NOAA). This event also involved the Pacific of Baja California, California sea lion population that experimented a decrease of pups and adult females in 2014 (Elorriaga et al., 2015) and in the same region in 2015 it was registered an anomalous dispersion of Guadalupe fur seal to northern areas in California, Oregon and British Columbia (Canada) (Aurioles-Gamboa, Rodríguez, Rosas, & Hernández-Camacho, 2017). In 2015, a high number of Guadalupe fur seal juvenile and pups were found stranded along the coast of California (Aurioles-Gamboa et al. 2017). The US California sea lion population suffered a decrease in juvenile numbers (DeLong et al., 2017) with specific low recruitment of two- and five-year old females. Even when in 2012 the pup production was very high in the US, the cohort of two-year old pups was the lowest recorded since 1998 (Laake et al. 2018).
Researchers found that this decrease in sea lion population is not directly related to the El Niño event but to sea surface temperature (SST) anomalies that correlate with pup and yearling survival even if not always appreciated due to the presence of biological and anthropogenic factors. The strong relationship seems to indicate an impact of climate change on sea lion population trends particularly in the US stock was found when positive SST anomaly exceeds 1 °C the population stops to increase and up to 2 °C start to decline (Melin, Orr, Harris, Laake, & DeLong, 2012; Laake et al. 2018). Pelayo-González (2018) obtained similar results in the Gulf of California. The author did not find any relation between pup production and El Niño events, but registered positive correlations when the SST anomaly surpassed + 0.5 °C in the northern and 1 °C in the central region; no relationship was found for the southern region.
Therefore, the observed sea lion decline in the northern and central regions could be related to a change in prey availability generated by positive SST anomalies. In the southern region the highest prey variety (Brusca et al., 2005 cited in Pelayo-González, 2018) could buffer this effect on sea lions. Pelayo-González (2018) found a positive relationship with small pelagic fish availability only in Los Cantiles which decreased with the sardine collapse. During execution of the PROMOBI projects, differences in the diet composition were found between colonies: in 2012, sea lions in San Jorge Island were the most generalist and San Pedro Nolasco the most specialist. An increase of 15N stable isotope reflecting a widening of the trophic niche (Gallo-Reynoso, Aurioles-Gamboa, & Hernández-Camacho, 2014) since 2004 was also found in the central region, during 2011-12 in the northern Gulf colonies and in 1999 in Los Islotes. This shows that at least three geographic areas are influenced by different environmental factors forcing sea lions to change their diet (Gallo-Reynoso et al., 2014). Particularly in 2015 it was found an increase of crustaceans which were more frequent than cephalopods, different to what was observed in1997. This might be related to the El Niño event but the presence of crustaceans in California sea lion diet had never been so high (Gallo-Reynoso et al, 2015).
Discussion
Considering a long-time frame period (1979-2016) the sea lion population could appear stable with zero growth due to extreme interannual fluctuations (Gallo-Reynoso Aurioles-Gamboa, Hernández-Camacho, 2015; Pelayo-González, 2018). After sea lion hunting came to an end in the 1970s (Zavala-González & Mellink, 2000), the population showed a constant increase until the onset of the 1990s when it drastically decreased. However, there is evidence that this low trend apparently started in 1984 (Zavala-González, 1999). Then, followed a period of relative stability (Pérez-Lezama, 2010) until La Niña event in 2011-2012 (Aurioles-Gamboa & Gallo-Reynoso, 2012) when the population showed an important growth but to levels reported in 1980s.
The population began to decrease during El Niño of 2015-2016 (Gallo-Reynoso et al., 2015). According to various authors (Aurioles-Gamboa & Zavala-González, 1994; Zavala-González, 1999; Peréz, 2010; Laake et al., 2018; Pelayo-González, 2018), the cyclic population fluctuation could reflect dependence with climatic factors, not directly with El Niño, but with SST anomalies that determine changes in prey availability. This seems to impact more the area where Monterrey sardine (Sardinops sagax) forms an important component of sea lion diet such as the A.G. region (Aurioles-Gamboa & Zavala-González ,1994; Pelayo-González, 2018) thus increasing the probability of extirpation (Gallo-Reynoso et al., 2014). The Southern region represents least concern due to major diet variability, so that it has been expected that in the year 2030 the southern colonies will comprise 33% of the total population (Ward et al., 2009; Pelayo-González, 2018). However, many doubts persist about the influence of environmental site-specific factors on local sea lions particularly related with juvenile recruitment and female survival rate (Zavala-González, 1999; Hernández-Camacho et al., 2008; Hernández-Camacho et al., 2015). Given the present state of knowledge, due to numerous inconsistencies reported in this review, it is difficult to conduct an assessment of California sea lion population in the Gulf. Particularly, because the data pre 1990’s are rather vague so it is difficult to compare these data with more recent investigations. Furthermore, between 1998 to 2010 there is an important information gap (the only year completed was 2005), so it is impossible to compare data among all ecoregions except for few years.
The only information comparable consist in the data collected during 2011-2016 because they show spatio-temporal consistency and standardized methodology. Hence, to improve the knowledge about the status of California sea lion population there is a need to conduct future investigations based on the recent years. To better understand which are the main factors that regulate population trends in California sea lions, more information is required based on seasonal surveys in all reproductive and a selection of resting colonies. Data on diet, sardine fishing pressure, migration rates, genetics, prey availability, life cycle and regional environmental variation must be considered to ascertain short- and long-term population and colony spatial-temporal patterns. Statistical errors need to be assessed and monitoring methods should be standardized. It is important to promote research on related topics such as harmful algal blooms, pathogens, persistent organic pollutants and heavy metal tissue concentrations. These factors may influence sea lion mortality/fertility rate and the lack of these data could generate errors in the construction of life tables and population viability analyses.

