











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkExisten evidencias que el efecto invernadero es producto del incremento de las emisiones de gases generadas por el hombre (IPCC, 2000), el CO2 ha incrementado de 280 a 375 mg.kg-1 en la atmósfera en los últimos 200 años y continúa el aumento a una tasa anual superior a 1.5 mg.kg-1 (White, Velarde, Alegre, & Tomich, 2005), especialmente por actividades antrópicas en combinación con eventos naturales que inciden sobre la disminución de la vegetación. También al disminuir la vegetación también se merma su capacidad para incorporar el carbono al proceso fotosintético en los diferentes órganos de las plantas. La fotosíntesis es una estrategia para disminuir el CO2 en el ambiente, con especial énfasis en algunas especies perennes de amplia distribución y rápido crecimiento, como: Dendrocalamus latiorus (Munro) var. latiorus (Lin), Guadua angustifolia Kunth, G aculeata (Rupr. ex E. Fourn) y Phyllostachys heterocycla var. pubescens (J. Houz) (Lobovikov, Paudel, Piazza, Ren, & Wu, 2007).
Algunas especies de bambú tienen altas tasas de crecimiento de 7.5 a 100 cm por día (Buckingham et al., 2011) que favorecen la acumulación de carbono orgánico (Lobovikov, Schoene, & Lou, 2012) en cantidades sustanciales para mitigar los efectos del cambio climático (Nath, Lal, & Das, 2015). Se ha demostrado con P. pubescens en China donde presenta diferente capacidad de fijación de carbono con 31 Mg.ha-1 (Yiping, Yanxia, Breckinghan, Henley & Guomo, 2011), 37 Mgha-1 (Huaqiang et al., 2011) a 40 Mgha-1 (Zhang, et al., 2014) al año. En Taiwán con P. heterocycla se cita fijación anual de 41 Mg.ha-1 y con P. makinoi Ohwi de 50 Mg.ha-1 (Yen & Lee, 2011). En India con D. strictus (Roxb.) Nees se consignan 30 Mg.ha-1 (Singh & Singh, 1999), con Pseudostachyum polymorphum Munro 23 Mg.ha-1 y con Schizostachyum dulloa (Gamble) 21 Mg.ha-1 (Singnar et al., 2017).
Bambusa oldhamii Munro fija 51.98 Mg.ha-1 de carbono al año en México (Castañeda, Vargas, Gómez, Valdez, & Vaquera, 2005), en Bolivia 27.53 Mg.ha-1 (Rojas-Quiroga et al., 2013) y en Costa Rica 46.91 Mg.ha-1 (Elizondo-Briceño et al., 2016). En Colombia G. angustifolia Kunth fija 20.9 Mg.ha-1 de carbono (Camargo et al., 2010), mientras que otros autores citan amplia escala de fijación de 24.6 a 51.98 Mg.ha-1 (Arango & Camargo, 2010), o bien, 54 Mg.ha-1 (Riaño, Londoño, López, & Gómez, 2002). Con base en lo anterior, se evaluó cuantitativamente la biomasa aérea de G. angustifolia y B. oldhamii, para determinar la tasa de secuestro de carbono y caracterizar los principales rasgos de manejo de bambú en comunidades de los municipios de Teziutlán y Hueytamalco, Puebla, México.
Materiales y métodos
La investigación se desarrolló con dos especies y dos localidades. La primera con B. oldhamii en el Rancho La soledad del municipio de Teziutlán, Puebla, México (19°46’ - 19°58’ N & 97°19’ - 97°25’ W) a 1 600 m de altitud con clima templado húmedo, precipitación promedio de 2 350 mm anuales, temperatura media de 18 °C y suelos Andosoles. G. angustifolia, se estudió en el Rancho El bambusal, comunidad Loma Alta, en Hueytamalco, Puebla, México (19º51’ - 20º12’ N & 97º12’ - 97º23’ W) a 582 m de altitud. El clima es semicálido húmedo con lluvias en verano, la precipitación promedio anual es de 2 000 mm y la temperatura media de 22 ºC con suelos regosoles (INEGI, 2010). Las especies presentan patrones de crecimiento diferentes en el rizoma. En G. angustifolia Kunth es monopodial y en B. oldhamii Munro es simpodial. Estos atributos fueron estudiados con metodologías distintas en cada especie.
Tamaño de muestra y variables: En ambas especies, la estimación de población y distribución de biomasa aérea se obtuvo a una selección del 10 % de una densidad de 400 plantas en una hectárea (ha), a las cuales se registró diámetro de tallo (cm) a 1.3 m de altura; tomando en cuenta el establecimiento del cultivo el año 2011. Los muestreos comenzaron el año 2016 cuando la planta ya tenía una edad comercial de 5 años, considerando el año 2016 como año uno para los diámetros comerciales de los culmos. La altura de las plantas se determinó mediante el equipo Field Map Data Collector (IFER-Monitoring and Mapping Solutions, Ltd, Republica Checa).
La estimación de la densidad de B. oldhamii Munro en la plantación establecida en 2011 se obtuvo con los datos de 40 individuos con follaje maduro tomados al azar en los culmos como indicador de máximo crecimiento. En G. angustifolia Kunth se determinó en una muestra de 400 plantas y se consideró el número de culmos que emergieron en el año 1 (2016). A través de la relación del diámetro y el año de la planta se calculó la biomasa total de ambas especies para estimar el coeficiente de regresión por el modelo Y = αDβ (Castañeda et al., 2005).
Para ambas especies se aplicó el ajuste propuesto por Baskerville (1972) para eliminar el sesgo asociado a la transformación logarítmica del modelo, que expresado nuevamente en escala aritmética se define como:
Yci = e (α+β ln(D)+CME/2)
Donde Yci es la biomasa calculada del iésimo componente, e es la base del logaritmo natural y CME es el cuadrado medio del error calculado por componente y edad.
Los parámetros usados para obtener biomasa en B. oldhamii Munro fueron constantes, pendientes y valores de R2 para las ecuaciones de biomasa total. En G. angustifolia Kunth se utilizaron parámetros que relacionaron la biomasa seca de tallos, ramas y follaje (Cuadro 1).
Cuadro 1 Constantes, parámetros y valores de R2 para las ecuaciones de biomasa aérea de B. oldhamii y G. angustifolia
| - | Coeficientes usados para B. oldhamii* | - | ||
|---|---|---|---|---|
| Edad (años) | Alfa | Beta | CME | R2 |
| 1 | 6.85 | 1.24 | 0.0142 | 0.77 |
| 2 | 5.75 | 1.84 | 0.0112 | 0.79 |
| 3 | 5.07 | 2.23 | 0.0144 | 0.95 |
| 4 | 6.02 | 1.64 | 0.0153 | 0.87 |
| - | Parámetros y valores para G. angustifolia** | - | ||
| Estructura de la planta | Alfa | Beta | CME | R2 |
| Tallos | 2.6896 | 0.961 | 0.0478 | 0.9524 |
| Ramas | 0.0510 | 1.443 | 0.1214 | 0.8762 |
| Follaje | 9.4 x 10-6 | 4.027 | 0.3665 | 0.7153 |
| Total | 2.2311 | 1.059 | 0.0508 | 0.9480 |
*(Castañeda et al., 2005); **(Riaño et al., 2002).
Resultados
La distribución de los culmos en el terreno fue diferente por especie (e.g. G. angustifolia y B. oldhamii) debido a sus diferencias en morfología y crecimiento (Fig. 1). La densidad de culmos en G. angustifolia fue de 3 300 ha-1, la cual y ésta densidad de población que representa un valor intermedio en plantaciones comerciales establecidas.

Fig. 1 (A) Distribution of the culms of B. oldhamii, in Rancho la Soledad, and G. angustifolia. (B) Rancho el Bambusal, Puebla, Mexico.
La biomasa en brotes con el mismo diámetro se incrementó Se incrementó la biomasa en brotes con el mismo diámetro en B. oldhamii en comparación a G. angustifolia. Además, se presentó relación de crecimiento definido por edades y diámetro en las dos especies. En el primer año, los culmos estuvieron definidos por edades más tempranas en G. angustifolia hasta alcanzar la madurez final, es decir, el diámetro no cambió con el tiempo.
El crecimiento simpoidal de B. oldhamii aumentó su dinámica de crecimiento y en consecuencia la densidad de plantas a través del tiempo. Después de 2 años se incrementó 24 % el número de culmos. Sin embargo, en el año tres, el aumento representó 110 % con relación al año anterior, y en el cuarto año, disminuyó la emisión de tallos a un nivel inferior a la población inicial (Fig. 2). La extracción de culmos abre espacio para el desarrollo de nuevos brotes y se refleja en aumento de sus dimensiones, también se incrementa el intervalo dimétrico de la especie hasta el año 1, con diámetros de 10 cm. En especies de bambú de tipo simpodial, los culmos jóvenes son más productivos que los viejos, por lo que esta condición genera la recomendación de no cosechar los culmos menores de cuatro años de edad.
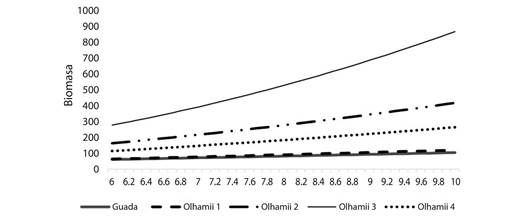
Fig. 2 Relación del diámetro y biomasa de G. angustifolia y B. oldhamii en plantaciones forestales de Puebla, México. Fig. 2. Relation of the diameter and biomass of G. angustifolia and B. oldhamii in forest plantations of Puebla, Mexico.
Disminuyeron los culmos en B. oldhamii después del año cuatro de siembra a este tiempo, la relación diámetro-biomasa está por debajo de la curva de crecimiento de los años anteriores. En cambio, las plantas jóvenes incrementan el desarrollo de ramas laterales. B. oldhamii produce más biomasa en comparación a G. Angustifolia y B. oldhamii incrementa su crecimiento dimétrico culmo por culmo.
El número de culmos por planta aumentó gradualmente hacia las generaciones más recientes, pasó de 3.96 en la cohorte más adulta (cuatro años de edad) a 4.5 en la más joven (un año de edad) (Cuadro 2). El crecimiento del diámetro en los culmos de B. oldhamii fue exponencial durante los tres primeros años después de la siembra (Fig. 3 a), sin embargo, a este tiempo, el crecimiento de la población no alcanza la estabilidad (Fig. 3 b), el culmo continúa creciendo y es característico en plantaciones jóvenes.
Cuadro 2 Promedios de los culmos de B. oldhamii en cuatro cohortes en Teziutlán Puebla, México
| - | - | Edad de culmo (años) | - | ||
|---|---|---|---|---|---|
| Variable | 4 (2013) | 3 (2014) 2 (2015) | 1 (2016) | ||
| Culmos planta-1 | 3.96 | 12.05 | 5.71 | 4.57 | |
| Culmos ha-1 | 120 | 470 | 223 | 184 | |
| Diámetro (cm planta-1) | 4.96 | 6.3 | 7.49 | 7.33 | |
| Intervalo diamétrico | (1.7-6.6) | (4.1-8.9) | (4.3-9.7) | (2.6-10) | |
| Altura (m planta-1) | 9.06 | 14.65 | 18.05 | 17.4 | |
| Intervalo en altura | (7.4-4.6) | (12.3-8.8) | (14.8-21.5) | (13.9-0.45) | |
Averages of the culms of B. oldhamii in four cohorts in Teziutlán Puebla, México
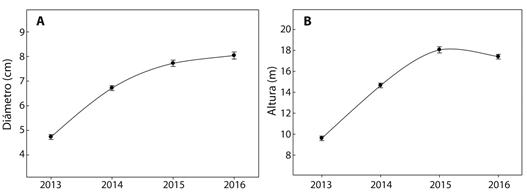
Fig. 3 Diameter and height of the culms of Bambusa oldhamii in the last four years, in plantations located in Teziutlán, Puebla, México.
La biomasa aérea total estimada de B. oldhammi con base a la ecuación de Castañeda et al., (2005) fue de 10 340 348 Mgha-1 distribuida en culmos de los cuatro años. Lo anterior representó acumulación promedio de 25 Mgha-1 por año. La biomasa aérea total estimada con base en la ecuación de Riaño et al., (2002) en la plantación de G. Angustifolia fue de 47 665 11 Mgha-1. Para ambos casos y de acuerdo con Brown (1997), quien cita que la biomasa total acumula 50 % del carbono en tejido vegetal, se estimó, que la cantidad de carbono retenida y calculada en la parte aérea de la plantación de B. oldhammi sería de 12.5 Mgha-1, y para G. Angustifolia de 23.8 Mgha-1 por año.
Discusión
La densidad de culmos en G. angustifolia representa un valor intermedio en otras plantaciones establecidas, con valores de 1 000 hasta 8 000 culmos ha-1 (Riaño et al., 2002). Sin embargo, la densidad cambia con el tiempo al incorporarse más culmos cada año (Riaño et al., 2002), como sucede con B. oldhamii por su desarrollo simpodial (Castañeda et al., 2005). Las diferencias encontradas en los niveles de población, sugieren un proceso de regulación natural por parte de la planta a través del tiempo.
La disminución de culmos en B. oldhamii después del año cuatro de siembra es probable que se deba a la competencia por espacio lateral en las plantaciones y la concomitante disminución de la actividad fotosintética. A este tiempo, la relación diámetro-biomasa está por debajo de la curva de crecimiento de los años anteriores. No obstante, las plantas jóvenes incrementan el desarrollo de ramas laterales y la distribución de la biomasa está influenciada por la disponibilidad de la luz y la edad de las especies (Anten & Hirose, 1998). B. oldhamii produce más biomasa en comparación a G. Angustifolia y B. oldhamii incrementa su crecimiento dimétrico. El efecto anterior sugiere, mayor fijación de carbono e incremento de biomasa aérea y radical (Fig. 2).
El crecimiento diferencial entre altura y diámetro del culmo en las dos especies estuvo definido por la edad. En el primer año en G. angustifolia el diámetro no varió y este comportamiento se atribuye a la ausencia de crecimiento secundario (Judziewicz, 1999). Resultados semejantes a los encontrados en esta investigación cita Cusack (1997) con alturas de 18 m y diámetros de 10 cm en la misma especie. En cambio, en B. oldhamii se incrementó de manera exponencial durante los tres primeros años. Este crecimiento es característico de plantaciones jóvenes (Chrystanty, Mailly, & Kimmins, 1996), pero con altura inferior entre 6 y 9 m y diámetros de 5 a 6 cm en los culmos de B. oldhamii (Shilin, Naixun, & Maoyi, 1994). Es probable que el incremento contrastante en crecimiento en la misma especie esté influenciado en condiciones ambientales en cada región.
La cantidad de biomasa aérea y los valores de Carbono almacenado calculados en la parte aérea para ambas especies está dentro de las cantidades registradas otros autores (Kleinhenz & Midmore, 2001; Arango & Camargo, 2010) con variaciones locales influenciadas por condiciones bióticas y abióticas, edad y densidad de la población. El rápido crecimiento de las dos especies y la aceptación local en la Sierra Nor Oriental de Puebla permite considerarlas como sumidero de carbono, además, aumenta las posibilidades de usos y aplicaciones de la planta.
En el presente trabajo se demuestra que las plantaciones comerciales de bambú en la Sierra Nor Oriental de Puebla, que utilizan especies como G. Angustifolia y B. oldhamii por su facilidad de manejo y tradición. El crecimiento simpodial de B. Oldhamii permite pronosticar su crecimiento con facilidad. Tomando en cuenta que B. Oldhamii es una especie introducida en la zona hace más de 15 años, ha desarrollado un papel importante en el desarrollo económico de la zona por su facilidad de manejo y capacidad de almacenar carbono; estas especies pretenden ser el potencial económico de la zona mediante las distintas formas de aprovechamiento del bambú.
Con los resultados obtenidos de la biomasa aérea total estimada en la plantación de B. oldhamii de 103 403 48 Mgha-1 distribuida en los culmos de los cuatro años y los concomitantes beneficios potenciales al ambiente a medida que aumenta su diámetro, presenta diferencia notable en comparación con G. Angustifolia que logra 50 % menos biomasa aérea total estimada. Lo anterior permite proponer a B. oldhamii en la zona como especie promisoria de bambú para disminuir las concentraciones de carbono.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de cualquier tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. El documento firmado se encuentra en los archivos de la revista.

