











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
La expansión agrícola especialmente en los trópicos, afecta la riqueza y abundancia de plantas y animales de los sitios donde ocurre y sus alrededores (Foley et al., 2005; Foley et al., 2011), debido a la modificación de la cobertura natural y su remplazo por coberturas más homogéneas como pastizales o monocultivos (Ramankutty, Evan, Monfreda, & Foley, 2008). Por lo tanto, comprender como estos cambios afectan la abundancia y riqueza de las especies nativas, es imperativo si queremos ayudar a que las especies nativas permanezcan y utilicen estos paisajes modificados (Fischer et al., 2008; Vilchez Mendoza et al., 2014).
No todas las áreas agrícolas tienden a ser plantaciones extensivas y monocultivistas; pues en las regiones rurales cercanas a las ciudades existen pequeños desarrollos agrícolas que presentan una matriz más heterogénea de hábitats (Perfecto, Rice, Greenberg, & Van der Voort, 1996; Miles & Kapos, 2008). Por ejemplo, una mezcla de diferentes tipos de plantaciones (ej.: café, cítricos, pastos, caña de azúcar, o forestales), con vegetación natural remanente y regeneraciones naturales, cercas vivas y pequeños desarrollos urbano-rurales (Weibull, Bengtsson, & Nohlgren, 2000; Tscharntke et al., 2008). Esta heterogeneidad ambiental podría incrementar la complejidad de la composición y estructura de las comunidades de animales, al mantener características del hábitat previo, que pueden favorecer a especies nativas, así como crear nuevas condiciones de hábitat atractivas para especies colonizadoras (Wunderle & Latta, 1996; Greenberg, Bichier, & Sterling, 1997; Greenberg, Bichier, Cruz Agnon, & Reitsma, 1997).
Por el contrario, estos cambios en el uso de suelo y la ocurrencia de una mayor heterogeneidad ambiental también podrían influir negativamente en la abundancia de las especies nativas y positivamente en la abundancia de las especies colonizadoras (Stiles, 1990; Biamonte, Sandoval, Chacón, & Barrantes, 2011; Barrantes, Ocampo, Ramírez-Fernández, & Fuchs, 2016). Además, la reducción y fragmentación del hábitat natural puede reducir el tamaño de las poblaciones nativas, al incrementar el aislamiento entre poblaciones, lo cual limita el intercambio genético interpoblacional (Bierregaard & Lovejoy, 1988; Hughes, Daily, & Ehrlich, 1997; Manhães & Loures-Ribeiro, 2005). Otra consecuencia de la reducción del hábitat nativo es el aumento inicial de la densidad de las poblaciones, acelerando la transmisión de enfermedades o incrementando el número de interacciones agresivas entre los individuos (Costantini, Greives, Hau, & Partecke, 2014; Soulsbury & White, 2016). Por otro lado, la fragmentación aumenta el área de ocurrencia y promueve el flujo genético dentro de especies de zonas abiertas o con disturbios, al conectar poblaciones previamente aisladas (Meillère, Brischoux, Parenteau, & Angelier, 2015; Beninde et al., 2016).
Uno de los grupos taxonómicos donde la mayoría de especies dependientes de hábitats naturales se ven afectados por estos cambios de uso de suelo debido a la agricultura son las aves, ya que muchas especies están restringidas a un tipo particular de hábitat y si ese hábitat disminuye o se elimina, lo hacen las especies asociadas (Stiles, 1985; Stotz, Fitzpatrick, Parker, & Moskovits, 1996). Por esta razón la riqueza y abundancia de aves se han utilizado como un indicador del efecto producido por el cambio de uso de suelo, porque permiten comparar las comunidades de aves entre sitios con diferentes tipos de cobertura agrícola y natural (Tilghman, 1987; Tscharntke et al., 2008). No obstante, la mayoría de estudios previos buscaron relacionar la matriz agrícola con la riqueza de especies y su abundancia, pero los cambios en la riqueza y abundancia a través del año y entre años, han sido escasamente evaluados. Por lo tanto mi objetivo fue analizar la variación anual en la riqueza y abundancia de las aves que ocurre en un mosaico agrícola-rural a lo largo de un periodo de ocho años. Adicionalmente analicé los cambios en riqueza y abundancia de las especies tomando en cuenta el grado de dependencia por los hábitats (ej.: bosque o zonas abiertas), la dieta, y si son o no residentes en el país.
Materiales y métodos
Área de estudio y muestreo: El estudio lo realicé en Getsemaní, provincia de Heredia, Costa Rica (10°01’43” N, 84°06’44” W, 1300 msnm), En un área de aproximadamente 30 ha, donde: 40 % corresponde a cafetales (con sombra y sin sombra con cercas vivas), 40 % a pastizales para ganadería (con árboles aislados y cercas vivas), 10 % de vegetación en regeneración natural (incluyendo vegetación riparia), y 10 % de zonas urbano-rural (Fig. 1; Sandoval, 2011). Durante el periodo de estudio (diciembre 2002 a abril 2010), no se dio ningún cambio en el uso de suelo, salvo pequeñas podas de cafetales debido al mantenimiento, caída azarosa de árboles o arbustos debido a las lluvias o vientos fuertes, o muerte de plantas por su senescencia normal.
Para recolectar lo datos recorrí a velocidad moderada en promedio una vez al mes (2002 solo un muestreo y 2010 solo tres muestreos) un transecto de 3 km de longitud con bandas fijas de 25 m a cada lado, desde las 06:00 hasta las 09:00, que abarcaba la mayoría del área de estudio y pasaba por los cuatro tipos de uso de suelo del área (potreros, cafetales, regeneración natural y urbano rural; Fig. 1). Para evitar recontar individuos en sitios diferentes el punto más cercano entre dos sectores del transecto fueron 100 m. Además, alterné la dirección de inicio del muestreo entre conteos para aumentar la probabilidad de detección de los individuos en todos los hábitats y áreas de muestreo.
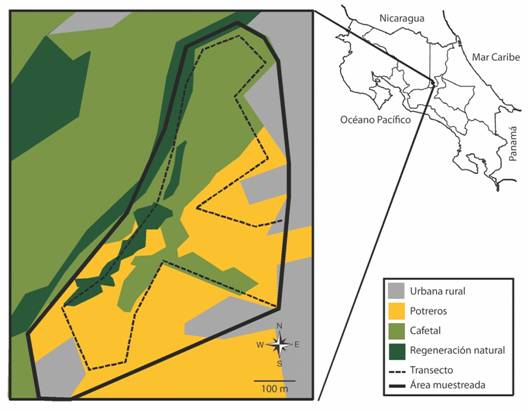
Fig. 1 Fig. 1 Área de estudio (Getsemaní, provincia de Heredia, Costa Rica) mostrando los cuatro usos de suelos principales y el transecto utilizado para muestrear las aves de 2002 a 2010. Mapa de uso de suelo creado con base en imágenes de Google Earth durante el periodo de estudio. En la esquina superior derecha el mapa de Costa Rica indicando la ubicación del sitio de estudio en el país. Four main land uses at the study area (Getsemaní, Heredia province, Costa Rica) with transect used on bird sample from 2002 to 2010. Land use was created using Google Earth images form study period. In the upper right corner is the Costa Rica map showing the location in the country.
Durante el recorrido, anoté la cantidad de individuos observados y escuchados por especie. Siguiendo la dirección de desplazamiento de las aves o bandadas, e intentando dentro de lo posible estimar si se dirigían a sectores aun no muestreados del transecto, evité contar los individuos de esas especies si se encontraban a menos ≤ 100 m de la primera observación para no sobreestimar la abundancia de individuos por especie. En especies que cazan y pasan la mayor parte del tiempo volando (ej.: golondrinas o zopilotes) anote el número máximo de individuos observados en cualquier momento del conteo, en lugar de ir adicionando los individuos observados (ej.: si observaba dos golondrinas al iniciar el conteo, cinco a la mitad, y dos al final el número de golondrinas anotado sería cinco, y no nueve) para evitar sobreestimaciones.
Todas las especies fueron clasificadas según su residencia en el país: residentes, si permanecen todo el año en la zona; migratorios altitudinales residentes, si tiene desplazamientos a lo largo de las vertientes de Costa Rica; migratorios norteños, si se reproducen en Norte América y pasan el invierno del hemisferio norte en Costa Rica; y migratorios sureños, si se reproducen en Costa Rica y migran hacia Sur América después de reproducirse (Stiles & Skutch, 1989). Además, clasifiqué las especies según el grado de dependencia del bosque siguiendo lo propuesto por Stiles (1985), y modificado por Sandoval & Barrantes (2009) y Biamonte et al. (2011): 1 = especies que necesitan bosques maduros para sobrevivir, 2 = especies que viven en áreas con al menos 50 % de cobertura boscosa, 3 = especies que viven en áreas abiertas; 4 = especies que viven en lagunas o humedales, y aquellas especies con valores 1-2 y 2-3 en Stiles (1985) fueron reclasificados como 1.5 y 2.5 respectivamente. Donde especies con valor de 1.5 son especies que viven tanto en bosques maduros y áreas con al menos 50 % de cobertura boscosa; y 2.5 son especies que viven tanto en áreas con al menos 50 % de cobertura boscosa y áreas abiertas.
Finalmente, según los criterios de Stiles y Skutch (1989) se clasificaron como: cazadores, si se alimentan de vertebrados o carroña; frugívoros grandes, si pesan > 20 g y se alimentan de frutas; frugívoros pequeños, si pesan ≤ 20 g y se alimentan de frutas; insectívoros, si se alimenta de insectos u artrópodos; nectarívoros, si se alimentan de néctar; semilleros, si se alimentan de semillas; y omnívoros, si su dieta está compuesta por dos o más tipos de alimentos en proporción similar.
Análisis estadísticos: Realicé un modelo lineal mixto (MLM; Neter, Kutner, Nachtsheim, & Wasserman, 1996) por categoría de residencia en el país para comparar como cambia la riqueza de especies (variable dependiente) entre años (variable independiente) y utilizando como variable aleatoria el mes. Para las categorías de residencia: todos (que incluye todas las especies), solo residentes y migratorios altitudinales, no utilicé los datos de 2002 (con muestreo únicamente en diciembre) ni del 2010 (con muestreos de enero a marzo). Sin embargo, para la categoría de migratorios norteños si utilicé los datos de todo los años, porque los periodos migratorios empiezan en agosto y terminan en mayo, siendo la variable independiente el periodo migratorio (ej.: 2002-2003, 2003-2004, y así sucesivamente hasta 2009-2010). Luego utilice otro MLM por categoría de residencia en el país para comparar como cambia la riqueza de especies (variable dependiente) entre meses (variable independiente) y utilizando como variable aleatoria el año. Para comparar si la abundancia y riqueza de todas las especies y las especies residentes por mes fue similar entre meses realicé un análisis de similitud (ANOSIM). En este caso usé como algoritmo de separación de los grupos la distancia de similitud del índice de Morisita, comparando la abundancia y riqueza de todas las especies y de las especies residentes por mes. Para dicho análisis, la variable de agrupación fue cada mes, por lo tanto un resultado significativo indica que al menos hay un mes donde la similitud de los muestreos dentro del mes en términos de la riqueza de especies y abundancia de cada especie fue mayor, que entre los muestreos de diferentes meses.
Finalmente, realicé un análisis lineal generalizados (ALG; con una distribución de Poisson) para cada tipo de dieta y cada grado de dependencia del bosque (agrupe las categorías 1 y 1.5 en una sola para este análisis, debido a que registré muy pocas especies) para comparar si la riqueza de especies (variable dependiente) cambia entre los meses (variable independiente). Para todos los MLM y los ALG que resultaron estadísticamente significativos realicé pruebas post hoc para determinar entre cuales de las variables independientes se encontraban las diferencias, y estas diferencias se mencionan en el texto. Para realizar los MLM y ALG utilicé el software JMP versión 7.0 (SAS Institute, Cary, NC, EEUU), y para los análisis de ANOSIM el software PAST versión 3.21 (Hammer, Harper, & Ryan, 2001).
Resultados
De 2002 a 2010 realicé un total de 89 muestreos en los cuales registré un total de 160 especies de aves en el área de estudio (Tabla 1): 59 migratorias (55 norteñas y cuatro sureñas) y 101 residentes (de las cuales 13 fueron migratorias altitudinales). La riqueza total de especies (MLM: F6,68 = 2.93, P = 0.01) y de especies residentes (F6,68 = 3.93, P = 0.002) varió entre años y en ambos casos fue menor durante 2004 (Tabla 2). Sin embargo, la riqueza de especies migratorias altitudinales (F5,20 = 1.46, P = 0.25), migratorias norteñas (F7,63 = 1.86, P = 0.09), y migratorias sureñas (F7,20 = 1.73, P = 0.16) fue similar entre todos años (Tabla 2).
TABLA 1 TABLE 1 Lista de aves muestreadas de 2002 a 2010 en Getsemaní, Heredia, Costa Rica, a lo largo de un transecto lineal de 3 km, dentro de una matriz agrícola rural de 30 ha Bird list at Getsemaní, Heredia, Costa Rica from 2002 to 2010 sampled using 3 km transect inside 30 ha of agricultural rural matrix
| Taxa | Abundancia | *Residencia | Dependencia del Bosque | Tipo de Dieta |
| Accipitridae | ||||
| Buteo albonotatus | 0.06 | R | 2 | Cazador |
| Buteo brachyurus | 0.07 | R | 1 | Cazador |
| Buteo plagiatus | 0.36 | R | 2 | Cazador |
| Buteo platypterus | 2.78 | MN | 3 | Cazador |
| Buteogallus anthracinus | 0.01 | R | 2 | Cazador |
| Elanoides forficatus | 0.03 | R | 2 | Cazador |
| Elanus leucurus | 0.19 | R | 3 | Cazador |
| Apodidae | ||||
| Chaetura pelagica | 0.02 | MN | 3 | Insectívoro |
| Chaetura vauxi | 0.73 | R | 3 | Insectívoro |
| Cypseloides niger | 0.79 | R | 3 | Insectívoro |
| Streptopocne rutilus | 0.02 | R | 3 | Insectívoro |
| Streptoprocne zonaris | 2.55 | R | 3 | Insectívoro |
| Ardeidae | ||||
| Bubulcus ibis | 2.72 | R | 3 | Omnívoro |
| Bombicillidae | ||||
| Bombycilla cedrorum | 0.69 | MN | 2.5 | Frugívoro pequeño |
| Caprimulgidae | ||||
| Chordeiles minor | 0.01 | MN | 3 | Insectívoro |
| Nyctidromus albicollis | 0.28 | R | 3 | Insectívoro |
| Cardinallidae | ||||
| Habia rubica | 0.49 | R | 1.5 | Frugívoro pequeño |
| Passerina cyanea | 0.11 | MN | 3 | Semillero |
| Pheucticus ludovicianus | 0.36 | MN | 3 | Frugívoro pequeño |
| Piranga bidentata | 0.37 | R | 2.5 | Frugívoro pequeño |
| Piranga olivacea | 0.28 | MN | 3 | Frugívoro pequeño |
| Piranga rubra | 0.48 | MN | 2.5 | Frugívoro pequeño |
| Cathartidae | ||||
| Cathartes aura | 1.29 | R | 3 | Cazador |
| Coragyps atratus | 2.62 | R | 3 | Cazador |
| Charadriidae | ||||
| Pluvialis dominica | 0.01 | MN | 4 | Cazador |
| Ciconiidae | ||||
| Mycteria americana | 0.03 | R | 4 | Cazador |
| Columbidae | ||||
| Columba livia | 0.96 | R | 3 | Semillero |
| Columbina inca | 2.65 | R | 3 | Semillero |
| Columbina passerina | 0.39 | R | 3 | Semillero |
| Columbina talpacoti | 0.02 | R | 3 | Semillero |
| Leptotila verreauxi | 8.49 | R | 2.5 | Semillero |
| Patagioenas fasciata | 0.21 | MA | 2 | Semillero |
| Patagioenas flavirostris | 3.53 | R | 3 | Semillero |
| Zenaida asiatica | 3.75 | R | 3 | Semillero |
| Corvidae | ||||
| Psilorhinus morio | 5.39 | R | 3 | Frugívoro grande |
| Cuculidae | ||||
| Coccyzus americanus | 0.01 | MN | 3 | Insectívoro |
| Crotophaga sulcirostris | 3.37 | R | 3 | Insectívoro |
| Piaya cayana | 1.61 | R | 2.5 | Insectívoro |
| Tapera naevia | 0.02 | R | 3 | Insectívoro |
| Emberizidae | ||||
| Atlapetes albinucha | 0.66 | R | 2.5 | Semillero |
| Chlorospingus ophthalmicus | 0.08 | MA | 2 | Frugívoro pequeño |
| Melozone cabanisi | 0.93 | R | 2.5 | Semillero |
| Melozone leucotis | 5.21 | R | 2.5 | Semillero |
| Zonotrichia capensis | 4.00 | R | 3 | Semillero |
| Fringillidae | ||||
| Chlorophonia callophrys | 0.47 | MA | 1.5 | Frugívoro pequeño |
| Euphonia elegantisima | 0.02 | MA | 2 | Frugívoro pequeño |
| Euphonia leuticapilla | 0.02 | R | 2.5 | Frugívoro pequeño |
| Hirundinidae | ||||
| Hirundo rustica | 0.93 | MN | 3 | Insectívoro |
| Petrochelidon pyrrhonota | 0.35 | MN | 3 | Insectívoro |
| Pygochelidon cyanoleuca | 15.03 | R | 3 | Insectívoro |
| Riparia riparia | 0.28 | MN | 3 | Insectívoro |
| Stelgidopteryx ruficollis | 0.12 | R | 3 | Insectívoro |
| Stelgidopteryx serripennis | 0.09 | R | 3 | Insectívoro |
| Icteridae | ||||
| Dives dives | 2.65 | 3 | Frugívoro pequeño | |
| Icterus galbula | 5.10 | R | 2.5 | Frugívoro pequeño |
| Icterus spurius | 0.06 | MN | 3 | Frugívoro pequeño |
| Molothrus aeneus | 0.67 | R | 3 | Frugívoro pequeño |
| Psarocolius montezuma | 0.01 | R | 3 | Frugívoro grande |
| Quiscalus mexicanus | 1.85 | R | 3 | Omnivoro |
| Sturnella magna | 1.66 | R | 3 | Semillero |
| Mimidae | ||||
| Dumetella carolinensis | 0.01 | MN | 3 | Frugívoro pequeño |
| Momotidae | ||||
| Momotus lessonii | 2.56 | R | 2.5 | Omnívoro |
| Odontophoridae | ||||
| Colinus leucopogon | 2.89 | R | 2.5 | Semillero |
| Dendrortyx leucoprys | 0.04 | R | 2 | Semillero |
| Parulidae | ||||
| Basileuterus rufifrons | 5.73 | R | 2 | Insectívoro |
| Cardellina pusilla | 0.83 | MN | 2.5 | Insectívoro |
| Geothlypis poliocephala | 0.03 | R | 3 | Insectívoro |
| Icteria virens | 0.01 | MN | 3 | Insectívoro |
| Mniotilta varia | 0.24 | MN | 2 | Insectívoro |
| Oporornis formosus | 0.02 | MN | 1 | Insectívoro |
| Oporornis philadelphia | 0.10 | MN | 2.5 | Insectívoro |
| Oreothlypis peregrina | 3.72 | MN | 2 | Insectívoro |
| Parkesia motacilla | 0.01 | MN | 3 | Insectívoro |
| Protonotaria citrea | 0.07 | MN | 2 | Insectívoro |
| Seiurus aurocapilla | 0.04 | MN | 2 | Insectívoro |
| Setophaga cerulea | 0.02 | MN | 2 | Insectívoro |
| Setophaga discolor | 0.01 | MN | 2.5 | Insectívoro |
| Setophaga fusca | 0.26 | MN | 2 | Insectívoro |
| Setophaga pensylvanica | 0.34 | MN | 2.5 | Insectívoro |
| Setophaga petechia | 0.31 | MN | 3 | Insectívoro |
| Setophaga pinus | 0.01 | MN | 2.5 | Insectívoro |
| Setophaga ruticilla | 0.04 | MN | 2.5 | Insectívoro |
| Setophaga townsendi | 0.01 | MN | 2.5 | Insectívoro |
| Setophaga virens | 0.01 | MN | 2 | Insectívoro |
| Vermivora chrysoptera | 0.02 | MN | 2.5 | Insectívoro |
| Vermivora chrysopteraXpinus | 0.03 | MN | 2.5 | Insectívoro |
| Wilsonia canadensis | 0.12 | MN | 2.5 | Insectívoro |
| Picidae | ||||
| Colaptes rubiginosus | 0.01 | R | 2 | Frugívoro pequeño |
| Dryocopus lineatus | 0.17 | R | 2.5 | Frugívoro grande |
| Melanerpes hoffmannii | 8.12 | R | 2.5 | Frugívoro pequeño |
| Pipridae | ||||
| Chiroxiphia linearis | 0.56 | R | 2 | Frugívoro pequeño |
| Psittacidae | ||||
| Amazona albifrons | 0.06 | R | 2 | Frugívoro grande |
| Brotogeris jugularis | 0.10 | R | 3 | Frugívoro pequeño |
| Pionus senilis | 0.28 | R | 2 | Frugívoro grande |
| Psittacara finschi | 18.46 | R | 3 | Frugívoro grande |
| Rallidae | ||||
| Aramides cajaneus | 0.01 | R | 2 | Omnívoro |
| Striigidae | ||||
| Glaucidium brasilianum | 0.29 | R | 2.5 | Cazador |
| Megascops choliba | 0.03 | R | 3 | Cazador |
| Thamnophilidae | ||||
| Thamnophilus doliatus | 0.01 | R | 2.5 | Insectívoro |
| Thraupidae | ||||
| Cyanerpes cyaneus | 0.01 | R | 2 | Frugívoro pequeño |
| Saltator atriceps | 0.01 | R | 3 | Frugívoro pequeño |
| Saltator coerulescens | 2.54 | R | 3 | Frugívoro pequeño |
| Saltator maximus | 0.24 | R | 3 | Frugívoro pequeño |
| Tangara larvata | 0.02 | R | 3 | Frugívoro pequeño |
| Thraupis episcopus | 12.88 | R | 3 | Frugívoro pequeño |
| Thraupis palmarum | 0.01 | R | 3 | Frugívoro pequeño |
| Tiaris olivacea | 0.10 | R | 3 | Semillero |
| Volatinia jacarina | 0.03 | R | 3 | Semillero |
| Tityridae | ||||
| Pachyramphus aglaiae | 0.02 | R | 2.5 | Frugívoro pequeño |
| Tityra semifasciata | 0.74 | R | 2.5 | Frugívoro pequeño |
| Trochilidae | ||||
| Amazilia saucerrottei | 0.39 | R | 2.5 | Nectarívoro |
| Amazilia tzacatl | 2.74 | R | 3 | Nectarívoro |
| Campylopterus hemileucurus | 0.07 | MA | 2 | Nectarívoro |
| Chlorostilbon canivetii | 0.10 | MA | 3 | Nectarívoro |
| Eugenes fulgens | 0.02 | MA | 2 | Nectarívoro |
| Heliodoxa jacula | 0.01 | MA | 2 | Nectarívoro |
| Heliomaster constantii | 0.01 | MA | 2.5 | Nectarívoro |
| Troglodytidae | ||||
| Campylorhynchus rufinucha | 0.07 | R | 2.5 | Insectívoro |
| Cantorchilus modestus | 3.16 | R | 3 | Insectívoro |
| Henicorhina leucophrys | 0.03 | R | 1 | Insectívoro |
| Thryophilus rufalbus | 0.16 | R | 1.5 | Insectívoro |
| Troglodytes aedon | 5.58 | R | 3 | Insectívoro |
| Turdidae | ||||
| Catharus aurantiirostris | 1.90 | R | 2.5 | Frugívoro pequeño |
| Catharus frantzii | 0.03 | MA | 2.5 | Frugívoro pequeño |
| Catharus minimus | 0.01 | MN | 2.5 | Frugívoro pequeño |
| Catharus ustulatus | 0.58 | MN | 2 | Frugívoro pequeño |
| Hylocinchla mustelina | 0.33 | MN | 1.5 | Frugívoro pequeño |
| Turdus grayi | 26.91 | R | 3 | Frugívoro pequeño |
| Turdus plebejus | 0.17 | MA | 2 | Frugívoro pequeño |
| Tyrannidae | ||||
| Contopus cinereus | 0.11 | MN | 3 | Insectívoro |
| Contopus cooperi | 0.20 | MN | 2.5 | Insectívoro |
| Contopus sordidulus | 0.73 | MN | 2 | Insectívoro |
| Contopus virens | 0.64 | MN | 2.5 | Insectívoro |
| Elaenia flavogaster | 1.15 | R | 3 | Frugívoro pequeño |
| Elaenia frantzii | 0.09 | MA | 2.5 | Frugívoro pequeño |
| Empidonax flaviventris | 0.02 | MN | 2 | Insectívoro |
| Empidonax minimus | 0.03 | MN | 2.5 | Insectívoro |
| Empidonax traillii | 0.02 | MN | 2.5 | Insectívoro |
| Empidonax alnorum | 0.06 | MN | 2.5 | Insectívoro |
| Empidonax virecens | 0.15 | MN | 2 | Insectívoro |
| Legatus leucophaius | 0.31 | MS | 3 | Insectívoro |
| Megarhynchus pitangua | 3.36 | R | 2.5 | Frugívoro grande |
| Mionectes oleagineus | 0.07 | R | 2 | Frugívoro pequeño |
| Mionectes olivaceus | 0.16 | MA | 2 | Frugívoro pequeño |
| Myiarchus crinitus | 0.11 | MN | 2 | Insectívoro |
| Myiarchus tuberculifer | 0.02 | R | 2.5 | Insectívoro |
| Myiarchus tyrannulus | 0.02 | R | 2.5 | Insectívoro |
| Myiodynastes luteiventris | 0.29 | MS | 2.5 | Frugívoro pequeño |
| Myiodynastes maculatus | 0.01 | MS | 2.5 | Frugívoro pequeño |
| Myiozetetes similis | 1.47 | R | 3 | Frugívoro pequeño |
| Pitangus sulphuratus | 3.72 | R | 3 | Frugívoro pequeño |
| Todirostrum cinereum | 0.01 | R | 3 | Frugívoro pequeño |
| Tyrannus melancholicus | 6.40 | R | 3 | Frugívoro pequeño |
| Tyrannus tyrannus | 0.18 | MN | 3 | Frugívoro pequeño |
| Tyrannus verticalis | 0.20 | MN | 3 | Frugívoro pequeño |
| Vireonidae | ||||
| Vireo flavifrons | 0.33 | MN | 2 | Frugívoro pequeño |
| Vireo flavoviridis | 1.27 | MS | 3 | Frugívoro pequeño |
| Vireo gilvus | 0.01 | MN | 2.5 | Frugívoro pequeño |
| Vireo leucophrys | 0.13 | R | 3 | Frugívoro pequeño |
| Vireo olivaceus | 1.71 | MN | 3 | Frugívoro pequeño |
| Vireo philadelphicus | 0.19 | MN | 2.5 | Frugívoro pequeño |
La abundancia corresponde a la cantidad de individuos promedios detectados en 89 muestreos. Las especies están clasificadas según su tipo de residencia en el país, dependencia del bosque, y tipo de dieta. *Residencia: R = Residente, MN = Migratoria de Norte América, MA = Especie residente con migración altitudinal, MS = Migratoria de Sur América.
Abundance is the mean number of individuals per sampling day for a total of 89 samples. Each species was classified according to resident status in the country, forest dependence, and food guild. *Resident status: R = Resident, MN = North America Migratory, MA = Resident species with altitudinal migration, MS = South America Migratory.
TABLA 2 TABLE 2 Número de especies de aves promedio (± error estándar) por año según el tipo de residencia en el país, dentro de una matriz agrícola rural en Getsemaní, Heredia, Costa Rica Bird species mean (± standard error) per year according to resident status in the country, inside 30 ha of agricultural rural matrix at Getsemaní, Heredia, Costa Rica
| Año | Todos | Residentes | Migratorio Altitudinal | Migratorio Norteño | Migratorio Sureños |
| 2003 | 36.41 ± 1.84 | 30.61 ± 1.18 | 1.58 ± 0.32 | 5.78 ± 1.46 | 1.37 ± 0.19 |
| 2004 | 32.84 ± 1.90 | 27.73 ± 1.23 | 1.16 ± 0.42 | 6.07 ± 1.05 | 1.04 ± 0.27 |
| 2005 | 38.89 ± 3.50 | 30.65 ± 2.46 | 0 ± 0 | 2.62 ± 1.38 | 1.01 ± 0.3 |
| 2006 | 39.87 ± 1.99 | 33.48 ± 1.33 | 2.17 ± 0.37 | 6.49 ± 1.31 | 1.68 ± 0.25 |
| 2007 | 41.68 ± 2.80 | 35.26 ± 1.97 | 1.58 ± 0.46 | 6.41 ± 1.25 | 1.01 ± 0.39 |
| 2008 | 43.00 ± 4.08 | 37.03 ± 2.97 | 1.22 ± 0.49 | 7.22 ± 1.96 | 1.31 ± 0.4 |
| 2009 | 41.01 ± 3.45 | 35.56 ± 2.41 | 1.77 ± 0.56 | 5.82 ± 2.01 | 1.78 ± 0.32 |
| 2010 | 4.61 ± 1.42 |
La riqueza total de especies fue mayor en enero, febrero, marzo, y setiembre; pero menor en junio y diciembre (F11,68 = 2.85, P = 0.004; Tabla 3). La riqueza de especies residentes fue mayor en enero, febrero, y marzo; pero menor en diciembre (F11,67 = 2.32, P = 0.02; Tabla 3). La riqueza de especies migratorias altitudinales fue mayor en enero y febrero; pero menor en octubre y noviembre cuando no detecté ninguna de estas especies (F9,19 = 2.96, P = 0.02; Tabla 3). Aunque detecté especies migratorias norteñas durante todo el año, la riqueza de estas especies fue mayor de setiembre a abril, y menor el resto del año (F11,58 = 4.68, P < 0.001; Tabla 3). La riqueza de especies migratorias sureñas fue similar entre todos los meses (F7,29 = 0.39, P = 0.90; Tabla 3).
TABLA 3 TABLE 3 Número de especies de aves promedio (± error estándar) por mes (2002 a 2010) según el tipo de residencia en el país, dentro de una matriz agrícola rural, Getsemaní, Heredia, Costa Rica Bird species mean (± standard error) per month (2002 to 2010) according to resident status in the country, inside of agricultural rural matrix at Getsemaní, Heredia, Costa Rica
| Mes | Todos | Residentes | Migratorio Altitudinal | Migratorio Norteño | Migratorio Sureño |
| Enero | 42.60 ± 4.53 | 36.00 ± 3.18 | 1.60 ± 0.98 | 6.60 ± 1.63 | |
| Febrero | 44.75 ± 1.11 | 35.75 ± 1.6 | 1.75 ± 0.48 | 9.00 ± 0.82 | |
| Marzo | 42.36 ± 1.52 | 34.36 ± 1.14 | 1.00 ± 0.36 | 7.27 ± 0.97 | 0.73 ± 0.27 |
| Abril | 36.30 ± 2.08 | 28.30 ± 1.27 | 0.70 ± 0.21 | 7.30 ± 1.09 | 0.70 ± 0.30 |
| Mayo | 33.57 ± 2.19 | 30.86 ± 1.98 | 0.43 ± 0.30 | 1.71 ± 0.52 | 0.86 ± 0.34 |
| Junio | 29.00 ± 2.68 | 27.75 ± 2.50 | 0.75 ± 0.48 | 0.25 ± 0.25 | 1.00 ± 0.41 |
| Julio | 32.83 ± 0.91 | 30.83 ± 0.91 | 0.33 ± 0.21 | 0.33 ± 0.21 | 1.50 ± 0.43 |
| Agosto | 36.10 ± 1.68 | 33.50 ± 1.49 | 0.70 ± 0.26 | 1.50 ± 0.43 | 1.10 ± 0.28 |
| Setiembre | 39.93 ± 2.29 | 32.27 ± 1.86 | 0.33 ± 0.13 | 7.07 ± 1.03 | 0.67 ± 0.21 |
| Octubre | 34.43 ± 1.97 | 27.43 ± 1.15 | 0 ± 0 | 6.57 ± 1.04 | 0.43 ± 0.20 |
| Noviembre | 35.25 ± 4.42 | 27.50 ± 2.40 | 0 ± 0 | 7.75 ± 2.43 | |
| Diciembre | 29.67 ± 3.67 | 22.50 ± 3.01 | 0.33 ± 0.33 | 6.50 ± 1.59 |
Encontré que la riqueza y abundancia de especies fue similar entre los meses al comparar todas las especies (ANOSIM: R = 0.07, P = 0.16; Fig. 2), las especies residentes (R = 0.02, P = 0.34; Fig. 2), y las migratorias altitudinales (R = 0.10, P = 0.14; Fig. 2). Pero existió variación de la riqueza y abundancia de las especies migratorias, tanto norteñas como sureñas, por mes. Los muestreos en setiembre presentaron una mayor similitud de riqueza y abundancia de especies migratorias norteñas entre sí que con los otros meses (R = 0.40, P < 0.001; Fig. 2); mientras que la riqueza y abundancia de especies migratorias sureñas fue más similar dentro del mes de julio y agosto, que entre los otros meses (R = 0.21, P = 0.03; Fig. 2).
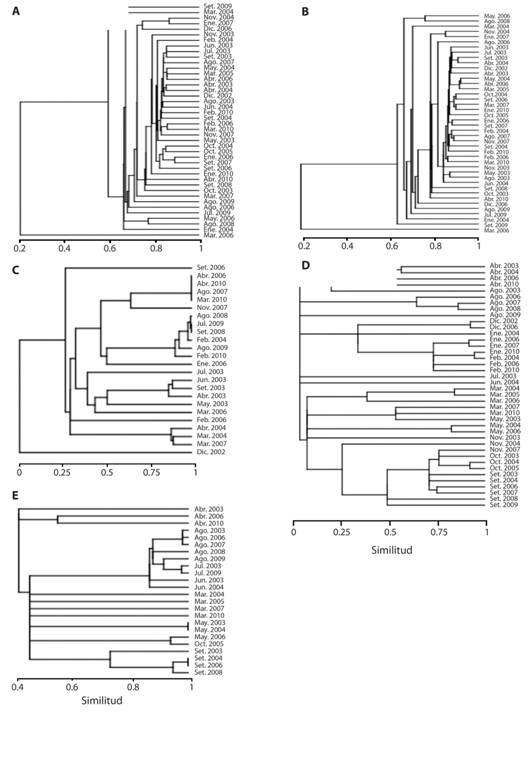
Fig. 2 Fig. 2 Análisis de conglomerados tomando en cuenta la riqueza de especies de aves y su abundancia por mes de 2002 a 2010, utilizando la distancia del índice de Morisita y el método de unión simple. Cada conglomerado representa un tipo de residencia en el país, y en que fueron clasificadas las aves: A) Todas las especies, B) Residentes, C) Migratorias Altitudinales, D) Migratorias Norteñas, y E) Migratorias Sureñas. Valores cercanos a 1 representan una mayor similitud entre la riqueza de especies y su abundancia. Cluster analysis including species richness and abundance by month from 2002 to 2010, using Morisita index of similarity with single linkage union method. Each shows the bird species status in the country: A) All species, B) Residents, C) Altitudinal Migrants, D) Northern Migrants, and E) Southern Migrants. Values close to 1 are high level of similarity in richness and abundance.
La riqueza de especies por grado de dependencia al bosque, varió entre meses únicamente para aquellas especies que viven en áreas con al menos 50 % de cobertura boscosa y clasificadas en las categoría 2 (ALG: Χ2 = 41.52, g.l. = 11, P < 0.001) y aquellas especies que viven tanto en áreas con al menos 50 % de cobertura boscosa y áreas abiertas que fueron clasificadas en la categoría 2.5 (Χ2 = 25.29, g.l. = 11, P = 0.008). Las especies de categoría 2 fueron más abundantes de enero a marzo y menos en junio y julio (Tabla 4); mientras que las especies de categoría 2.5 fueron más abundantes en febrero y menos en junio (Tabla 4). Para las especies más dependientes de bosque (categorías 1 y 1.5) no hubo diferencias entre la riqueza de especies por mes (Χ2 = 5.32, g.l. = 11, P = 0.81; Tabla 4). Lo mismo concluí para las especies de zonas abiertas, donde la riqueza de especies fue igual entre meses (Χ2 = 11.72, g.l. = 11, P = 0.39; Tabla 4). Durante los ocho años de estudio únicamente registré dos aves asociadas a humedales (categoría 4): la Cigueña Americana Mycteria americana y el Chorlito Dorado Chico Pluvialis dominica (Tabla 1), por lo que ningún análisis se realizo con estas especies.
TABLA 4 TABLE 4 Número de especies de aves promedio (± error estándar) por mes (2002 a 2010) según el grado de dependencia de bosque dentro de una matriz agrícola rural en Getsemaní, Heredia, Costa Rica Bird species mean (± standar error) per month (2002 to 2010) according to forest dependence inside of agricultural rural matrix sampled at Getsemaní, Heredia, Costa Rica
| Mes | 1-1.5 | 2 | 2.5 | 3 |
| Enero | 2.67 ± 0.88 | 6.80 ± 1.36 | 12.8 ± 0.92 | 21.40 ± 1.81 |
| Febrero | 1.25 ± 0.25 | 5.50 ± 0.96 | 14.75 ± 1.25 | 23.00 ± 1.47 |
| Marzo | 1.14 ± 0.14 | 5.27 ± 0.54 | 12.91 ± 0.71 | 23.45 ± 0.64 |
| Abril | 2.00 ± 0.58 | 4.50 ± 0.58 | 11.90 ± 0.90 | 19.20 ± 1.31 |
| Mayo | 1.00 ± 0 | 3.00 ± 0.58 | 10.00 ± 0.93 | 20.86 ± 0.99 |
| Junio | 0 ± 0 | 1.50 ± 0.50 | 8.25 ± 1.31 | 19.25 ± 1.03 |
| Julio | 1.00 ± 0 | 1.50 ± 0.22 | 9.00 ± 0.68 | 21.83 ± 0.79 |
| Agosto | 1.20 ± 0.20 | 2.89 ± 0.26 | 10.60 ± 0.72 | 22.30 ± 1.01 |
| Setiembre | 1.00 ± 0 | 3.93 ± 0.44 | 12.93 ± 0.86 | 22.80 ± 1.23 |
| Octubre | 1.00 ± 0 | 3.57 ± 0.81 | 9.14 ± 0.70 | 21.57 ± 0.92 |
| Noviembre | 0 ± 0 | 4.25 ± 1.03 | 10.75 ± 1.89 | 20.25 ± 1.70 |
| Diciembre | 2.00 ± 0 | 3.00 ± 0.71 | 9.00 ± 0.82 | 17.83 ± 2.36 |
Valores de 1 y 1.5 son muy dependientes de bosque y valores de 3 nada dependiente de bosque.
Values of 1 and 1.5 are species highly dependent of forest, and values of 3 are species no-dependent of forest.
Al analizar la riqueza de especies por tipo de dieta solo las frugívoras pequeñas (ALG: Χ2 = 23.86, g.l. = 11, P = 0.01) e insectívoras (Χ2 = 16.49, g.l. = 11, P < 0.001) variaron según el mes. Detecté mayor cantidad de especies frugívoras pequeñas en enero y febrero (Tabla 5); mientras que setiembre presentó la mayor riqueza de aves insectívoras (Tabla 5). Para las demás categorías de aves según su dieta (cazadores, frugívoros grandes, nectarívoros, omnívoros, y semilleros) la riqueza fue similar entre meses (P > 0.14 para todos, Tabla 5).
TABLA 5 TABLE 5 Número de especies de aves promedio (± error estándar) por mes (2002 a 2010) según el tipo de dieta, de una matriz agrícola rural en Getsemaní, Heredia, Costa Rica Bird species mean (± standar error) per month (2002 to 2010) according to food guild, inside of agricultural rural matrix at Getsemaní, Heredia, Costa Rica
| Mes | Cazadores | Frugívoros Grandes | Frugívoros Pequeños | Insectívoros | Nectarívoros | Omnívoro | Semilleros |
| Enero | 3.20 ± 0.73 | 3.20 ± 0.58 | 15.40 ± 2.29 | 9.80 ± 1.36 | 2.00 ± 0 | 2.00 ± 0.45 | 7.00 ± 0.89 |
| Febrero | 3.00 ± 0.41 | 3.00 ± 0 | 15.75 ± 0.85 | 11.75 ± 0.95 | 1.75 ± 0.48 | 2.25 ± 0.48 | 7.25 ± 1.03 |
| Marzo | 3.18 ± 0.40 | 3.18 ± 0.18 | 13.27 ± 0.89 | 10.27 ± 0.89 | 1.64 ± 0.15 | 2.45 ± 0.16 | 8.36 ± 0.51 |
| Abril | 1.78 ± 0.22 | 2.56 ± 0.18 | 11.80 ± 1.09 | 11.20 ± 0.84 | 1.63 ± 0.18 | 1.70 ± 0.21 | 6.40 ± 0.52 |
| Mayo | 2.67 ± 0.33 | 3.14 ± 0.34 | 9.86 ± 0.74 | 8.14 ± 0.77 | 1.86 ± 0.34 | 1.17 ± 0.17 | 7.29 ± 0.81 |
| Junio | 1.75 ± 0.48 | 2.00 ± 0.41 | 9.50 ± 1.32 | 7.00 ± 0.71 | 1.67 ± 0.67 | 1.00 ± 0 | 6.75 ± 0.25 |
| Julio | 1.80 ± 0.20 | 3.17 ± 0.31 | 9.83 ± 0.31 | 8.17 ± 0.79 | 1.20 ± 0.20 | 1.40 ± 0.24 | 8.00 ± 0.68 |
| Agosto | 2.30 ± 0.40 | 3.00 ± 0.33 | 11.00 ± 0.63 | 9.30 ± 0.72 | 1.30 ± 0.21 | 1.44 ± 0.18 | 7.90 ± 0.66 |
| Setiembre | 3.00 ± 0.32 | 2.80 ± 0.24 | 10.53 ± 0.79 | 14.00 ± 1.07 | 1.50 ± 0.15 | 1.53 ± 0.13 | 7.27 ± 0.38 |
| Octubre | 3.00 ± 0.45 | 2.71 ± 0.36 | 10.43 ± 1.02 | 10.57 ± 0.78 | 1.00 ± 0 | 1.57 ± 0.20 | 5.86 ± 0.63 |
| Noviembre | 2.50 ± 0.29 | 2.50 ± 0.29 | 10.50 ± 2.63 | 11.50 ± 1.19 | 1.67 ± 0.67 | 1.25 ± 0.25 | 5.75 ± 0.25 |
| Diciembre | 1.83 ± 0.48 | 2.50 ± 0.34 | 9.50 ± 0.89 | 9.00 ± 1.81 | 1.50 ± 0.34 | 1.40 ± 0.24 | 4.17 ± 0.83 |
Discusión
La riqueza de aves registrada en Getsemaní durante ocho años de estudio fue alta en comparación a otros sitios agrícolas y urbanos relativamente cercanos (ej.: Los Chigüites, Parque La Libertad, Centro Conservación Santa Ana, Universidad de Costa Rica) (Naoki, Duran, & Sánchez, 2003; Alvarado-Quesada & Bolaños-Redondo, 2010; Solano-Ugalde, 2011; Biamonte et al., 2011). Además, la riqueza de especies registradas en el área representa un 17.5 % de toda la riqueza de aves del país (N = 920; Sandoval & Sánchez, 2017), un 19.7 % de las aves terrestres del país, y un 34.9 % de las aves migratorias latitudinales (Sandoval & Sánchez, 2017).
Los datos de riqueza demuestran que las matrices agrícolas que mezclan cafetales con sombra y sol, charrales o tacotales (estadios tempranos de sucesión vegetal), potreros o pasturas, bosques de galería, y zonas urbano-rurales son importantes para la ocurrencia de muchas especies de aves residentes y migratorias. Mis resultados concuerdan con lo reportado en sistemas de café con sombra (Wunderle & Latta, 1996; Johnson, 2000) y plantaciones de cacao (Reitsma, Parrish, & McLarney, 2001; Faria, Laps, Baumgarten, & Cetra, 2006). La riqueza de aves en las matrices mixtas son mayores a la riqueza de las matrices homogéneas, como por ejemplo pastizales, plantaciones de palma aceitera, y piñales, tanto para aves residentes como migratorias (López-Ricuarte, Edwards, Romero-Rodríguez, & Gilroy, 2017).
Los pocos cambios que sufrió la estructura del hábitat del sitio de estudio durante los ocho años de muestreo, pueden explicar porqué la riqueza de especies entre años fue similar (en 7 de 8 años), y no hubo un patrón de aumento o disminución de especies entre años. Posiblemente para detectar cambios de especies dentro de estos ambientes agrícolas-rurales con pocos cambios en la cobertura vegetal es necesario mayores periodos de tiempo, mayores cambios en la matriz circundante, o aumento de la cobertura boscosa como ha ocurrido en otros sitios agrícolas que sí reportan cambios en la riqueza de especies a través del tiempo (Chamberlain, Fuller, Bunce, Duckworth, & Shrubb, 2000; Magurran et al. 2010; Biamonte et al. 2011).
Fenológicamente, cada grupo de aves (según tipo de residencia, dependencia del bosque y dieta) presentó un comportamiento diferente a lo largo del año. Sin embargo, para la mayoría de las categorías de especies donde varió la riqueza por meses, la mayor riqueza ocurrió de enero a marzo, que corresponde con la época seca en el área de estudio (Sandoval, 2011). La mayor riqueza de especies de enero a marzo por tipo de residencia puede ser el resultado de la mezcla de las aves residentes, con las migratorias norteñas, y los residentes que realizan migración altitudinal durante esos meses como lo registré.
Por su parte, de diciembre a marzo es común que en las partes altas de las montañas en Costa Rica y en especial las que dividen la vertiente Caribe de la Pacífico presenten condiciones climáticas adversas para las aves, como fuertes vientos, lluvias constantes y bajas temperaturas (obs. pers.). Esto favorece que varias especies de aves (ej.: Paloma Collareja Patagioenas fasciata, Pitorreal Ptiliogonys caudatus, o Rualdo Chlorophonia callophrys) se desplacen de las zonas altas a las zonas bajas aledañas en busca de mejores condiciones climáticas o alimento (Avalos, 2005), por lo que aumenta la probabilidad de encontrar una mayor riqueza de especies a menor altitud. Por otro lado, aunque de enero a marzo ya no hay aves migratorias desplazándose de norte a sur, que es el momento de mayor riqueza de este grupo de especies en el país (Stiles & Skutch, 1989; Stiles, 1994); la riqueza de aves migratorias en el área de estudio fue constante durante los ocho meses que hibernan en el país (Tabla 2), contribuyendo durante ese periodo a la riqueza total del sitio de estudio.
Un aumento en la riqueza de especies con dependencia intermedia del bosque (categorías 2 y 2.5) de enero a marzo se puede atribuir a que la mayoría de aves migratorias altitudinales y norteñas que se encontraron en el área de estudio son clasificadas en estas categorías (Stiles, 1985; Biamonte et al., 2011), y para estos meses es cuando hay una mayor riqueza de especies en el área. Mientras que las especies con grado de dependencia alta (categorías 1 y 1.5) y baja del bosque (categoría 3) no presentaron cambios en la abundancia de riqueza entre meses. Las especies con alto grado de dependencia del bosque son raras en el área de estudio, debido a que las áreas grandes de bosque son escasas en los alrededores. La falta de bosques maduros primarios o secundarios son el resultado principalmente de una gran expansión cafetalera y agrícola al interior del Valle Central de Costa Rica y posteriormente al desarrollo urbanístico que disminuyó el área de los bosques naturales y las conexiones entre los bosques remanentes (Stiles, 1990; Biamonte et al., 2011). Finalmente, que la riqueza de aves poco dependientes de bosque (categoría 3) fuera constante entre los meses, se explica porque el recurso que utilizan estas especies fue constante todo el año.
Al analizar la riqueza de especies por el tipo de dieta los frugívoros pequeños variaron su riqueza a lo largo del año, aumentando de enero a marzo. Lo anterior se puede deber a que, durante el periodo seco en el área de estudio es común encontrar árboles como higuerones Ficus sp. (Moraceae), arbustos y epífitas (ej.: Loranthaceae) en su pico de fructificación (obs. pers.), favoreciendo la ocurrencia de especies en esta categoría como Rualdo, Veraneros (Euphonia elegantisima), Yigüirros de Montaña (Turdus plebejus), Siete Colores (Tangara larvata), Cardenales (Piranga rubra), o Caciques (Icterus galbula) que pueden aprovechar la abundancia de esos frutos (Stiles & Skutch 1989). Pese a esta variación en frugívoros pequeños y su posible asociación con la abundancia de frutos, los frugívoros grandes no variaron su abundancia a lo largo del año. Esto se podría deber a que en general estas especies son menos comunes tanto dentro (como lo encontré) como alrededor del área de estudio, y por lo tanto pese a que su abundancia puede variar en respuesta a la presencia de frutos, estas variaciones son pequeñas y por lo tanto no fueron detectables en este estudio. Por otro lado que el gremio insectívoro fuera más abundante en setiembre esto porque la mayoría de aves migratorias norteñas son principalmente insectívoras y para este mes es cuando incrementa este grupo de especies en el área de estudio. La riqueza de aves en las otras categorías de dieta fue similar a lo largo del año, lo que puede indicar que el recurso alimenticio es constante durante el año en el sitio.
En conclusión, este estudio reveló diferentes patrones de riqueza de aves en un paisaje agrícola-rural a lo largo del año según el grado de dependencia del bosque, tipo de dieta, y residencia en el país. También demostró que las matrices agrícolas-rurales permiten la ocurrencia de una gran riqueza de especies residentes y migratorias. Lo anterior demuestra que estas matrices poseen una riqueza de especies muy alta y en constante variación a lo largo del año. Por lo tanto si se pretende generar políticas de conservación para mantener estos ambientes (ej.: evitar que se transformen en sitios urbanos) y las aves que los habitan se deben tomar en cuenta los cambios en los patrones de riqueza por grupo de especies ya que no todas varían de la misma forma.
Declaración de ética: el autor declara que está de acuerdo con esta publicación; que no existe conflicto de interés de ningún tipo; y que ha cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

