











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
The genus Platymantis is highly diverse, with 45 known species found in the Philippines, Palau, Fiji, New Guinea, and in the Admiralty, Bismarck, and Solomon Islands (Brown, Siler, Richards, Diesmos, & Cannatella, 2015; amphibiaweb.org). In the Philippines, dozens of newly discovered species, including many members of Platymantis, await formal description. Diesmos, Alcala, Siler, and Brown (2014) recently summarized many of these discoveries (e.g., P. biak Siler, Diesmos, Linkem, Diesmos, and Brown, 2010 from the karst forest of central Luzon) and a new phylogenetic hypothesis has been proposed for this group (Brown et al., 2015).
Platymantis is a direct developer (i.e., without tadpole stage). Alcala, Alcala, and Dolino (2004) and Alcala and Alcala (2005) studied the population densities of lowland forest herpetofauna, with emphasis on threatened species such as Platymantis spelaeus Brown & Alcala, 1982 (endemic to the karst habitats of Southwestern Negros) and P. insulatus Brown & Alcala, 1970, endemic to small islands (Islas de Gigantes) off Northern Iloilo. The most recent ecological study involving Platymantis and the importance of their microhabitats as “thermal buffers” included the series of papers by Scheffers, Brunner, Ramirez, Shoo, Diesmos, and Williams (2013a) and Scheffers, Phillips, Laurance, Sodhi, Diesmos, and Williams (2014) on Mt. Banahaw, Luzon Island. These works demonstrated the very specific arboreal microhabitat use of Platymantis montanus (Taylor, 1922), a species demonstrated to be a very close relative of P. hazelae (Brown et al. 2015). Alcala (1962) conducted extensive observations on the breeding behaviour of Philippine frogs (including Platymantis hazelae Taylor, 1920), including life history and microhabitat use (e.g. leaf axils of screw pines), as well as oviposition sites; he also provided morphological description of the species’ embryos and froglets. P. hazelae has been known as nocturnal, sexually dimorphic (males slightly smaller, 21.8-28.2 mm SVL, with vocal sacs; females 26.2-5.2 mm SVL), and juveniles below 20 mm SVL (Alcala, 1962).
The importance of phytotelmata (water bodies held by terrestrial plants) has been emphasized by a number of studies (Alcala, 1962; Ferreira, Schineider, & Teixeira, 2012; Kitching, 2000; Lehtinen, 2002; Scheffers et al. 2013a, 2014; Domingos, Arantes, Cavalcanti, & Jotta, 2015). A number of Philippine frog species have been documented to utilize leaf axils of screwpines (Pandanus) as breeding sites, such as P. hazelae and P. negrosensis (Alcala, 1962) and Pleophryne brevipes (Alcala & Brown, 1982; Denzer, 1994). Scheffers et al. (2013a) described the importance of phytotelmata to act as thermal buffers during extreme climatic conditions. Frogs that are dependent on phytotelmata may be otherwise subject to desiccation, contributing to Alcala, Bucol, Diesmos, and Brown (2012) classification of these species as “highly vulnerable”. The goal of this paper was to describe the abundance and spatio-temporal distribution of P. hazelae based on a six-month survey during the rainy season (September 2016 to February 2017) in three forest zones (montane, sub-montane, and lowland). We also described some aspects of the species’ breeding behaviour and characters of embryos and developing froglets.
Materials and methods
Study site: The forests surrounding the Twin Lakes of Balinsasayao (Fig. 1) and Danao in Negros Oriental, Philippines, form part of an extensive tropical rainforest ecosystem within the 133 000 half the geothermal reserve of the Energy Development Corporation (formerly Philippine National Oil Company, PNOC). The area has an elevation range of ~650-900 m.a.s.l. near the lakes and extends to about 1 750 m.a.s.l. at the highest peak known as “Guinsayawan” towards the Southwest. A dome-shaped peak known as “Guintabon” is adjacent to the lakes in the North. A third, much smaller water body, known as “Kabalin-an” Lake, located Northwest of Guintabon has a maximum water surface area reaching ~3.4 ha in 2014 but reduced to only ~1.3 has during low rainfall months, notably right after the El Niño phenomenon in 2010.
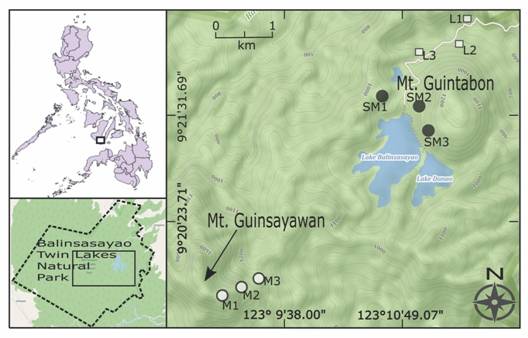
Fig. 1 Location of the study sites within Balinsasayao Twin Lakes Natural Park (montane sites indicated by M1-M3, submontane sites by SM1-SM3, and lowland sites by L1-L3). Inset: Philippine map and the boundaries of Balinsasayao Twin Lakes Natural Park. Map generated using QGIS® and OpenStreetMap®.
Three study zones were pre-determined based on the general description by Brown and Alcala (1961). The geographic coordinates of selected study zones are as follows, montane zone (9°22'1.68" N & 123°10'54.66" E), submontane zone (9°22'7.85" N & 123°11'9.92" E), and lowland zone (9°22'20.78" N & 123°11'19.00" E). In each zone, three sites were selected (where possible with minimal signs of human activities) and marked using a GPS unit. In each site, three replicate plots of 20 x 20 m were established, each corner marked with a flagging tape. The montane zone (> 1 300 m.a.s.l.) included in this study is located at the Guinsayawan peak (Fig. 2A). This site requires ~4-5 h of uphill climb along the trails at the top of ridges. The forest at this zone is typically composed of trees growing on steep slopes mixed with laurels (Lauraceae) and other small to medium sized trees, and in most locations, the presence of screw pines (Pandanus) and Heterospathe palms. Although the trail continues towards the summit of Guinsayawan, sampling at > 1 300 m elevation was not undertaken, due to safety concerns. All plots in this zone were marked from the trail (top of the ridge) towards the Northwest side of the terrain, which has a gentler slope compared to the opposite side of the ridge. The submontane zone is located at the foot of Mt. Guintabon. The slope in this site is gentle but is characterized by the presence of large volcanic boulders (Fig. 2B). The three selected sites in this zone are 200 to 400 m from each other. Large trees with diameters at breast height of > 30 cm and height of > 20 m include the “almaciga” (Agathis philippensis) while the remaining vegetation is comprised of smalls trees (e.g. Lauraceae), tree ferns (e.g. Sphaeropteris glauca and Cyathea negrosiana), abaca (Musa textilis), and wild banana or “pakol” (Musa sp.). The lowland zone is characterized by steep slope (> 30°) with native forest trees growing on steep ravines and slopes (Fig. 2C). The slope in this zone is gentle and characterized by the presence of large volcanic boulders. Conspicuous trees in this site are Acacia mangium (a reforested species), and a combination of patches of native trees (e.g. Shorea spp.), abaca (Musa textilis), and Ficus species (Moraceae) growing on large boulders (~2 m in height).

Fig. 2 Forest zones: A) montane, B) submontane, C) lowland) in Balinsasayao Twin Lakes Natural Park, Negros Island, Philippines.
Frog abundance and distribution: Upon establishment of sample plots, a team of six people carefully searched for individuals of Platymantis hazelae, with efforts to minimize disturbance to frog microhabitats. Four people were assigned to go into each plot to search, count, and capture frogs; the other two researchers remained outside the plots (Dodd, 2016) to record temperature and relative humidity data and receive captured frogs. To avoid counting the same frog, all collected frogs were placed in plastic bags and after counting and examining them (sexing was made seeing males vocal sacs and the size, females are generally larger and age class, juvenile or adult SVL < 20 mm considered juveniles), all were released back to their original microhabitats.
Because Platymantis hazelae is nocturnal (Alcala, 1962), sampling was undertaken on a monthly basis, from 5-11pm (5-10 nights/month). Adult males are easily distinguished by their distinct advertisement calls. It should be pointed out that in Platymantis, only male frogs produce advertisement calls and that females lack vocal sacs (Alcala & Brown, 1998). Additionally, each species has a distinct male call, which has been used by frog taxonomists as a source of diagnostic characters (Brown & Diesmos, 2009; Diesmos, Brown, Alcala, Sison, Afuang, & Gee, 2002; Siler, Linkhem, Diesmos, & Alcala, 2007).
Environmental variables: Within each plot we measured 14 environmental variables, tree density, tree DBH, canopy cover, leaf litter mass, leaf litter depth, amount of rainfall, air temperature, fern height, fern density, relative humidity, altitude, tree height, density of palms, and screw pines (Table 1). Tree height was visually estimated by a trained research staff member while diameter at breast height (DBH) of each tree was measured using a tape measure. Tree density was expressed as the total count of individual trees (> 5 cm DBH) per plot and canopy cover was assessed using Ocular (Visual) Estimation Method, expressed in percent cover. Elevation was determined using a hand-held GPS unit (Garmin). Rainfall was monitored per site weekly or right after heavy rain downpour by a local assistant using an improvised rain gauge (10 cm wide cylinder). Air temperature and relative humidity (RH) as well as temperature and RH of each microhabitat during each frog survey within plots were taken using a digital hygrometer (model #HC 520) with an external temperature probe. Air temperature was measured using data loggers (HOBO Temp/Light pendant logger, UA-002-64) deployed in each sampling site. Data loggers were launched using the accompanying HOBO software through ONSET Universal Optic-USB Base Station (Model, BASE-U-4). Data logging was set to record every 80 min interval for about 140 d period.
Data processing and analyses: To estimate abundance, count data were expressed as individuals per plot (ind./ha), is also presented for easier comparison with other studies done in the study area (e.g. Brown & Alcala, 1961; Amoroso, Alcala, & Alcala, 2014 unpublished report). Because this study surveyed permanent plots throughout the survey period (6 months), a two-way Analysis of Variance (ANOVA) with repeated measures was used. Using nortest and lmtest packages in R (R Core Team, 2015), tests were performed to determine normality of data (Anderson-Darling test and Q-plot), normality of residuals (plotted studentized residuals), as well as homoscedasticity (Breusch-Pagan test) and presence of outliers. Values were log(x+1)-transformed prior to analysis to meet parametric assumptions of ANOVA.
The relationship between abundance of P. hazelae and the environmental variables was visualized using the Principal Component Analysis (PCA). The use of PCA was necessary to address the multicollinearity (i.e., high variance inflation factor > 5) detected among the predictor variables (Domingos, Arantes, Cavalcanti, & Jotta, 2015). PCA plot was generated using the ggbiplot package in R platform (R Core Team, 2015). Interpretation of PCA was based on eigenvalues and projected inertia (%) using the ade4 package (Dray & Dufour 2007). Prior to multicollinearity test and subsequently PCA, data were log(x+1) transformed to address the presence of zeros (e.g. zero count of frog) in the dataset (Zar, 1999).
Results
Abundance and spatio-temporal distribution: Platymantis hazelae was found only in the submontane and montane zones (Fig. 3). The species was not found in the lowland sites, despite intensive search efforts over six months. In the submontane zone, the species was limited to only one site (Site 1) with densities ranging from 2.78-13.89 ind./ha. In the montane zone, the species was consistently encountered at 30.56-86.11 ind./ha (±15.65 SE; N = 56). Most individuals of Platymantis hazelae found were males, except in November (montane zone), December, and February (submontane zone) (Fig. 4). Juveniles of P. hazelae were observed only in November (montane zone) although outside of the study plots, eggs and embryos of this species were found in Pandanus leaf axils along the trail going to Guinsayawan (montane zone). Females of P. hazelae were rarely found, often only detected when pairs were found together.
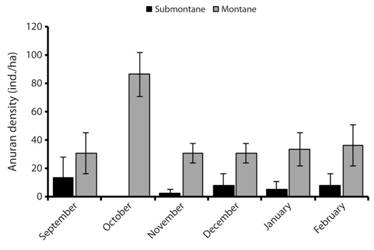
Fig. 3 Density of Platymantis hazelae in three zones from September 2016 to February 2017 (absent in lowland sites) in Balinsasayao Twin Lakes Natural Park, Negros Island, Philippines.
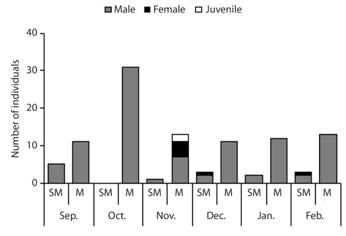
Fig. 4 Abundance of Platymantis hazelae according to sex classes in submontane (SM) and montane (M) zones from September 2016 to February 2017 (absent in lowland sites) in Balinsasayao Twin Lakes Natural Park, Negros Island, Philippines.
Frog abundance and distribution: Table 1 presents the ranges of environmental variables measured during the study (Table 1). When subjected to principal component analysis (PCA), among the 14 environmental variables, rainfall, altitude, relative humidity, were positively correlated with density of P. hazelae (Fig. 5). Similarly, Two-Way ANOVA with repeated measures over six months on relative humidity (RH) indicated a P < 0.05 for zone, indicating that zone significantly affect local humidity. However, month did have a significant effect (d.f. = 5, F = 15, P < 0.001), which indicates that the humidity levels recorded at various months are significantly different (SS = 459.72, df = 5, F = 15, P < 0.001). In addition, we detected a significant interaction between zone and month (SS = 271.32, d.f. = 10, F = 27.13, P < 0.001), suggesting that the relationship between month and RH depends on the identity of the area (zone) sampled (Table 2). Air temperature appears negatively correlated with the above-mentioned variables. A Two-Way ANOVA with repeated measures, over six months, with air temperatures as the dependent variable indicated no significant effect of zone (P > 0.05), indicating that zones did not have different air temperatures. Month of sampling did have a significant effect (SS = 41.42, d.f. = 5, F = 11.84, P < 0.001), which indicates that statistically detectable air temperatures varied monthly. We also found a significant interaction effect between zone and month (SS = 25.13, d.f. = 10, F = 2.73, P < 0.001), suggesting that the air temperatures differentially vary in response to the source of variation studied here (area [zone] versus date of sampling effort [month]).
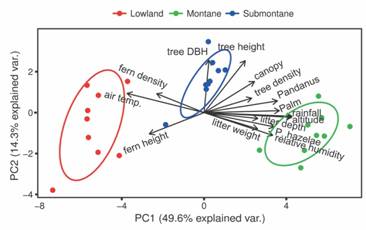
Fig. 5 PCA plot showing main two principal components (PC1, PC2) and biplots showing the trajectories of environmental variables in relation to density of Platymantis hazelae in Balinsasayao Twin Lakes Natural Park, Negros Island, Philippines.
TABLE 1 Average values of 14 environmental variables measured in each site in the three forest zones. A, B, and C are sites for each zone
| Environmental variables | Lowland | Submontane | Montane | ||||||
| A | B | C | A | B | C | A | B | C | |
| 1. Air temperature (°C) | 22.60 | 21.94 | 21.74 | 21.26 | 21.59 | 22.08 | 18.96 | 18.51 | 18.38 |
| 2. Relative humidity (%) | 87.58 | 90.85 | 86.96 | 87.52 | 89.93 | 88.80 | 95.21 | 96.59 | 94.22 |
| 3. Elevation (m.a.s.l.) | 655 | 700 | 780 | 953 | 930 | 900 | 1350 | 1330 | 1300 |
| 4. Rainfall (mm/hr) | 0.49 | 0.5 | 0.47 | 0.73 | 0.69 | 0.72 | 3.17 | 3.15 | 2.8 |
| 5. Leaf litter wt. (kg) | 1.42 | 0.50 | 0.79 | 0.99 | 1.65 | 1.14 | 1.39 | 1.73 | 1.42 |
| 6. Leaf litter depth (cm) | 2.33 | 2.11 | 1.74 | 2.83 | 2.42 | 3.03 | 2.89 | 3.25 | 2.67 |
| 7. Tree DBH (cm) | 10.26 | 19.46 | 23.44 | 21.89 | 17.98 | 19.10 | 20.76 | 14.07 | 15.73 |
| 8. Tree density (ind./400m2) | 21.67 | 11.67 | 10.00 | 35.00 | 62.33 | 62.00 | 43.00 | 29.33 | 29.33 |
| 9. Tree height (m) | 8.06 | 11.86 | 15.03 | 14.57 | 16.70 | 16.53 | 14.85 | 12.95 | 15.03 |
| 10. Fern height (cm) | 88.57 | 59.36 | 41.04 | 63.78 | 36.28 | 23.84 | 21.67 | 26.67 | 47.50 |
| 11. Canopy cover (%) | 28.33 | 31.67 | 41.67 | 73.33 | 83.33 | 85.67 | 50.50 | 65.00 | 65.00 |
| 12. Pandanus (ind./400m2) | 1.00 | 0.00 | 0.67 | 15.00 | 7.00 | 5.00 | 10.00 | 8.33 | 32.33 |
| 13. Fern density (fronds/m2) | 21.00 | 13.33 | 15.33 | 34.00 | 20.33 | 15.33 | 4.00 | 8.00 | 10.10 |
| 14. Palm (ind./400m2) | 1.67 | 0.00 | 0.67 | 10.00 | 4.33 | 9.00 | 18.00 | 26.67 | 33.33 |
Elevation: Lowland (655-780 m.a.s.l.); Submontane (900-953 m.a.s.l.); and Montane (1300-1350 m.a.s.l.)
TABLE 2 Summary of Two-way ANOVA with repeated measures on ambient temperature and relative humidity
| Air temperature (°C) | Sum of sqrs | df | Mean square | F | p-value | significance |
| Zone: | 128.714 | 2 | 64.357 | 0.071 | 0.931 | n.s. |
| Month: | 41.416 | 5 | 8.283 | 11.84 | 1.90E-09 | *** |
| Zone x Month: | 25.127 | 10 | 2.513 | 2.727 | 0.003 | ** |
| Zone x subj: | 46 809.1 | 52 | 900.174 | |||
| Month x subj: | 90.943 | 130 | 0.699 | |||
| Zone x Month x sub: | 239.592 | 260 | 0.922 | |||
| Total: | 824.176 | 161 | ||||
| Relative humidity (%) | ||||||
| Zone: | 545.283 | 2 | 272.641 | 1.59E-02 | 0.984 | n.s. |
| Month: | 459.715 | 5 | 91.943 | 15 | 1.30E-11 | *** |
| Zone x Month: | 271.318 | 10 | 27.132 | 3.146 | 0.00081 | *** |
| Zone x subj: | 892 261 | 52 | 17 158.9 | |||
| Month x subj: | 796.995 | 130 | 6.131 | |||
| Zone x Month x sub: | 2 242.1 | 260 | 8.624 | |||
| Total: | 5 930.56 | 161 |
Our temperature data (late September 2016 to early February 2017) includes 6 843 observations (~2 300 each; Fig. 6) taken at 80 min interval for about 140 days. Lowland air temperatures ranged from 18.71 to 29.85 °C (mean = 22.31 °C) and submontane forest ranged from 17.28 to 27.96 °C (mean = 20.65 °C). As expected, among the three zones, high elevation montane forest exhibited the lowest temperature ranging from 14.61 to 20.42 °C (mean = 18.71 °C). Variation in air temperature and relative humidity (Fig. 7, Fig. 8) indicate that mean air temperatures ranged from 16.6±0.2 to 23.7±0.6 °C, while mean RH ranged from 83.3 to 97.9 %. These data indicate the uniqueness of both lowland and montane sites demonstrate that the submontane site was intermediate in ranges of variation detected over our 140 day sampling effort. With increasing elevation, we note a decline in air temperature and the submontane zone exhibited wide variance in temperature, which contrasted sharply with the montane zone, which consistently remained at or slightly below 20 °C.
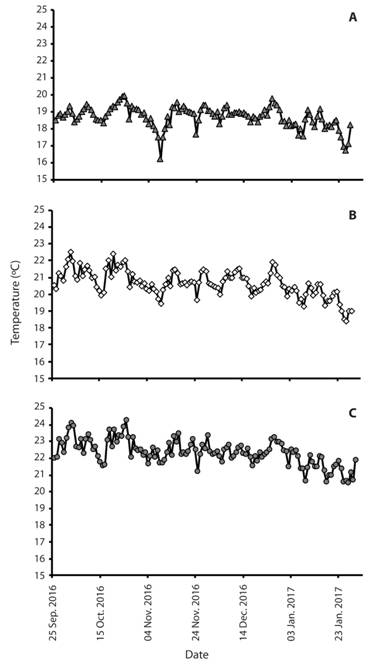
Fig. 6 Daily temperature readings using HOBO data loggers in three zones A) montane, B) submontane, C) lowland from September 23, 2016 to January 29, 2017 in Balinsasayao Twin Lakes Natural Park, Negros Island, Philippines.
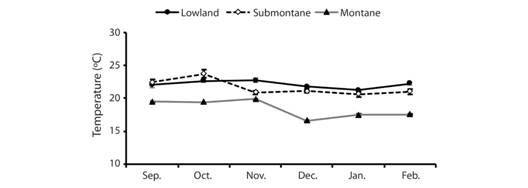
Fig. 7 Monthly temperature readings using hygrometer in three zones A) montane, B) submontane, C) lowland from September 2016 to February 2017 in Balinsasayao Twin Lakes Natural Park, Negros Island, Philippines.
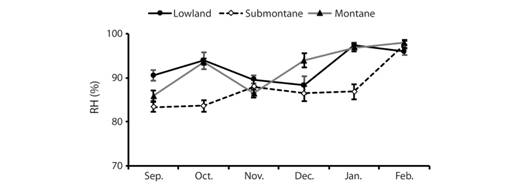
Fig. 8 Monthly relative humidity readings using hygrometer in three zones A) montane, B) submontane, C)lowland) from September 2016 to February 2017 in Balinsasayao Twin Lakes Natural Park, Negros Island, Philippines.
The number of individuals (total counts for each zone) varied among months (10-64 ind.), with highest counts either in submontane or montane zones (Fig. 9). Platymantis hazelae was restricted to the submontane and montane zones, but this species has been known take place > 200 m.a.s.l. within its known range. As shown in figure 8, the highest count per site was recorded in the montane zone during month of high rainfall (Fig. 8). Rainfall was unaffected by zone (P = 0.227), but was significantly influenced seasonally. The interaction effect between these two variables was significant (SS = 18.5, d.f. = 10, F = 3.871, P < 0.001), suggesting that the relationship between month and rainfall depends on the zone sampled (Table 3). Average monthly rainfall ranged from 0.28-5.69 mm/hr, peaks of which were recorded in the montane zone from October to December 2016.
TABLE 3 Summary of Two-Way ANOVA with repeated measures on rainfall from September 2016 to February 2017 measured at each site
| Rainfall | Sum of sqrs | df | Mean Square | F | p-value | significance |
| Zone: | 19.241 | 2 | 9.620 | 1.627 | 0.227 | n.s. |
| Month: | 8.627 | 5 | 1.725 | 3.496 | 0.010 | * |
| Zone x Month: | 18.500 | 10 | 1.850 | 3.871 | 0.0003 | ** |
| Zone x subj: | 94.590 | 16 | 5.9119 | |||
| Month x subj: | 19.739 | 40 | 0.493 | |||
| Zone x Month x subj: | 38.230 | 80 | 0.478 | |||
| Total: | 143.87 | 53 |
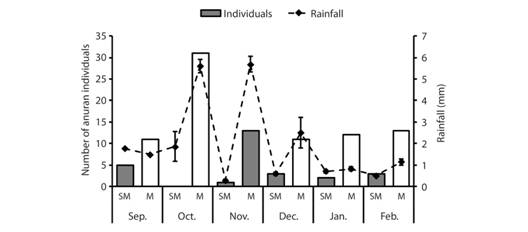
Fig. 9 Abundance of Platymantis hazelae superimposed with rainfall data from September 2016 to February 2017 (absent in lowland sites), SM-submontane, M-montane.
Microhabitat use: Platymantis hazelae was found mainly in arboreal microhabitats, ranging from ~0.5 m to 5 m above the ground. Of the 64 individuals observed, 51.56 % were found in leaf axils and surfaces of screw pines (Pandanus spp.), while 14.06 % were found on surfaces of palm fronds (Heterospathe sp.), 10.94 % were found on tree ferns (e.g. Cyathea negrosiana), 3 % were found on Musa (wild bananas, abaca) leaf axil/leaves, 4.69 % were found among low-hanging (0.5-1 m above the ground) tree leaves, and 4.69 % were found on tree buttresses, and 10.94 % were found on shrubs (Fig. 10). Platymantis hazelae, like all Philippine Shrub Frogs is mainly arboreal, but was occasionally found on lower vegetation strata, occasionally in the same microhabitats as P. dorsalis, a terrestrial species, which appears scansorial in the montane zone. In this study, P. hazelae, was most frequently encountered in their preferred microhabitats (leaf surfaces and axils of Pandanus), except during periods of high precipitation, especially in the montane zone, where they were found more frequently among ground vegetation (erect fern fronds), from near ground level to ~5 m above the ground.
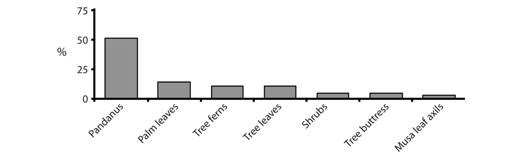
Fig. 10 Microhabitat use (%) of Platymantis hazelae (N = 64 individuals) in Balinsasayao Twin Lakes Natural Park, Negros Island, Philippines.
Oviposition and froglets: During surveys performed on January 20th, 2017 (~7 pm), a male calling from a hanging coiled abaca (Musa textiles) leaf was documented by our local field assistant (E. Zosima) ~0.5 m above the steep slope in the submontane sampling site. Upon inspection, the individual was sitting on top of an egg clutch (Fig. 11), containing six developing embryos, with completely developed operculum and vascularized abdominal sacs, indicating that the clutch was approximately 30 days old (Alcala, 1962). It is noteworthy that all clutches were attached on vertical or inclined surfaces that never accumulate excess water (Fig. 11A). More study, however, is needed to confirm our initial observations. Eyes and abdominal buds were already conspicuous during this stage (Fig. 11, Fig. 12C, Fig. 12D). This clutch hatched on February 05-07, 2017 (Fig. 11E, Fig. 11F). Diurnal observations on an additional three egg clutches of varied developmental stages (found on January 29, 2017 at Mt. Guinsayawan), however, did not include male attendance of clutches. Along existing trails, we also checked for possible egg clutches laid in Pandanus leaf axils. In about 1km walk along the trail, we found two clutches of varying stages laid in inclined Pandanus leaf axils, one newly laid (about blastula stage, with 8 embryos) and another clutch with four embryos having well-developed abdominal sacs. Interestingly, one clutch found in November 2016 that was laid on top of a fallen log, about 1.5 m above the ground, remain undeveloped.
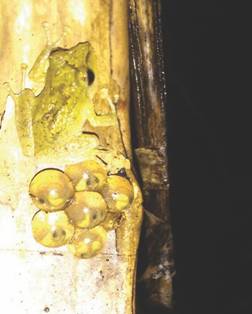
Fig. 11 A male Platymantis hazelae found near an egg clutch (Photo by E. Zosima: 20 January 2017 ~7:00 pm) in Balinsasayao Twin Lakes Natural Park, Negros Island, Philippines.
Developed embryos were observed moving their appendages while inside egg capsules. On February 05, 2017 the head of one of the froglets emerged (Fig. 12E) but remained motionless and on the following day was completely freed from the egg capsule (Fig. 12F). On February 07, 2017 all froglets were found outside of the egg capsules. Upon emerging from the egg capsule, these froglets had transparent integuments (Fig. 12G), especially on the ventral side and lower limbs. When provided access to standing water, they readily immersed themselves in petri dishes of water.

Fig. 12 Eggs and froglets of Platymantis hazelae hatching from egg capsules A) egg clutch found in Pandanus leaf axil; B) newly deposited eggs with fungal infestation; C) egg clutch with developing froglets attached to dried banana leaf; D) close-up of developing froglets in A; E) egg clutch with first hatched froglet with emerging head; F) hatched froglets; G)close-up of youngest froglet showing transparent integument; H)froglet 3-5 days after hatching; I) fully metamorphosed froglet perched on Pandanus leaf; J) juvenile Platymantis hazelae perched at night on palm leaf) in Balinsasayao Twin Lakes Natural Park, Negros Island, Philippines.
Metamorphosed froglets were cream colored with blackish patches on lateral body surfaces, and dorsum with longitudinal whitish line at mid-dorsal line from tip of snout to lumbar region and the rest of body covered with white speckles. Posterior to axillary region shows with dark blotch. Upon release, one fully metamorphosed froglet quickly jumped, attached itself on leaf surfaces and eventually concealed within a leaf axil of Pandanus. At Mt. Guinsayawan, we also observed fully metamorphosed froglets, either on Pandanus leaf surfaces or immersed in rainwater contained in leaf axils during daytime (Fig. 12I). During sampling in November 2016, juveniles (Fig. 12J) with reddish brown warts and overall yellowish coloration were also observed (probably foraging) at night on surfaces of palm and shrub leaves. These juveniles were found roughly 2-5 m away from calling males.
Discussion
It should be pointed out that unlike the previous studies (e.g. rapid assessments on frog abundance by Amoroso et al. (2014), which relied only on male calls), this study quantified frog abundance by intensive searches within repeatedly visited study plots. Our abundance data are based on subjects either captured or directly observed by investigators. We consider our data more comprehensive, compared to previous surveys involving only counts of males detected by advertisement calls and multiplied by the factor of 2 (assuming 1:1 sex ratio), which Alcala and Alcala (2005) suggested might result in overestimating the actual number of individuals.
Platymantis hazelae was found only in the submontane and montane zones and was not encountered in lowland sites, despite intensive search for 6 months. In the submontane zone, this species was limited to only one site (Site 1) with densities ranging from 2.8 to 13.9 ind./ha. In the montane zone, density was estimated at 30.6-86.1±15.65 ind./ha (Fig. 3). When compared with published data from half a century ago (Brown & Alcala, 1961), our estimates were 1.5-10 times lower than previous estimates. During that time period, the forest condition on Cuernos de Negros mountain was still in pristine condition (A. Alcala, personal communication, April 25, 2019). However, comparisons with our study and that of Brown and Alcala (1961) should be taken with caution because their study site was located about 10 km from our study area.
The distribution of P. hazelae in this study and its abundance in the three forest zones were found consistent with the findings by Brown and Alcala (1961). For example, with the combined quadrat and cruising surveys, Brown and Alcala (1961) observed P. hazelae was more abundant in the submontane and montane forest zones. Most of P. hazelae encountered in this study were males, except in November (montane zone), December, and February (submontane zone). Juveniles of P. hazelae were observed only in November (montane zone) although outside of the study plots, eggs and embryos of this species were found in Pandanus leaf axils along the trail going to Guinsayawan (montane zone). It appears that females of P. hazelae were rarely encountered, often detected when pairs were found. It is possible that the paucity of encounters of females may relate to later emergence or lower detectability of non-vocalizing subjects.
It should be noted that values for temperatures were slightly lower (14-31 °C) compared to the values reported in other mountain areas (habitats of phytotelmata-associated Platymantis species in the Philippines such as in Mt. Banahao, Luzon by Scheffers et al. (2013a). Most relative humidity values recorded in this study were larger than 75 %, different from the ones reported by Scheffers (2013) that detected a wider range (40-100 %) in Mt. Banahao. Rainfall has been shown to influence survivorship and activities (e.g., breeding, calling) of phytotelmata-associated frogs (Summers & McKeon, 2004). As expected, this study observed high densities of P. hazelae during months of high rainfall in montane and submontane habitats.
In Philippine mountains, screw pines, especially Pandanus, are most abundant at high elevation, from ~1 000 m to 2 000 m.a.s.l. (Brown & Alcala, 1961; Alcala, 1962) and we recorded densities ranging from 35 ind./400 m2. Despite their importance, understory vegetation such as Pandanus and palms are rarely reported or published. Lehtinen (2002) surveyed the herpetofauna (e.g. frogs and lizards) associated with Pandanus but did not quantify density or abundance of this plant except mentioning that a total of 1 700 plants in their study area of at least 457 ha in the coastal or littoral forest in Madagascar. Platymantis hazelae utilized mainly leaf axil and screwpines surfaces (Pandanus), usually below 3 m from the ground (but juveniles were observed closer to the ground; 0.1-0.8 m; and were often seen hiding in Pandanus axils containing water). These observations are consistent with those of Brown and Alcala (1961) and Alcala (1962). Lehtinen (2002) enumerated several species of amphibians and reptiles that utilize Pandanus as their microhabitats. As documented in this study, the abundance of P. hazelae appears correlated with the quantity (or availability) of arboreal (but lower stratum) habitats such as screwpines (Pandanus), areal ferns, wild abaca, and palms. Alcala (1962) observed the species' use of these microhabitats as breeding habitats.
Juveniles and developing embryos of P. hazelae were found during the months of October 2016 through February 2017, in contrast to an earlier study by Alcala (1962) in which embryos were found from March to July, suggesting a prolonged breeding period. Given the known development period of P. hazelae embryos of ~50 d (Alcala, 1962), the single clutch found in the submontane zone that successfully hatched on February 05-07, 2017 was probably laid in early to middle of December 2016, during which prolonged heavy rainfall was observed. In this study, four to eight embryos were observed per clutch, about the same size clutch reported by Alcala (1962).
This study described in detail variation in abundance, macrohabitat type, and microhabitat use in P. hazelae, based on a six-month survey (September 2016-February 2017) in three major forest zones (montane, submontane, and lowland) within the Twin Lakes Balinsasayao Natural Park. Apparently, this species is uncommon at low elevations, and was undetected despite intensive search during the duration of the survey. Its absence in the lowland might be attributed to absence of potential microhabitats such as screwpines (Pandanus) and palms. While we documented a male sitting on top of an egg clutch, suggesting male-guarding behaviour, there is a need to conduct more observations. Observations of its breeding behavior suggest an extended embryonic period. We also noted dependence of newly metamorphosed juveniles on phytotelmata microhabitats.

