











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Los camarones de la Sonda de Campeche son un recurso económico importante y se han estudiado con rigurosidad para apoyar la pesca sostenible (Ramírez-Rodríguez, 2015; Wakida-Kusunoki, González Cruz, Medellín Ávila, & Arreguín-Sánchez, 2010; Wakida-Kusunoki, García-Solorio, & Vázquez Benavides, 2008) y evitar el deterioro irreversible del recurso. La mayoría de los trabajos ecológicos se han centrado en los organismos adultos que son más fáciles de capturar y estudiar, para lo cual a menudo se pesan, miden y sexan con vistas a conocer las características de las poblaciones a medida que transcurre el tiempo. El conocimiento sobre las primeras etapas del ciclo de vida en el medio natural es limitado debido a la dificultad de los muestreos y a lo laborioso del trabajo en el laboratorio.
Como parte de los estudios generalmente se establece la relación longitud-peso a través de modelos de regresión que permiten estimar el peso de un individuo usando la longitud correspondiente. En muchos casos es más fácil conocer la longitud que el peso y conociendo una de las variables se puede estimar el valor de la otra. La relación longitud peso puede estar asociada al sexo, a la especie, a los factores ambientales y puede variar en dependencia de la edad de los organismos. El crecimiento puede ser un buen índice de la fluctuación de las condiciones estacionales y un prerrequisito para la modelación de la dinámica de las poblaciones bajo diferentes condiciones naturales y antropogénicas (Béguer, Rochette, Girardin, & Boët, 2011).
El factor de condición relativa se usa para comparar la condición, grasa corporal o condición física de los animales, basándose en la suposición de que los animales más pesados de una determinada longitud exhiben mejores condiciones físicas. Ambos conceptos se han utilizado en la investigación pesquera desde principios del siglo XX (Froese, 2006) y ayudan a la diferenciación de pequeñas unidades taxonómicas al igual que otras relaciones morfométricas (Le Cren, 1951).
La Laguna de Términos por sus características resulta ser una zona importante para el reclutamiento de postlarvas y para la cría hasta juvenil, (Alvarez, Gracia, & Soto, 1987; Sánchez & Soto, 1987; Gracia & Soto, 1990), etapa en que los animales migran para completar su desarrollo en aguas abiertas.
Teniendo en cuenta que los camarones son organismos residentes en el estuario durante todo el año (Ramírez Rodríguez, 2002), que en la zona de estudio son relativamente numerosos y pueden ser colectados con facilidad, se considera que pueden servir como indicadores que reflejen los cambios que se manifiestan en el ecosistema y contribuir a explicar la dinámica de las poblaciones de este importante recurso (Béguer et al., 2011). Considerando esta premisa el presente trabajo se enfoca a conocer las características biométricas del cefalotórax y de la longitud total, así como la relación con el peso de los organismos colectados y evaluar el índice de condición relativa para determinar si pueden ser utilizados como indicador de las condiciones ambientales existentes en el período de estudio.
Materiales y métodos
Los muestreos se realizaron en la noche, una vez al mes durante la fase de luna nueva, entre los meses de mayo 2011 y marzo 2012. Los sitios incluyeron tres áreas con vegetación acuática sumergida (algas, pastos marinos como Thalassia testudinum, y en menor abundancia Halodule wrightii, y Syringodium filiforme) en la Laguna de Términos, Golfo de México. Los sitios se ubicaron en la parte sur de la isla Carmen en Punta Gorda (18°43' N & 91°33' W), Boca de Matamoros (18°39' N & 91° W) y El Cayo (18°38' N & 91°42' W) (Fig. 1). Los sitios de muestreo se corresponden con el rango de distribución de F. duorarum (Gracia & Soto, 1990). El trabajo se realizó bajo el marco del Convenio de colaboración CONANP (Comisión Nacional de Áreas Naturales Protegidas) UNACAR (Universidad Autónoma del Carmen) con fecha del 2011, que entre sus clausus contempla el monitoreo de especies marinas.
Fig. 1. Fig. 1 Los puntos negros indican los sitios de muestreo en la Laguna de Términos. The black dots indicate the sampling sites in the Términos Lagoon.
Se realizaron tres muestreos por sitio utilizando una red de Renfro (boca de 2.0 × 0.5 m, longitud total de 2.4 m y tamaño de malla de 2.0 mm); desde un lancha fondeada y ubicada perpendicularmente a la costa se desplegaron 25 m de la cuerda de remolque para cubrir un área de muestreo de 50 m2 por arrastre. La red se recuperó a mano a una velocidad de aproximadamente 0.5 m/s. Todas las muestras se conservaron inmediatamente en una solución tamponada al 4 % de formalina en agua de mar. La salinidad, el pH y la temperatura de cada sitio de muestreo se midieron usando una sonda YSI 63 y el oxígeno disuelto se midió usando una sonda YSI 85 (YSI Inc., https://www.ysi.com ).
En el laboratorio, los camarones peneidos fueron separados manualmente usando tamices metálicos de 500 μm, luego fueron identificados y contados. La identificación al nivel de especie se realizó utilizando las características morfológicas descritas en la literatura (Ditty & Bremer, 2011; Lawrence & Won 1986; Pérez-Farfante, 1969, 1970, 1988; Ringo & Zamora, 1968; Williams, 1959). El material separado se conservó en alcohol etílico al 70 % en viales etiquetados hasta el análisis.
Las medidas se obtuvieron a partir de fotografías tomadas con una cámara Canon PowerShot A630 montada en un soporte universal. Para las mediciones se empleó el software Image-Pro Plus v 4.5 (Media Cybernetics, Inc.). Una imagen de un vernier con una abertura de 0.5 mm sirvió como referencia para la calibración del software
La longitud del cefalotórax (LC) se midió desde el margen postorbital hasta el margen posterior del cefalotórax. La longitud total (LT) se midió desde el extremo anterior del rostrum hasta el final del telson. El peso húmedo (P) se determinó usando una balanza digital Mettler-Toledo con una aproximación de 0.0001 g. El sexo fue determinado con base en la presencia del télico en las hembras (H) y petasma para los machos (M). En los especímenes pequeños, es difícil diferenciar entre los sexos, pero se puede distinguir por la forma y posición de los endopoditos en el primer par de pleópodos (Pérez-Farfante, 1969). Aquellos en los que no se pudo determinar el sexo fueron clasificados como indefinido (I).
Toda la información se agrupó según la categoría de sexo predefinida, se utilizaron las opciones de filtros en MS Excel para seleccionar los datos extremos dentro de cada categoría y realizar los análisis con aproximadamente el mismo tamaño de muestra y con la mayor variabilidad asociada al muestreo.
Se realizó una estadística descriptiva de los datos de Peso (P), Longitud del Cefalotórax (LC), Longitud Total (LT) para cada categoría de sexo y se establecieron los modelos de regresión que explican la relación entre la longitud total y la longitud del cefalotórax. Estos modelos (LT-LC) en las diferentes categorías de sexo fueron comparados a través de un análisis de covarianza (ANCOVA) (Sokal & Rohlf, 1995).
Se realizó un análisis de regresión para las categorías de sexo considerando al peso como la variable dependiente y al cefalotórax o a la longitud total como la variable independiente. La relación longitud del cefalotórax - peso-1 (LC-P-1) se estableció usando la fórmula convencional P = a. LCb, donde P representa el peso de los organismos, LC la longitud del cefalotórax siendo a y b los parámetros del modelo. Este modelo potencial se convierte en uno línea mediante la transformación logarítmica quedando como: log P = log a + b log LC, donde "log a" es el intercepto en el eje y, y el coeficiente de regresión b es un exponente que indica crecimiento isométrico cuando está cerca de tres o alométrico si es diferente.
Para describir el comportamiento del factor de condición por mes y por categoría de sexo, se utilizó el factor de condición relativa Kn (KnLC o KnLT) según estuviesen implicados LC o LT respectivamente, donde Kn = peso / pesos medios suavizados, (Le Cren, 1951). Se realizó un ANOVA de dos vías para comparar los valores de Kn por sitio de muestreo por mes y categoría de sexo. Para todos los procedimientos estadísticos, se eligió un nivel de significación P < 0.05 y se empleó el Paquete Statistica, Stat Soft. Inc V. 10 para el análisis estadístico de la información (StatSoft, 2011).
Resultados
Los datos ambientales en los diferentes sitios se encontraron dentro de los rangos establecidos para la región y no hubo diferencias notables entre los registros para los sitios en el mismo mes. La salinidad y la temperatura mostraron una disminución de agosto a diciembre (Fig. 2), lo cual se espera que ocurra para la región en esta temporada climática debido a la nubosidad y presencia de precipitaciones.
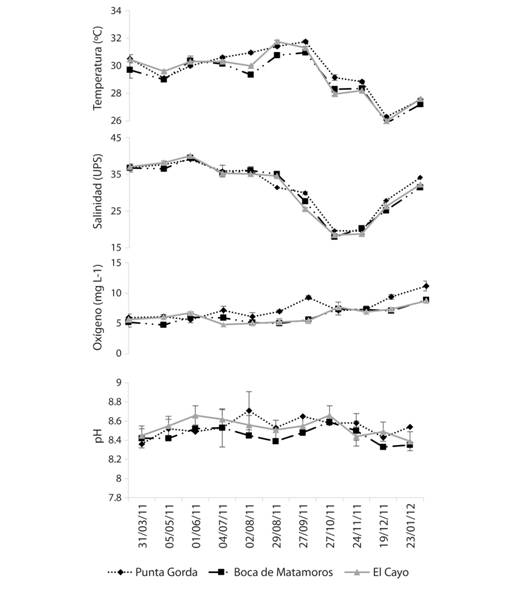
Fig. 2. Fig. 2 Valores medios de los parámetros en los diferentes sitios. Las barras verticales representan el error estándar. Average values of the parameters in the different sites. The vertical bars represent the standard error.
La estadística descriptiva para las diferentes categorías de sexo se muestra en la tabla 1. Todos los parámetros estadísticos se presentan con dos cifras decimales.
TABLA 1 TABLE 1 Parámetros poblacionales para cada sexo Population parameters per sex
| Indefinido | Machos | Hembras | |||||||
| P (g) | LC (mm) | LT (mm) | P (g) | LC (mm) | LT (mm) | P (g) | LC (mm) | LT (mm) | |
| Media | 0.04 | 3.70 | 17.05 | 0.62 | 9.55 | 44.35 | 0.75 | 10.01 | 46.22 |
| ES | 0.00 | 0.03 | 0.13 | 0.02 | 0.11 | 0.49 | 0.03 | 0.12 | 0.52 |
| Mediana | 0.02 | 3.43 | 15.80 | 0.35 | 8.43 | 39.42 | 0.37 | 8.76 | 40.58 |
| Moda | 0.01 | 2.14 | 12.24 | 0.28 | 7.69 | ----- | 0.21 | 6.68 | ----- |
| DE | 0.05 | 1.51 | 6.98 | 0.78 | 4.00 | 18.09 | 1.07 | 4.48 | 20.16 |
| Kurtosis | 17.18 | 0.79 | 0.87 | 13.50 | 5.71 | 5.71 | 19.94 | 3.71 | 3.81 |
| Skewness | 3.25 | 0.96 | 0.98 | 3.26 | 2.00 | 2.02 | 3.88 | 1.78 | 1.80 |
| Min | 0.00 | 1.26 | 7.00 | 0.01 | 3.98 | 18.76 | 0.06 | 3.67 | 17.86 |
| Max | 0.64 | 10.81 | 52.57 | 6.57 | 38.27 | 168.14 | 9.88 | 32.16 | 149.33 |
| n | 3 063 | 3 063 | 3 063 | 1 386 | 1 386 | 1 386 | 1 514 | 1 514 | 1 514 |
| C.L 95.0 % | 0.00 | 0.05 | 0.25 | 0.04 | 0.21 | 0.95 | 0.05 | 0.23 | 1.02 |
ES = error estándar), DE = desviación estándar, Min = mínimo, Max = máximo, n = tamaño de muestra, C.L = 95 % nivel de confianza, P = peso, LC = largo del cefalotórax, LT = largo total.
El sexo en los camarones con un LC pequeño (< 3.67 mm para las hembras, < 3.98 mm para los machos) generalmente puede ser diferenciado. Algunos camarones con CL ≥ 10.81 mm se clasificaron como indefinidos porque las diferencias morfológicas entre los sexos no eran evidentes. Los coeficientes “a" y "b" del modelo LT = a + b LC para cada categoría de sexo se encuentran en la tabla 2. El ANOVA de la regresión indicó que existe correlación entre LC y LT en las diferentes categorías de sexo (P = 0.000). El ANCOVA de los tres modelos de regresión (I, M, H) mostró que las pendientes son significativamente diferentes (F2, 3986 = 12.05, p <0.0001), así como los interceptos (F2, 3988 = 66.53, P < 0.0001). Esto indica que se deben usar diferentes modelos para expresar la relación entre la longitud del cefalotórax y la longitud total de especímenes según el sexo. El ANCOVA mostró además que no hubo diferencias entre las categorías de sexo masculino y femenino para la pendiente (F1, 2676 = 2.46, P = 0. 117) ni para el intercepto (F1, 2678 = 2.41, P = 0.121); por lo tanto, se infiere que los datos se originaron de la misma población estadística y se fusionaron para expresar los nuevos parámetros del modelo (Macho-Hembra (M-H)) (Tabla 3). Las rectas de regresión de los grupos M-H e I fueron significativamente diferentes para el pendiente (F1, 3988 = 21.16, P < 0.0001) y el intercepto (F1, 3989 = 129.6, P < 0.0001); lo cual indica que es necesario usar diferentes ecuaciones para expresar la relación entre LC y LT para especímenes indefinidos y con sexo identificado.
TABLA 2 TABLE 2 Parámetros de la regresión lineal LT-LC para cada categoría sexual Linear regression parameters for LT-LC and sex category
| Categoría sexual | n | Intercepto (a) | pendiente (b) | R2 | p |
| Indefinidos (I) | 1 312 | 0.07 (0.08) | 4.62(0.02) | 0.974 | 0.000 |
| Machos (M) | 1 316 | 1.73(0.20) | 4.48(0.02) | 0.975 | 0.000 |
| Hembras (F) | 1 364 | 1.97(0.18) | 4.44(0.17) | 0.980 | 0.000 |
| Machos-Hembras (M-H) | 2 680 | 1.89(0.13) | 4.46(0.01) | 0.978 | 0.000 |
n = tamaño de muestra, a = intercepto, b = coeficiente de regresión. Los valores en paréntesis corresponden al error estándar de los coeficientes, R2 = coeficiente de determinación, P = valor de probabilidad en el ANOVA de la regresión.
TABLA 3 TABLE 3 Relaciones entre el peso (P) y longitud del cefalotórax (LC) o longitud total (LT) para diferentes especies de peneidos reportada en varios estudios Relationships between weight (P) and length of cephalothorax (LC) or total length (LT) for different penaeid species reported in several studies
| Coeficientes LC-P Forma potencial | Coefficients LT-P Forma potencial | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Estudio | Especie | Localidad | Fecha | Long Cefalotórax (mm) | n | a | b | a | b | |
| May 2011- March 2012 | 1.30-10.81 | Indefinido 1312 | 0.0004 | 3.232 | 0.000003 | 3.224 | ||||
| Actual estudio | F. duorarum | Laguna de Términos | May 2011- March 2012 | ♂ = 3.98-26.01 ♀ = 3.67-32.16 | ♂ = 1 316 ♀ = 1 364 | 0.0016 0.0015 | 2.519 2.544 | 0.00003 0.00002 | 2.583 2.635 | |
| May 2011- March 2012 | ♂♀ = 3.67-32.16 | ♂♀ = 2 680 | 0.0016 | 2.532 | 0.00002 | 2.612 | ||||
| Diaz et al., (2001) | F. duorarum | Florida | Nov 1997 | ♂ = 3-27 ♀ = 3-30 | ♂ = 440 ♀ = 487 | 0.000731 0.000865 | 3.024 2.951 | |||
| (McCoy, 1972) | Pink shrimp | North Carolina | Oct 1969-Jul 1972 | 15-40 | ♂ = 297 ♀ = 503 | 0.00148 0.00209 | 2.77 2.66 | |||
| (McCoy, 1972) | Brown shrimp | North Carolina | Oct 1969-Jul 1972 | ♂ = 12-40 ♀ = 12-40 | ♂ = 259 ♀ = 243 | 0.000819 0.00113 | 2.94 2.84 | |||
Los coeficientes a y b son los parámetros de la función P = aLb, n = tamaño de muestra. Tomado de (Diaz et al., 2001).
Las relaciones entre el peso (P) y la longitud del cefalotórax (LC) o la longitud total (LT) observadas en el presente estudio así como las relaciones de longitud del cefalotórax para otras especies de camarones peneidos según (Diaz, Smith, Serafy, & Ault, 2001) se presentan en la tabla 3.
Los resultados del promedio del LC para cada uno de los tres sitios en cada mes se muestran en la figura 3 (Fig. 3). No hay una variación marcada en el tamaño medio de los camarones indefinidos durante el año, pero hay un aumento notable en la LC del grupo de M-H en Punta Gorda desde el final del año hasta los primeros tres meses del próximo. Se observaron patrones similares cuando se examinó LT debido a la relación directa entre LC y LT (Fig. 4).
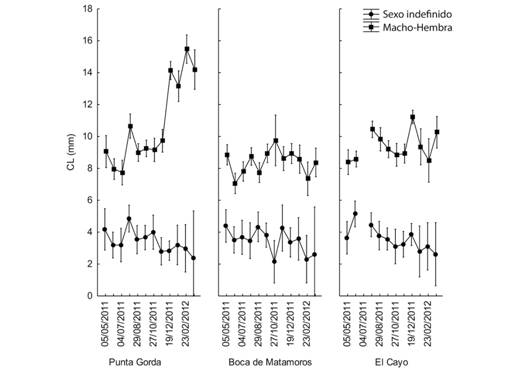
Fig. 3. Fig. 3 Longitud media del cefalotórax en cada sitio según el sexo y fechas de muestreo. Las barras verticales indican intervalos de confianza al 95 %. Average length of the cephalothorax in each site according to sex and sampling dates. The vertical bars indicate 95 % confidence intervals.
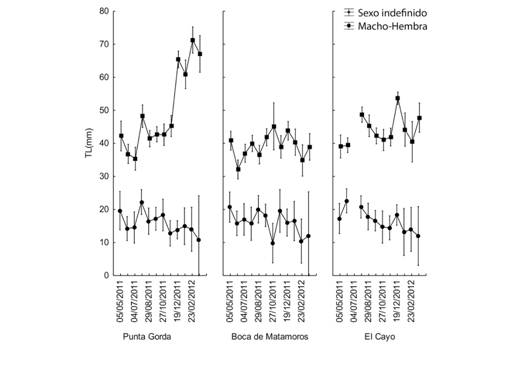
Fig. 4. Fig. 4 Longitud media total en cada sitio según el sexo y fechas de muestreo. Las barras verticales indican intervalos de confianza al 95 %. Total average length in each site according to sex and sampling dates. The vertical bars indicate 95 % confidence intervals.
Como no se encontró bibliografía sobre cual de los valores de Kn (Kn LC o Kn LT) sería mejor para determinar el factor de condición relativa y con ello describir el estado de los camarones, se calcularon tanto KnLC como KnLT (Fig. 5, Fig. 6). Los resultados del ANOVA factorial teniendo en cuenta los sitios, la fecha, el sexo y su interacción se presentan en la tabla 4 y tabla 5.
TABLA 4 TABLE 4 ANOVA factorial de KnLC para sitios, fechas, sexo y su interacción KnLC factorial ANOVA for sites, dates, sex and their interaction
| SS | DF | MS | F | p | |
| Intercepto | 0 | ||||
| Sitio | 0.0048 | 1 | 0.004767 | 0.04686 | 0.828628 |
| Fecha | 23.6353 | 10 | 2.363526 | 23.23533 | 0.000000 |
| Sitio × Fecha | 19.2815 | 21 | 0.918165 | 9.02628 | 0.000000 |
| Sitio × Sexo | 0.0019 | 1 | 0.001886 | 0.01854 | 0.891691 |
| Fecha × Sexo | 9.9780 | 10 | 0.997796 | 9.80912 | 0.000000 |
| Sitio × Fecha × Sexo | 9.8709 | 21 | 0.470044 | 4.62090 | 0.000000 |
| Error | 398.9507 | 3 922 | 0.101721 |
TABLA 5 TABLE 5 ANOVA factorial de KnLT para sitios, fechas, sexo y su interacción KnLT factorial ANOVA for sites, dates, sex and their interaction
| SS | DF | MS | F | p | |
| Intercepto | 0 | ||||
| Sitio | 0.0386 | 1 | 0.038559 | 1.19566 | 0.274259 |
| Fecha | 13.0616 | 10 | 1.306162 | 40.50163 | 0.000000 |
| Sitio × Fecha | 10.0280 | 21 | 0.477524 | 14.80713 | 0.000000 |
| Sitio × Sexo | 0.0917 | 1 | 0.091699 | 2.84343 | 0.091828 |
| Fecha × Sexo | 4.1778 | 10 | 0.417780 | 12.95457 | 0.000000 |
| Sitio × Fecha × Sexo | 6.6903 | 21 | 0.318586 | 9.87875 | 0.000000 |
| Error | 126.4830 | 3 922 | 0.032250 |
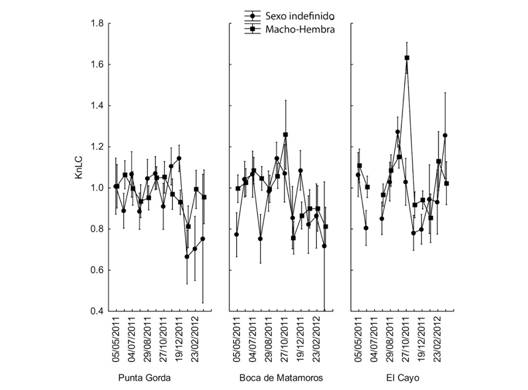
Fig. 5. Fig. 5 Comportamiento de KnCl en los sitios de muestreo durante el período de estudio. Las barras verticales indican intervalos de confianza al 95 %. KnCl behavior at the sampling sites during the study. The vertical bars indicate 95 % confidence intervals.
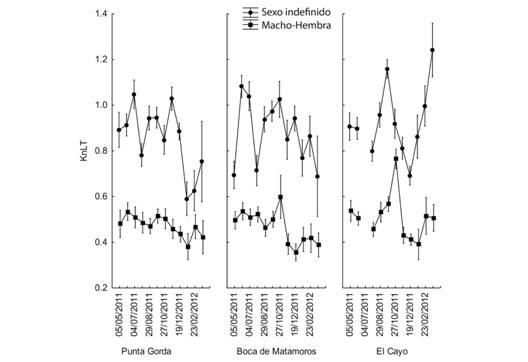
Fig. 6. Fig. 6 Comportamiento de KnLT en los sitios de muestreo durante el período de estudio. Las barras verticales indican intervalos de confianza al 95 %. KnLT behavior at the sampling
El comportamiento mostrado por ambos Kn es similar para los grupos I y M-H; no se observaron diferencias significativas entre los sitios de muestreo y la interacción sitios-sexo no fue significativa. Sin embargo, cuando se incluye la fecha en el análisis, las comparaciones mostraron diferencias significativas (Tabla 4 y Tabla 5).
El comportamiento de Kn LC en el gráfico para las dos categorías de sexo (I y M-H) en los diferentes sitios de muestreo es parecido y confuso para interpretar, mientras que los resultados son más claros cuando la longitud LT es empleada como dato para encontrar Kn. Por otra parte, se pueden observar diferencias considerables en la representación gráfica de ambos factores de condición evidenciándose factores de condición relativos mayores en los organismos pequeños o indefinidos (Fig. 5, Fig. 6).
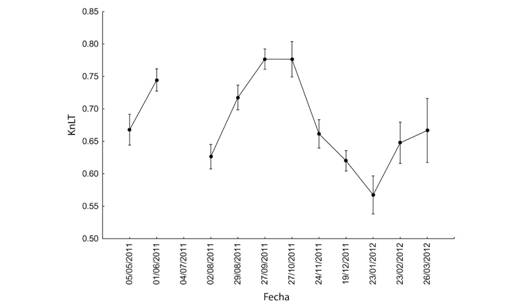
El comportamiento anual de KnLC y KnLT, involucrando a todos los organismos se muestra en las figuras 7 y 8 (Fig. 7, Fig. 8). Diferentes análisis no revelaron un patrón claro en el comportamiento de Kn, pero todos indicaron que hubo diferencias entre los meses muestreados (Tabla 4 y Tabla 5). Se observaron valores más altos en septiembre y octubre, lo que indica que durante ese período el peso medio estimado de la población para una talla dada está más cercano al peso real de los organismos y por lo tanto presentan una mejor condición. La diferencia entre KnLC y KnLT radica en que los valores de KnLT son de menor magnitud que los de KnLC lo cual puede interpretarse como que la Longitud del Cefalotórax es un mejor indicador del peso de los organismos que la Longitud Total o que la variación del peso del organismo está más asociada con el peso del abdomen.
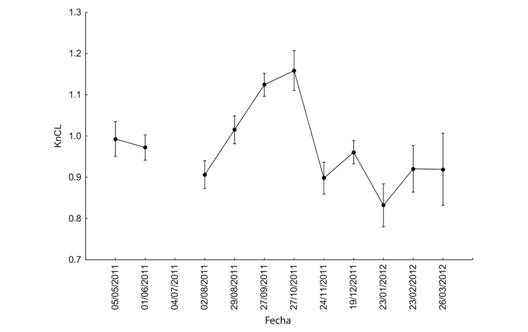
Fig. 7. Fig. 7 Comportamiento de KnLC durante el período de estudio. Las barras verticales indican intervalos de confianza al 95 %. KnLC behavior during the study. The vertical bars indicate 95 % confidence intervals.
Discusión
El valor de crecimiento alométrico positivo indica una mejora en la condición, altura o ancho que ocurre simultáneamente con el aumento en la longitud (Froese, 2006) y este es el caso de los organismos indefinidos en el presente estudio. (Diaz et al., 2001) encontraron crecimiento isométrico en machos de camarón rosado entre 3-27 LC mm e indicaron la existencia de crecimiento dimorfo, lo cual difiere con lo encontrado en el actual. El grupo M, el de H, así como el grupo combinado M-H exhibió un crecimiento alométrico negativo, lo cual coincide con los resultados para hembras de F. duorarum (3-30 mm LC) y para otras especies de camarones (Diaz et al., 2001).
Los resultados revelaron que los animales con mayores tallas de LC ≈ 14 mm, LT ≈ 65 mm (Fig. 4), se encontraban en Punta Gorda entre diciembre y marzo, sitio cercano a una boca de la laguna. Este comportamiento coincide con lo reportado por (Reyes et al., 1997) y pudiera entenderse como una acción en la que los camarones más grandes se están preparando para migrar hacia el mar abierto y completar su ciclo de vida. Los organismos con tallas menores, (≈ 4 mm LC) se encontraron distantes de dicho canal durante todo el año lo cual apoya dicha hipótesis. El comportamiento induce a pensar que, si el ecosistema en el área se ve afectado por causas naturales o antropogénicas, la consecuencia sobre el reclutamiento de juveniles a mar abierto podría ser severa.
Según (Gracia & Soto, 1990), las mayores capturas de juveniles de F. duorarum se obtienen en noviembre y por lo tanto están asociadas con el régimen de precipitación, el escurrimiento de los ríos del área y las fluctuaciones de salinidad. Aunque el oxígeno y el pH permanecieron estables durante el año, la temperatura y la salinidad variaron evidentemente ligados a este fenómeno. Para los tres sitios la salinidad se encontró en el rango de estudios previos (20-40 ups) (Yáñez-Arancibia & Lara-Domínguez, 1983) o (18.3-35.8 ups) (Guevara, Álvarez, Mascaró, Rosas, & Sánchez, 2007).
La similitud existente entre los parámetros abióticos registrados en los tres sitios permite explicar la similitud encontrada en los Kn obtenidos en las mismas fechas. Los valores mayores de Kn en los meses de septiembre y octubre coinciden con los registros de salinidad más bajos en el área. La reducción de la salinidad podría estar favoreciendo a la abundancia o de la diversidad en anfípodos, poliquetos, copépodos harpacticoides y detritus que forman parte de la dieta de F. duorarum (Lanza & Guzmán, 1993; Albertoni, Palma-Silva, & Esteves, 2003; Gutiérrez et al., 2016). Por otra parte, se considera que las diferencias en el tipo de sustrato y composición de los pastos marinos (Sánchez, 1997; Corona, Soto, & Sánchez, 2000; Brito, Gelabert, Ángel, Alderete, & Guevara, 2017) así como su fauna asociada podrían ayudar a explicar las diferencias encontradas en LC y LT en Punta Gorda con respecto a los otros dos sitios de muestreo. El comportamiento de los parámetros abióticos asociados con las fechas podría ayudar a explicar las diferencias en Kn cuando se incluye los meses en el análisis.
Los resultados muestran que los valores del índice de condición obtenidos usando LC están más cerca de 1 que los encontrados al usar la LT. Esta situación revela que LC es un mejor estimador para el peso de los camarones. Las diferencias en el factor de condición relativa cuando se usa el LT son más pronunciadas debido a la mayor variabilidad de las mediciones, por lo tanto, puede ser una mejor herramienta para evaluar las características de la población de camarones y debería incluirse como un índice para describir la dinámica de población en la población.
El Factor de condición relativo proporciona información sobre la condición del organismo y actúan como un indicador indirecto de la calidad del entorno. Mayores valores significan que el animal tiene mayores reservas y puede ser más resistente a los cambios en el ecosistema. Los valores del índice de condición pueden ofrecer información predictiva acerca de la calidad del hábitat y de la condición reproductiva, lo cual pueda apoyar a los tomadores de decisiones con las políticas sobre la conservación (Stevenson & Woods, 2006). Este índice podría explicar sexo, edad, madurez y estacionalidad en peces como Perca fluviatilis (Le Cren, 1951) e indica las condiciones propicias del ambiente (Samat, Shukor, Mazlan, Arshad, & Fatimah, 2008).
Los resultados con cerca de 4 000 organismos analizados durante todo un año pueden servir como referencia para estudios posteriores e indirectamente inferir el estado del ecosistema a través del factor de condición relativa. Organismos que se desarrollan en un ambiente con suficiente alimento y en condiciones ambientales adecuadas, ostentarán un mayor peso corporal y un Factor de condición relativo cercano a 1.
En los registros obtenidos se puede concluir que es necesario usar diferentes ecuaciones para expresar la relación entre LC y LT para animales indefinidos y con sexo evidenciado. Como era de esperar, la relación longitud total-longitud del cefalotórax (LT-LC) para cada categoría de sexo fue lineal, evidenciándose que la longitud del cefalotórax es un mejor estimador del peso que la longitud total de los organismos. Se encontró además que el grupo M, el de H, así como el grupo combinado M-H exhibieron un crecimiento alométrico negativo, mientras que los organismos clasificados como indefinidos presentaron crecimiento alométrico positivo. Por otra parte, se considera que el factor de condición relativa obtenido cuando se emplea la longitud total es un mejor indicador de las condiciones de las poblaciones de las postlarvas epibénticas en la región y pudiera contribuir a describir la dinámica de estas poblaciones.
Los resultados descritos para las postlarvas epibentónicas y los juveniles de F. duorarum contribuyen a mejorar el conocimiento sobre la especie en la Laguna de Términos y el Golfo de México. Esta información podría usarse como una referencia para contrastar los resultados de otros estudios y establecer una línea base que pueda ser utilizada en estudios de restauración ecológica del ecosistema.

