











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Las hormonas esteroideas sexuales 17ß - estradiol y progesterona actúan como mensajeros químicos y participan en el control de procesos como la diferenciación sexual, el mantenimiento de las características sexuales, la maduración de gametos y el comportamiento sexual (Norman & Henry, 2015; Ogino, Sato, & Iguchi, 2016; Vardanyan & Hruby, 2016). La presencia y biosíntesis de estas hormonas se ha demostrado en diferentes invertebrados (Janer, 2006; Fernandes, Loi, & Porte, 2011). En moluscos marinos la biosíntesis de 17ß - estradiol está registrada en Haliotis diversicolor (Zhou et al., 2011), Helix aspersa (Le Guellec et al., 1987) y en Clamys farreri (Liu, Xu, Robinson, Qin, & Li, 2014); así como la biosíntesis de progesterona en Clione antartica (Hines, Bryan, Wasson, McClintock, & Watts, 1996). Sin embargo, las funciones que desempeñan estas hormonas en los moluscos aún no han quedado del todo claras, aunque su presencia se relaciona con los eventos reproductivos. Las concentraciones más altas de 17ß - estradiol se han encontrado en los periodos de madurez de hembras de Crassostrea gigas (Matzumoto, Osada, Osawa, & Mori, 1997) y Sinonovacula constricta (Yan, Liu, Ke, Yu, & Kong, 2011). Por otra parte las concentraciones de progesterona incrementan durante la etapa de gametogénesis en Biomphalaria alexandrina (Omran, 2008).
En acuacultura el uso de hormonas ha permitido mejorar la calidad de los gametos, producción y control de la madurez gonádica de cultivos de reproductores (Mylonas, Duncan, & Asturiano, 2017; Wang et al., 2008). En el bivalvo Crassostrea gigas al inyectar 17ß - estradiol en la gónada se incrementa el diámetro de los ovocitos y la producción de vitelo (Li, Osada, Suzuki, & Mori, 1998) y en el caso de Placopecten magellanicus sirve para inducir el desove en (Wang & Croll, 2006). Hasta el momento, no hay registros sobre la implementación de estas hormonas en gasterópodos con fines reproductivos.
El caracol rosa (L. gigas) es una especie del Caribe que representa un negocio lucrativo para los mercados de Estados Unidos y Europa (Acosta, 2006; FAO, 2010). No obstante, su sobreexplotación ha provocado el declive de sus poblaciones, por lo que desde 1992 se encuentra en la lista de animales protegidos (Apéndice II Cites). El desarrollo de técnicas para su cultivo de la etapa larval hasta juveniles, comenzó desde mediados de los años 70 y continuó hasta principios de los años 90 (Davis, 2000). Desde ese momento la larvicultura tuvo un constante desarrollo, dejando a un lado a los reproductores. Si bien Shawl y Davis (2004) describen la obtención de huevos al colocar hembras y machos de esta especie en un área confinada, no se han sido realizados más estudios sobre el establecimiento del cultivo de reproductores e inducción al desove.
A comparación del amplio desarrollo de técnicas para la mejora reproductiva en otras especies de moluscos con casos similares e.g. Haliotis spp. y Trochus niloticus, la información en el caracol rosa es limitada. La única granja de cultivo activa de L. gigas depende de la temporalidad reproductiva y la colecta de masas de huevos del medio natural para continuar con el ciclo. A pesar de que el control de la reproducción es un elemento necesario para un desarrollo sostenible de esta actividad (Mylonas et al., 2017). Por ello el objetivo del presente trabajo es determinar la presencia de las hormonas esteroides 17ß - estradiol y progesterona en L. gigas a través de un método no invasivo y comparar dos técnicas para su cuantificación. Con base en los aportes de este estudio se determinará la viabilidad de ambas técnicas, costos, requerimientos, facilidad de manejo, y tiempo de análisis con el fin de continuar con los estudios reproductivos de esta especie.
MATERIALES Y MÉTODOS
Área y recolecta de muestras: Organismos adultos de L. gigas (n = 10) fueron recolectados a través de buceo libre cada dos meses durante un año en la caleta del parque Xel- Há, Quintana Roo, México (20°18’50”- 20°19’17” N & 87°21’45.5”- 87°21’02.5” W). Posteriormente se determinó el sexo y se colocaron en acuarios por un lapso de 5 horas, a una densidad de un organismo en 40 L de agua de mar. Las heces emitidas se recolectaron de forma individual con una pipeta Pasteur y se conservaron a una temperatura de entre 2 y 4 º C para su traslado al laboratorio de Biología y Cultivo de Moluscos del Cinvestav Mérida. Al finalizar la toma de muestras los organismos fueron regresados al sitio de colecta.
Extracción de hormonas esteroideas: Un gramo de heces fue pesado y mezclado con 10 ml de metanol al 80 % (Palme, Touma, Arias, Dominchin, & Lepschy, 2013). Esta mezcla fue homogenizada por de agitación manual; posterior a su sedimentación, el sobrenadante fue recuperado, filtrado (Millex-GV 0.22 µm) y dividido en dos frascos (uno para cada técnica). Las muestras se preservaron a -20 °C hasta su análisis.
Enzimoinmunoensayo (EIA): Las concentraciones de 17ß - estradiol y progesterona en heces de L. gigas fueron medidas usando el kit de EIA multi especie DetecX (Arbor Assay Inc.) específico para cada hormona. Este Kit permite medir la concentración a través de la unión de la hormona a un anticuerpo y un marcador (tetrametilbenzidina) con el cual se obtiene una reacción colorimétrica medible a través de espectrofotometría. La reactividad cruzada con metabolitos de cada hormona reportada es máxima de 0.02 %.
Para llevar a cabo la cuantificación de hormonas se estableció una curva de concentraciones estándar a partir diluciones de una solución patrón. De cada muestra se tomaron 100 µl y se diluyeron en 400 µl de Buffer de ensayo, de esta mezcla 50 µl se analizaron por duplicado siguiendo las instrucciones del fabricante. La reacción colorimétrica se midió a 450 nm en un espectrofotómetro de placas (Dynex Technologies). Las densidades ópticas registradas se analizaron con el software MyAssays (MyAssays Ltd), el cual calcula la concentración a través de una regresión logística de cuatro parámetros a partir de la curva de calibración. El resultado obtenido se multiplicó por el factor de dilución, el cual se calculó dividiendo el volumen final entre el volumen inicial de la muestra (Helms, 2008).
Cromatografía liquida de alta eficacia (HPLC): Los reactivos químicos como 17ß - estradiol y progesterona (99 % de pureza), agua y metanol grado HPLC (99.8 % de pureza) se adquirieron en Sigma (St. Louis, MO, USA). El equipo constaba de una bomba de pistón reciproca de doble acción (LC-20AT; Shimadzu, Japan), un automuestreador (SIL-20A; Shimadzu, Japan) con control de temperatura (CCTO-20A; Shimadzu, Japan), un detector de luz ultravioleta (SpD-20A; Shimadzu, Japan) y una columna C18 (5 µm, 150 x 46 mm, Restek, Bellenfonte, PA, USA). Para llevar a cabo el análisis de las muestras, la columna y el sistema de HPLC se mantuvieron a 22 °C con un volumen prefijado de 50 µl de inyección de la muestra. Se identificó el tiempo de retención y la tasa de flujo para cada hormona inyectando una solución patrón de 17ß - estradiol y una de progesterona. Posteriormente se construyeron las curvas de concentración estándar a partir de diluciones de las soluciones patrón. Las muestras para la detección de 17ß - estradiol se inyectaron por duplicado a un flujo de 0.8 ml / min, con una fase móvil de metanol / agua, a razón de 50 / 50 v/v y el detector UV fijado a 230 nm. En el caso de progesterona se utilizó una fase móvil de 70 metanol / 30 agua v/v y se inyectaron las muestras por duplicado a un flujo de 1 ml / min y con el detector UV fijado a 254 nm. Entre cada muestra se analizaron blancos de metanol como vehículo para detectar si había interferencias en la elución. Los cromatogramas de la curva de calibración y de las muestras fueron analizados a través del software Shimadzu LC Solutions para calcular la concentración de cada muestra.
Métodos estadísticos: Previo a su análisis la normalidad de los datos fue comprobada con la prueba de Shapiro - Wilks. La comparación de ambas técnicas se llevó a cabo usando el método de ajuste de Passing - Bablok (Passing & Bablok, 1983) junto con el gráfico de diferencias de Bland - Altman (Bland & Altman, 1986) y una prueba t- Student (Sokal, 1995) para la comparabilidad entre técnicas. La comparación de las concentraciones para cada hormona en el tiempo se llevó a cabo con análisis de varianza (ANOVA) y comparación de Tukey (Sokal, 1995). Estas pruebas estadísticas se llevaron a cabo con el software Analyse it (Analyse it Software, Lt).
RESULTADOS
17ß - estradiol EIA versus HPLC: Las concentraciones de esta hormona presentaron una variación temporal (Fig. 1A). De marzo a julio se mantuvieron constantes, con concentraciones entre 0.3 a 3.6 ng / ml con EIA y 0.4 a 2.1 ng / ml con HPLC. De septiembre a enero las concentraciones disminuyeron significativamente (P < 0.0001).
La técnica de EIA se tomó como la técnica de referencia. Con el método de Passing - Bablock se obtuvo la ecuación con una pendiente b = 0.5805 (IC 95 % 0.3785 a 1.083) y una correlación r = 0.673 (Fig. 2A). El ajuste entre ambas técnicas sugiere que existe diferencia significativa en la pendiente (t = -8.58, P < 0.0001). El análisis por diferencia de medias de Bland - Altman mostro que la concentración media de 17ß - estradiol con HPLC es de -0.17 ng / ml en comparación a la media de la concentración obtenida con EIA, y son comprables (t = -1.76, P = 0.08) (Fig. 2B).
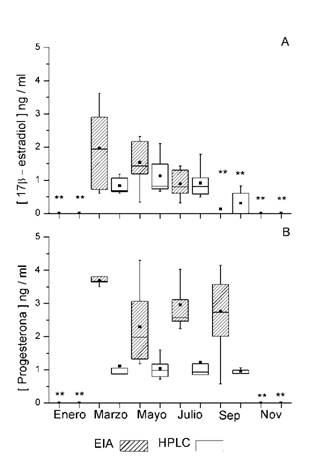
Fig. 1 Gráficas de cajas de la concentración de las hormonas 17ß - estradiol (A) y progesterona (B) en heces de Lobatus gigas en un periodo de un año, usando dos técnicas: Enzimoinmunoensayo (EIA) y Cromatografía líquida de alta resolución (HPLC). (**) representa las diferencias significativas entre meses para cada técnica (P < 0.0001).
Progesterona EIA versus HPLC: La variación temporal de esta hormona presento concentraciones mínimas y máximas de 0.5 a 4.30 ng / ml con EIA y de 0.7 a 1.9 ng / ml con HPLC de marzo a septiembre, las concentraciones durante los meses de enero y noviembre disminuyeron significativamente (P < 0.0001) (Fig. 1B).
Al igual que el caso anterior la técnica de EIA para progesterona se tomó como técnica de referencia. Al analizar las concentraciones obtenidas con el método de Passing - Bablock se obtuvo una pendiente b = 0.1849 (IC 95 % -1.832318 a -1.043762) y una correlación r = 0.389 (Fig. 2C). La comparación de métodos fue significativamente diferente en la pendiente (t = -20; P < 0.0001). El análisis de Bland - Altman indico que la diferencia media de las concentraciones de progesterona de EIA respecto a las obtenidas con HPLC fue de -1.43 ng / ml (Fig. 2D), con una diferencia significativa entre ambos métodos (t = -7.3; P < 0.0001).
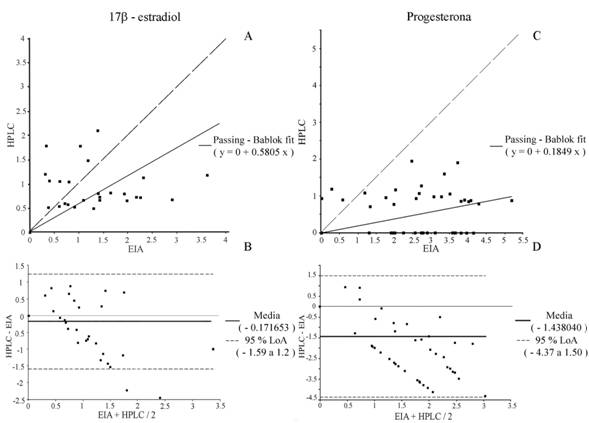
Fig. 2 Ajuste de Passing-Bablock y diferencia de medias de Bland - Altman de la concentración de las hormonas 17ß - estradiol (A, B) y progesterona (C, D) en Lobatus gigas con las técnicas de Enzimoinmunoensayo (EIA) y Cromatografía líquida de alta resolución (HPLC). Simbología (A, C): Misma escala (― ―).
DISCUSIÓN
El análisis hormonal en mamíferos terrestres y acuáticos se realiza por lo general en sangre y tejidos ( Budzinski, Devier, Labadie, & Togola, 2006; Amaral, 2010). En invertebrados, como crustáceos y moluscos se utilizan la hemolinfa y las gónadas (Bose, Majumdar, & Bhattacharya, 1997; Gorbushin & Iakovleva, 2006; Huang, Ye, Han, & Wang, 2009). En el caso de moluscos, la hemolinfa se extrae por punción cardíaca causando daños y la obtención de las gónadas implica el sacrificio del organismo (Matzumoto et al., 1997; Gorbushin & Iakovleva, 2006).
En especies protegidas e indexadas en CITES como es el caso de L. gigas, su colecta y más aún su sacrificio, requieren la obtención de permisos. Por lo anterior, una alternativa para el estudio de hormonas son los métodos no invasivos, a través de las excretas (Busso & Ruiz, 2011; Sheriff, Dantzer, Delehanty, Palme, & Boonstra, 2011).
En moluscos, las hormonas esteroideas han sido cuantificadas con diferentes técnicas. Con HPLC se midió 17ß - estradiol y estriol en las gónadas de hembras del ostión de Crassostrea gigas durante la madurez sexual (Matzumoto et al., 1997). A través de HPLC y EIA se registraron las concentraciones más altas de progesterona, 17ß - estradiol y testosterona durante el desove del bivalvo Mya arenaria (Siah, Pellerin, Benosman, Gagné, & Amiard, 2002; Gauthier-Clerc, Pellerin, & Amiard, 2006). En Achatina fulica con radio-inmuno-ensayos se observó el incremento de17ß - estradiol en la madurez sexual (Bose et al., 1997).
En este estudio, con HPLC y EIA se cuantificaron las hormonas 17ß - estradiol y progesterona. Las concentraciones de ambas variaron en el tiempo, siendo los meses de actividad reproductiva (marzo a septiembre) los que presentaron las concentraciones más altas, en comparación a los meses de reposo sexual (noviembre, febrero), cuyos valores fueron cercanos a cero. Estos resultados demuestran la presencia de un ciclo hormonal en L. gigas y su potencial influencia en el ciclo reproductivo de la especie.
Los resultados obtenidos en esta investigación para la progesterona con ambas técnicas fueron diferentes. Esto podría explicarse por la baja selectividad de HPLC para detectar esta hormona en las muestras (Cross & Hornshaw, 2016).
Por lo que se refiere a la inversión para ambas técnicas, el HPLC tiene un costo entre 10 000 a 25 000 USD y la técnica de EIA de1 500 a 5 000 USD. De esta última se adquieren paquetes que incluyen la hormona específica, los reactivos y los anticuerpos para realizar la reacción siendo de fácilmente el análisis. En cambio, HPLC requiere personal capacitado para calibración del cromatógrafo (Darwish, 2006; Cross & Hornshaw, 2016).
Con base en lo anterior la técnica de EIA y el método no invasivo utilizado en este trabajo son una opción adecuada para el estudio de hormonas en moluscos, en particular de especies amenazadas como L. gigas. El conocimiento generado en este trabajo permitirá monitorear y seleccionar organismos reproductores que se encuentren acondicionados en laboratorios y de esta manera no incidir en la colecta de masas ovígeras silvestre (Li et al., 1998; Klaoudatos & Klaoudatos, 2004; Mylonas et al., 2017).

