











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkSpatial distribution and reproduction are closely interlinked in benthic marine invertebrates, including echinoderms (Mercier & Hamel, 2009). Reproduction in sea cucumbers or holothuroids (Echinodermata: Holothuroidea) was demonstrated to require inter-individual chemical exchanges (Hamel & Mercier, 1996) and minimum densities of individuals to work efficiently (Anderson, Flemming, Watson, & Lotze, 2011). Most known species of sea cucumbers reproduce sexually and are either gonochoric (Mercier & Hamel, 2009; Leite-Castro et al., 2016) or in some cases hermaphroditic (e.g. Herrero-Pérezrul, Reyes-Bonilla, Garcia-Dominguez, & CintraBuenrostro, 1999). Gametogenesis leading to gamete release typically follows seasonal and/ or monthly cycles in low latitudes (Mercier, Ycaza, & Hamel, 2007; Leite-Castro et al., 2016), whereas it generally occurs annually, towards spring and early summer spawning in higher latitudes (e.g. Hamel, Himmelman, & Dufresne, 1993). This is exemplified by the sea cucumber Isostichopus fuscus that reproduces seasonally between July and September in Mexico (Herrero-Pérezrul et al., 1999), but reproduces monthly year-round in equatorial latitudes along the coast of Ecuador (Mercier et al., 2007). Similar patterns were reported in the Indo-Pacific, e.g. in the sea cucumber Holothuria scabra (Conand, 1993a). The gonad index (GI) also exhibits seasonality in other tropical species such as H. atra (Chao, Chen, & Alexander, 1994; Abdel-Razek, Abdel-Rahman, & El-Shimy, 2005; Muthiga, 2006), Actinopyga mauritana (Hopper, Hunter, & Richmond, 1998) and H. fuscogilva (Ramofafia, Battaglene, Bell, & Byrne, 2000) and Stichopus herrmanni (Balogh, Wolfe, & Byrne, 2018).
It is clear from the body of literature (e.g. Engstrom, 1980; Conand, 1982; Costelloe, 1985; McEuen, 1988; Hamel et al., 1993; Herrero-Perezrul et al., 1999; Drumm & Loneragan, 2004; Muthiga, 2006; Mercier et al., 2007; Balogh et al., 2018) that mature oocytes and spermatozoa of broadcast-spawning species must be present at the same time in the water column and in sufficient concentration to insure adequate fertilization rates. A review of publications on the control of reproduction in sea cucumbers and other echinoderms indicates that several endogenous and exogenous factors, acting either successively or in synergy, may be implicated in reproductive coordination (Mercier & Hamel, 2009). Exogenous factors controlling gametogenesis include temperature (Herrero-Perezrul et al., 1999; Drumm & Loneragan, 2004; Toral-Granda & Martínez, 2007), light intensity (Conand, 1982; Muthiga, 2006), photoperiod (Hamel et al., 1993), food availability (Smiley, McEuen, Chaffee, & Krishan, 1991), and change in food type and diffusible chemical signals as well as chlorophyll-a (Leite-Castro et al., 2016). Factors identified as spawning cues include light intensity (Cameron & Fankboner, 1986; Hamel & Mercier, 1995), time of the day (McEuen, 1988), seawater temperature (Conand, 1982), photoperiod (Conand, 1982), water velocity (Engstrom, 1980), salinity (Krishnaswamy & Krishnan, 1967), a combination of seawater temperature and photoperiod (Costelloe, 1985), lunar phase (Mercier et al., 2007), as well as phytoplankton abundance (Hamel et al., 1993; Muthiga, 2006).
Of the 49 species of holothuroids reported for the Central American Caribbean Sea (Alvarado, Solis-Marin, & Ahearn, 2008) at least 21 occur along the coast of Belize (Pawson, 1976; Hendler & Pawson, 2000; Rogers, 2013), of which H. mexicana is the most abundant (Rogers, 2013). Both H. mexicana and its reported hybrid H. floridana × H. mexicana (Fuente-Betancourt, Jesus-Navarrete, Sosa-Cordero, & Herrero-Perezrul, 2001) are large species that can attain lengths of ~50 cm (Fuente-Betancourt et al., 2001; Hasbún & Lawrence, 2002), although lengths of 58 and 76 cm for males and female, respectively, were reported from Florida, USA (Engstrom, 1980). The distribution range of H. mexicana is primarily in the Western Central Atlantic, in the Caribbean Sea (Bruckner, 2006), in Florida (Engstrom, 1980), Venezuela (Tagliafico, Rangel, & Rago, 2011) and the Yucatan Peninsula (Mexico) (Fuente-Betancourt et al., 2001), although it has also been reported in the Azores (Toral-Granda et al., 2013).
Various aspects of population biology in H. mexicana, including density, abundance, distribution and size structure, have been studied in Mexico (Fuente-Betancourt et al., 2001), Panama (Guzman & Guevara, 2002), Costa Rica (Bolaños, Bourg, Gomez, & Alvarado, 2005), Venezuela (Tagliafico et al., 2011) and Belize (Rogers, 2013). Rogers (2013) documented high abundances of H. mexicana in Southern Belizean waters, with up to 12.9 ind ha-1. Based on habitat preferences, H. mexicana was found primarily in seagrass (Thalassia testudinum and Syringodium filiforme) (Hendler, Miller Pawson, & Kier, 1995), coral rubble and patch reef habitats (Rogers, 2013). Fragmentary information is also available on the reproductive biology of H. mexicana. In Florida, gametogenesis and spawning were shown to occur during the spring-summer and late summer, respectively (Engstrom, 1980; Mosher, 1982). However, adults were noted to possess mature gametes from SeptemberNovember, outside peak spawning months (Engstrom, 1980). For the Curacao Reefs, van Veghel (1993) reported that > 70 % H. mexicana individuals spawned after the full moon between August and October. In Panama, Guzman, Guevara, & Hernandez (2003) set size at sexual maturity for H. mexicana between 13-15 cm and its maximum reproductive activity between May-July, when the temperature was 2 °C above the annual average and 3-4 °C higher than the lowest monthly average.
In espite of H. mexicana being the most abundant sea cucumber in the Caribbean as well as one of the most important commercial species in the region, basic biological questions remain incompletely answered and do not allow a full understanding of their vulnerability and resilience to growing anthropogenic pressures (loss of habitat, pollution and increased fishing pressure). Moreover, H. mexicana has been commercialized in Haiti (Ramdeen, Belhabib, Harper, & Zeller, 2012; Masters, 2014; Global Companies, 2015), Martinique (Daniel Taverny, sea cucumber exporter, pers. comm., February 2017), Honduras (SAGARPA, 2015), Nicaragua (Rogers, Hamel, & Mercier, 2017), Jamaica (Jamaica Observer, 2015; Kong, 2016), Mexico (Toral-Granda, 2008) and Belize (Perez & Garcia, 2012). In an effort to provide information of value to decision makers regarding the sea cucumber fishery in Belize and elsewhere in the distribution range of H. mexicana, the objectives of this study were to: (1) determine its annual reproductive cycle and spawning periodicity; (2) establish correlations with environmental variables (e.g. rainfall, temperature); and (3) analyze the population structure inside and outside protected areas and among habitats.
Materials and methods
Collection of sea cucumbers for size structure analysis: A total of 2 333 individuals of H. mexicana (Ludwig, 1875) were collected between February 2014 and September 2015 by snorkeling in the Caribbean Sea off Southern Belize (Fig. 1). The presence of other sea cucumber species was also documented. Each month, 32 to 400 individuals with lengths between 8 and 47 cm (measured mouth-anus in relaxed state underwater) were collected. The size structure (based on length and eviscerated wet weight) in fished and in protected areas was established from seagrass beds, coral rubble, sand and patch reefs around various cayes (small, low island on a coral reef platform) located either inside, adjacent to, or not near any of four marine protected areas (MPAs) (Fig. 1 and Supplementary material 1).
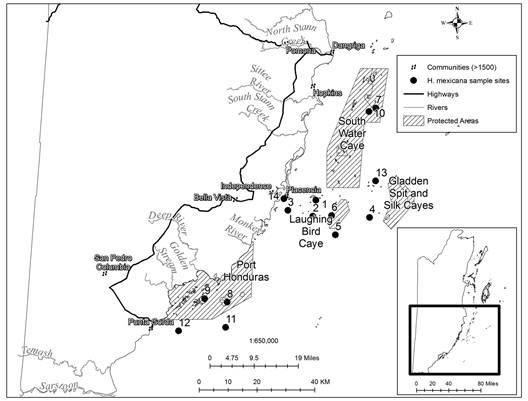
Fig. 1 Collection sites for H. Mexicana in Belize. 1. Lark Range (16˚30.339’ N: 088˚16.337’ W), 2. Sypio (16˚27.307’ N: 088˚16.913’ W), 3. East Placencia (16˚28.377’ N: 088˚21.77’ W), 4. Little Water & Hatchet (16˚27.135’ N: 088˚05.860’ W), 5. South Laughing Bird (16˚23.857’ N: 088˚12.491’ W), 6. West Laughing Bird (16˚27.463’ N: 088˚13.244’ W), 7. Pelican Range (16˚ 47.688’ N: 088 ˚04.7635’ W), 8. Snake Cayes (16˚11.092’ N: 088˚33.438’ W), 9. Tarpon and Abalone (16˚11.690’ N: 088˚37.828’ W), 10. Tobacco (16˚47.021’ N: 088˚06.097’ W), 11. Southeast PHMR (16˚06.347’ N: 88˚33.702’ W), 12. Southwest PHMR (16˚05.616’ N: 088˚42.827’ W), 13. Gladden Cayes (16˚33.993’ N: 088˚04.708’ W), 14. Placencia Lagoon (16˚30.569’ N: 088˚22.536’ W).
Size at sexual maturity, sex ratio and reproductive cycle: During the monthly collection (February 2014 to September 2015) ~30 adult individuals measuring 24.1 ± 6.3 cm SD (n = 587) were used to calculate the gonad index (GI) as percent ratio of gonad weight to eviscerated wet weight, and to conduct histological analysis (Shiell & Uthicke, 2006; Rasolofonirina, Vaitilingon, Eeckhaut, & Jangoux, 2007; Toral-Granda & Martínez, 2007). The gonad and all internal organs were extracted and excess water drained before taking the eviscerated weight. The body wall (including the muscle bands and aquapharyngeal bulb) was measured, and weighted wet (Chao, Chen, & Alexander, 1995; Conand, 1981; Guzman et al., 2003; Leite-Castro et al., 2016). The sex for each individual was established visually from the color of the gonad; creamy or white for the males and orange for the females (Herrero-Perezrul et al., 1999; Guzman et al., 2003; Leite-Castro et al., 2016). A sample of 30 gonads per month were prepared histologically for sex confirmation and level of gonad maturity (described below). Each entire gonad was preserved in 70 % ethanol, subsequently fixed in Bouin’s solution for 24 h, washed alternately in water and 50 % ethanol several times, and finally preserved in 70 % ethanol. The gonads were then dehydrated in a graded series of ethanol baths, embedded in paraffin and sectioned at 1 µm for slide preparation. They were stained with Mayer’s hematoxylin and eosin (Galigher & Kozloff, 1971; Guzman et al., 2003). Five gonad developmental stages were identified for each individual based on Hamel et al. (1993): Stage I post-spawning, Stage II recovery, Stage III growth, Stage IV advanced growth and Stage V mature (see descriptions in the Results section). Sex ratio was established for the entire study but also inside each monthly sample. The size at sexual maturity was determined based on the smallest individuals in samples collected during full gonad maturity period (around March).
Correlation with environmental factors: To understand the influence of external factors on the reproduction of H. mexicana in a location that is characterized by a dry and a rainy season (National Meteorological Service of Belize, 2017), monthly GI and gametogenic stages were compared with the following: mean monthly surface water temperature collected using a YSI Pro Plus Multi-parameter probe at each site; average monthly rainfall data obtained from the Meteorology Department for the Punta Gorda Town weather station (National Meteorological Service of Belize, 2016); mean monthly salinity obtained from the Toledo Institute for Development and Environment (Foley, Alvarez, Moore, Requena, & Warns, 2015); chlorophyll-a obtained from the National Aeronautics and Space Administration (NASA, 2015); day length obtained from Time and Date AS (2015); and tide level data obtained from Mobile Geographics (2016).
After confirming normality and homogeneity of variances using Kolmogorov-Smirnov and Levene’s tests, respectively, analysis of variance (ANOVA) was carried out to compare sea cucumber lengths across sites and habitats. Post-hoc pairwise comparisons were conducted using Tukey’s tests. A Mann-Whitney U test was used to compare the difference between male and female GI. The sex ratio was tested for equality using a Chi-square test. Pearson correlation was used to test the relationship between mean monthly GI and environmental factors for males and females separately. All analyses were conducted using Statistical Package for Social Sciences (SPSS; IBM, 2012).
Results
Population structure: When all sites and dates were combined, the size distribution was unimodal and dominated by individuals of 20 cm (Fig. 2 and Fig. 3). Mean length was 22.4 ± 0.2 cm and ranged from 8 to 47 cm. The mean lengths of individuals varied significantly among the various sites and habitats (ANOVA: P < 0.001), as detailed below.
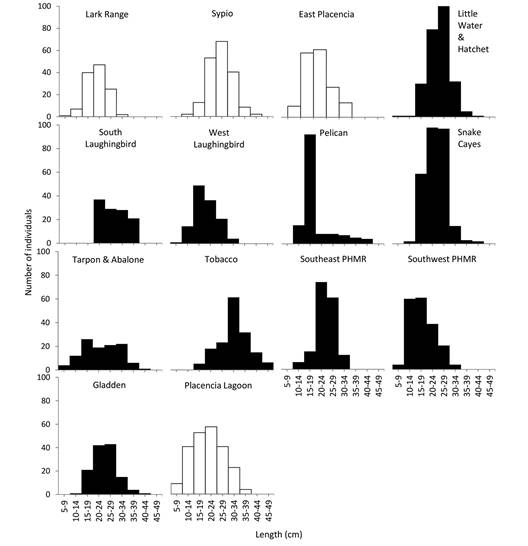
Fig. 2 Length distributions of H. mexicana found inside (black fill) and outside (white fill) boundaries of marine protected areas. See Supplementary material 1 for collection site geographic coordinates, depth and MPA proximity and Fig. 1 for map of collection sites.
In seagrass/coral rubble and sand, the largest individuals (n = 9; 46-47 cm; 430-837 g) found near Tobacco Caye at a depth of 11.3 m were significantly larger (Tukey SD, P < 0.05) than those found in all other habitats. The smallest individuals (n = 49) found in those habitats were between 10 and 13 cm, had a mean eviscerated wet weight between 35 and 65 g and were found in Lark Range, Tobacco, Gladden, Southeast PHRM, Southwest PHMR and Placencia Lagoon (Fig. 1; Supplementary material 1). In seagrass/sand habitats, the largest individuals (n = 45; 40-41 cm; 644-789 g) were found at various depths (Supplementary material 1) at Southwest PHMR, Placencia Lagoon, Sypio Caye and Tarpon and Abalone. The smallest individuals (n = 51; 10.5-12 cm; 35-52 g) were found at Lark Range, Snake Cayes, Placencia Lagoon, Sypio, East Placencia, Southeast and Southwest PHMR (Fig. 1; Supplementary material 1). In seagrass/sand and mud habitats (in the Placencia Lagoon), the largest individuals (n = 12; 35-39 cm; 650-893 g) were found at various depths (Supplementary material 1) and the smallest (n = 2; 8 cm; 16-24 g) contained no gonad. Habitat for the latter was near mangroves (Rhizophora mangle) at depths of 1.2 m (Fig. 1; Supplementary material 1). In patch coral/sand and seagrass the largest individuals (n = 12; 40-46 cm; 90 to 590 g) were found at various depths in Lark Range, Southeast PHMR, Little Water and Hatchet and Tarpon and Abalone (Supplementary material 1; Fig. 1). The smallest individuals (n = 21; 13-14 cm; 72-117 g) were found at Southwest PHMR, Little Water and Hatchet and Snake Cayes.
Eviscerated wet weight distribution was unimodal. The mean for all individuals collected ranged from 14 and 1 088 g with a mean of 317 ± 2.7 g (Fig. 3). The heaviest individuals (n = 89) had a mean weight of 380 g (550-949 g) and a mean length of 38.5 cm (31-46 cm); they were found in areas within or adjacent to MPAs, including Tobacco, Gladden, Little Water, Hatchet and Snake Cayes (Fig. 3). On average, there was a positive relationship between length and weight (Fig. 4) for all individuals, regardless of where they were collected relative to MPAs.
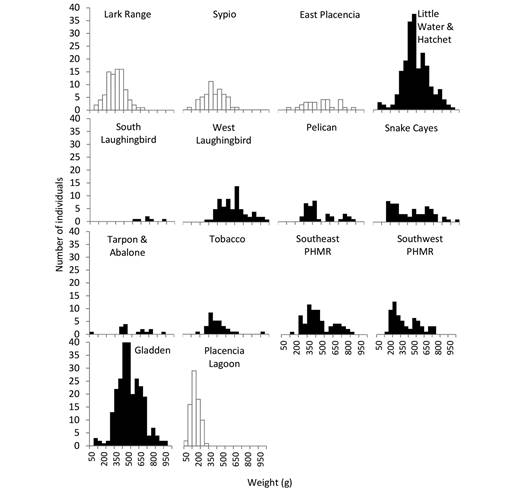
Fig. 3 Eviscerated wet weight distributions of H. mexicana found inside (black fill) and outside (white fill) boundaries of marine protected areas. See Supplementary material 1 for collection site geographic coordinates, depth and MPA proximity and Fig. 1 for map of collection sites.
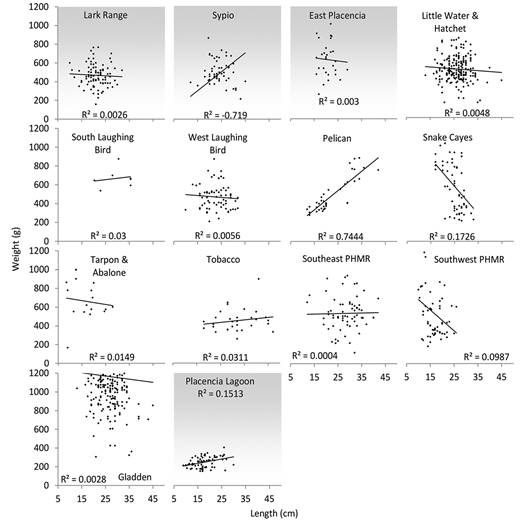
Fig. 4 Length and eviscerated wet weight relationship of H. mexicana found inside (no fill) and outside (gray fill) boundaries of marine protected areas. See Supplementary material 1 for collection site geographic coordinates, depth and MPA proximity and Fig. 1 for map of collection sites.
Other sea cucumber species: Several other species were present in the various sites; these included Isostichopus badionotus (along with H. mexicana in seagrass-associated habitats at all sites), H. thomasi (in coral patches East Placencia), Astichopus multifidus (in coral-associated areas between SWCMR and GSSCMR), Actinopyga agassizi (along with H. mexicana in seagrass-associated areas at Tarpon and Abalone in the PHMR and East Placencia; see Fig. 1 and Supplementary material 1), Euapta lappa (only at night at Pelican Range in the SWCMR; see Fig. 1 and Supplementary material 1) and H. floridana (along with H. mexicana in seagrass-associated habitats at all sites; see Fig. 1 and Supplementary material 1), and three other currently unidentified species (in seagrass habitats East Placencia; see Fig. 1 and Supplementary material 1).
Sex ratio and size at sexual maturity: Overall, there were 27 adult-sized individuals that had no gonad, 2 hermaphrodites (Fig. 5M), 290 males and 268 females, yielding a male:female ratio of 1.1:1.0. The slight apparent dominance of males was not significant (Chi square: P = 0.310). Moreover, monthly collections provided more males than females in some months and vice versa in other months (Supplementary material 2). The slight male dominance in two sites (Sypio and Tarpon) was not significant (Chi square: P = 0.34 and 0.38, respectively). Thirteen of the 27 individuals without gonad were found in Placencia Lagoon and ranged from 8 to 23 cm in length; the other 14 individuals were found in areas near Gladden (n = 1), Lark Range (n = 6), East Placencia (n = 1) and Snake Cayes (n = 6), and ranged from 14-29 cm.
The minimum size at sexual maturity was determined to be around 10 cm in length for both sexes; a size at which both mature and immature individuals occurred. However, it was only at 16 cm that 100 % of individuals were found sexually mature. Macroscopic and microscopic examinations confirmed that cream or white gonads were testes and that orange or purple gonads were ovaries. Macroscopically, the gonad tubules composing the testes were ramified, thinner and on average smaller than the ovaries, which were unramified and heavier. Specifically, testes of all stages combined had a mean weight of 16 ± 2 g ranging from 0.2-103 g, while ovaries of all stages had a mean weight of 26 ± 2 g, ranging from 0.5-305 g, with a significant difference between the two (Mann-Whitney, P < 0.001).
Reproductive cycle: Histological analysis showed that the variations in gonad weight, color and tubule diameter reflected different gametogenic stages (level of maturity). Detailed descriptions of the stages are provided in Supplementary material 3. The oogenic and spermatogenic stages illustrated in Fig. 5, and Fig. 6 show the monthly proportion of each stage in the sampled individuals. In 2014, the proportion of post-spawning stage (I) in males was the highest in August (74 %) and it remained relatively high through December (49-65 %); it was also high in April-May (60-65 %). In 2015, the highest proportions of post-spawning males were again April-May (58 %), and August and September (69 % and 79 %, respectively; Fig. 6A). Males in recovery stage (II) were more frequent from September to November 2014 (35-49 %), and reached 26 % in September 2015 (Fig. 6A). Male gonads in growth stage (III) occurred more frequently in February of 2014 (40 %) and in January of 2015 (77 %; Fig. 6A); this stage was found in all months except in March, April and May of 2014, and March and May of 2015. The advanced-growth stage (IV) in males was more frequent in February (45 %) of both years, whereas the mature stage (V) dominated in March (88 %) and July (83 %) of 2014 and in the same months of 2015 (83 % and 92 %, respectively; Fig. 6A). In 2014, the proportion of post-spawning (I) females was the highest in April, May and August (30 %, 37 % and 32 % respectively) and in April, August and September (60 %, 69 % and 47 % respectively) (Fig. 6B). Female gonads in the recovery stage (II) generally dominated through the fall of 2014, representing 71 % and 77 % in September and October, respectively, whereas in 2015 it was more frequent in May (72 %) and only 38 % in September (Fig. 6B). The highest proportion of female gonads in growth stage (III) was 45 % in February and 36 % in June of 2014 and it occurred more frequently in January-February of 2015 (6068 %; Fig. 6B). The female advanced-growth stage (IV) dominated in June of both years (43 and 56 %, respectively). Based on the dominance of mature and advanced-growth stages, immediately followed by the appearance of post-spawning stages, seen in both sexes, the spawning months were identified as March and July of both years (Fig. 6).
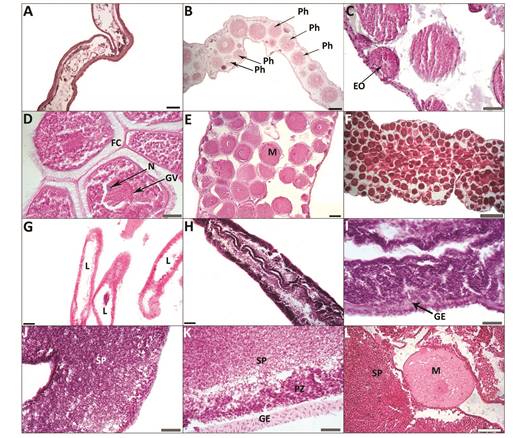
Fig. 5 Holothuria mexicana. Light micrographs of female tubules illustrating the oogenic cycle. (A) Post-spawning; (B) Recovery stage showing nutritive phagocytes (Ph) inside residual oocytes; (C) Growth stage showing early oocytes (EO) in a rather empty lumen; (D) Advanced-growth stage showing abundance of mature oocytes with germinal vesicle (GV), nucleoli (N), follicular cells (FC) and Germinal Vessicle (GV) around nucleous; (E) and (F) Mature stage showing lumen filled with mature oocytes (M). Light micrographs of male gonad tubules illustrating the spermatogenic cycle. (G) Post-spawning testis showing empty lumen areas in the gonad tubules (L); (H) and (I) Growth stage showing the highly convoluted germinal epithelium (GE); (J) and (K) Mature stage showing the thin gonad tubule wall, reduced or absent proliferation zone (PZ), and great numbers of spermatozoa (SP) in the lumen; (L) Hermaphrodite showing a mature oocyte (M) surrounded by a large number of spermatozoa (SP). Black scale bar represents 100 µm (A, B, E, G, H); grey scale bar represents 40 µm (D, F, I, J, K) and white bar represents 500 µm (C, L).
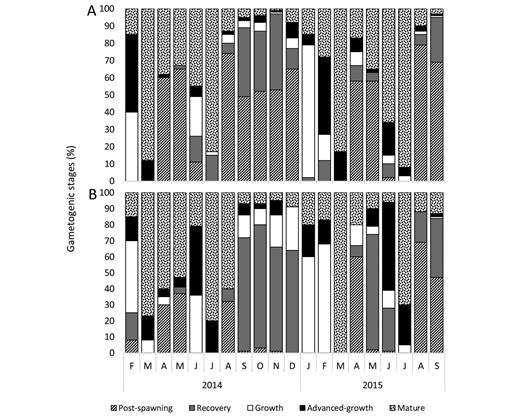
Fig. 6 Monthly percentage of each gametogenic stage for (A) males and (B) females from February 2014 to September 2015.
Gonad indices (GI) in both sexes displayed similar seasonal patterns (Fig. 7G). A gradual increase in mean GI from January-February for both 2014 and 2015 was observed. GI peaks were detected in March of 2014 and 2015 and July 2015 for females. For males, GI peaks were detected in March 2014 and March and June 2015. A gradual decline in GI followed from April to September 2014 for both males and females. The lowest GI values of the annual cycle was from October to January 2014, in both males and females. Similarly, in 2015, there was a gradual decline in GI values for both males and females from April to May but they increased again in June and July and underwent a gradual decrease again in August and September.
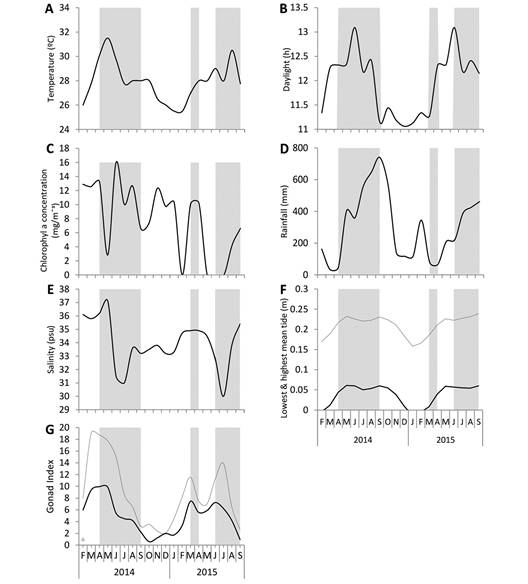
Fig. 7 Environmental parameters measured during the study of H. mexicana from February 2014 to September 2015. (A) sea surface temperature (ºC); (B) day length (h); (C) chlorophyll-a (mg m-3); (D) rainfall (mm month-1); (E) salinity (psu); (F) lowest (continuous line) and highest average tide (dotted line); (G) mean monthly male (continuous line) and female (dotted line) GI. The gray highlights identify months of spawning periods in H. mexicana.
Environmental Factors: During the annual cycle, the GI was positively correlated with sea surface temperature (r = 0.547, P < 0.05; Fig. 7A). The GI started to increase from February to March 2014 with warming temperatures and started to decrease as temperatures dropped from March-April to September. From September 2014 to January 2015, the GI remained at its lowest, concurrent with the coldest temperatures of the annual cycle, and started to increase again in JanuaryFebruary 2015 until March 2015. Correlations were also detected between GI and day length (r = 0.552, P < 0.05; Fig. 7B) with GI values increasing with day lengths and decreasing as days grew shorter. Spawning periods starting between March and April in 2014 and 2015 and again in July-August 2015 showed correlations with fluctuations and peaks in chlorophyll-a. There was a correlation between peak GI and an increase in chlorophyll-a levels (r = 0.551, P < 0.05; Fig. 7C) and the decrease in salinity (r = 0.671, P < 0.05; Fig. 7E). Similarly, there was a parallel between peak GI and the highest high tides (Fig. 7F).
Discussion
Small individuals of Holothuria mexicana found in the present study (around 8 cm long) were much smaller than the smallest individuals found in Panama (16 cm) (Guzman et al., 2003) and in Mexico (17 cm) (Fuente-Betancourt et al., 2001). Although there is currently no regulation to limit the harvest size (Rogers, Hamel, Baker, & Mercier, 2018), fishers typically caught individuals of ~12 - 24 cm total length (Perez & Garcia, 2012), which is above the length at first maturity found here. It is not clear how many are actually harvested annually since the Total Allowable Catch (TAC) is based on wet weight, and the exported volume (as partially or fully dehydrated sea cucumber cucumbers) is used to estimate whether the TAC has been met. Because the dry weight of a sea cucumber is about 5-25 % of its wet weight, this method allows the capture to exceed the quota, which may explain the drastic decline in abundance of both targeted sea cucumber species along the coast of Belize, forcing the closure of the fisheries in 2017 (Rogers et al., 2018). Their small size and abundance in the Placencia Lagoon in Belize (not seen elsewhere throughout the study sites) may suggest that a nursery habitat could be located near or within this site. The area was characterized by shallow seagrass, coral rubble, mud and sand habitats, which were all in the vicinity of Red mangrove forests (Rhizophora mangle). This finding is consistent with studies identifying seagrass close to mangroves as nursery areas for another tropical Holothuroidea sea cucumber, H. scabra, in the Solomon Islands (Mercier, Battaglene & Hamel 2000). The Placencia Lagoon, which is far from any MPA is challenging the assumption that the presence of MPAs can promote or enhance recruitment at least in the areas examined in Southern Belize. This is similar to the findings of Roberts & Polunin (1991) that suggested large areas of unprotected reef could produce as many eggs as small MPAs and that recruitment depends on larval dispersal pathways more than on the protected status of a site. The absence of early recruits in protected areas and their presence in heavily fished areas suggest that MPAs in Belize may not be adequately located or large enough to protect the sea cucumber resources. In fact, these MPAs were developed for other reasons. For instance, South Water Caye Marine Reserve was established in 1996 for its “outstanding universal value” as part of the Belize Barrier Reef System World Heritage Site (UNESCO World Heritage Center, 2017) and to protect valuable habitats for commercial and non-commercial species, including queen conch (Strombus gigas) and lobster (Panulirus argus), on which a number of coastal communities in Belize are dependent (Belize Fisheries Department, 2013). Nevertheless, the largest sea cucumbers were found inside MPAs when compared to other surveyed areas. This difference in size likely reflects the fact that large sea cucumbers (more visible and commercially valuable) are targeted by fishers, thereby decreasing their abundance outside protected boundaries. Therefore, while MPAs seem to protect adults (when no illegal fishing is taking place in these protected areas), these locations are apparently not promoting local recruitment.
Some tropical sea cucumbers have been demonstrated to have specific habitat preferences (Bruckner, Johnson, & Field, 2003). In the present study, the largest individuals of H. mexicana were found associated with coral reef (in areas of sand and seagrass, or sand and coral rubble). The proximity of reef areas could play a role in providing food and habitat for sea cucumbers (Hasbún & Lawrence, 2002; Conand, 2006; Toral-Granda, 2008), particularly when combined with seagrasses as is the case in the present study. Seagrass habitats were demonstrated to serve as settlement areas, and offer shelter and food to the newlysettled larvae and early juveniles of H. scabra (Mercier, Battaglene, & Hamel, 2000). The presence of larger individuals in deeper waters, such as reef areas in this study, may be due to temperature gradients (Choe, 1963), as well as shelter (Sloan & von Bodungen, 1980). Greater food availability in deep waters where organic content is the highest (Yingst, 1982) was also suggested for Stichopus japonicus (Choe, 1963) and Parastichopus parvimensis (Yingst, 1974). Although organic matter seems to be less abundant in reefs than in mangrove and seagrass meadows (Hemminga et al., 1994), corals are commonly found in close association with these two habitats and, depending on local hydrodynamics, seagrass-derived organic carbon may be exported to nearby environments (Unsworth et. al., 2008) including reef areas. Yet other studies suggest that habitat preference (Sloan & von Bodungen, 1980) and distribution of most tropical holothuroids on reef flats is related to shelter from wave action (Bakus, 1973). It is also possible that the abundance of larger individuals in deeper water may simply be the result of fishing activities that deplete them from shallower areas, which are easier to access.
In comparison to previous studies, maximum sizes of H. mexicana found in Belize (47 cm) were larger than in Panama (Guzman et al., 2003) and in Venezuela (Rodriguez-Milliet & Pauls, 1998); but smaller than in Mexico (Fuente-Betancourt et al., 2001) and Florida (USA) (Engstrom, 1980). These differences in sizes could be related to the increasing fishing pressure promoting the disappearance of certain size classes. In Belize, it may suggest that the largest individuals are rare and have been harvested over the years, thereby skewing the population structure compared to earlier published work conducted before extensive fisheries started in the focal locations (e.g. in Mexico where the intensive fishing activity only began about 10 years ago). The largest individuals found here were also smaller than those reported by several fishers and exporters a few years ago, e.g. individuals of 91 cm in Half Moon Caye within the Lighthouse Atoll in Belize (Lincoln Cuevas, sea cucumber fisher, pers. comm., June 2012).
The sizes of H. mexicana found in all sites studied in Southern Belize also varied markedly, although the most common range was 20-24 cm. Since size at sexual maturity was found to be around 10 cm, fished populations are likely mainly composed of mature adult individuals. The present data highlighted a very low abundance of small individuals and juveniles in most sites, suggesting that the density of sexually mature sea cucumbers may no longer be high enough to ensure efficient fertilization during spawning periods. This type of Allee effect was described as a major impediment to the recovery of heavily fished sea cucumber populations, e.g. H. scabra on Warrior Reef, Australia (Skewes, Dennis, & Burridge, 2000), Isostichopus fuscus in the Galapagos Marine Reserve, Ecuador (Toral-Granda & Martinez, 2007), and H. nobilis on the Great Barrier Reef, Australia (Uthicke & Benzie, 2000).
The minimum size at sexual maturity of H. mexicana found in Belize (10 cm in length) was smaller than that determined in Panama (13-15 cm) by Guzman et al., (2003). This size was also smaller than the 16 cm reported for H. atra in the Red Sea (Abdel-Razek et al., 2005), 20 cm for H. sanctori in Canary Islands (Spain) (Navarro, Garcia-Sanz, & Tuya, 2012), 20 cm for H. grisea in Brazil (Leite-Castro et al., 2016), and 27 cm for Stichopus variegatus in New Caledonia (Conand, 1993b). In spite of its maximum size of >50 cm, comparable and sometimes larger than other previouslymentioned commercial tropical sea cucumbers, the small size at which sexual maturity was recorded in H. mexicana in Belize may reflect an adaptive strategy of the species, which is under heavy pressure from the fisheries. Maturation at younger and smaller sizes was proposed for other marine species as a result of high selective fishing pressure (as per Vainikka & Hyvärinen, 2012; Pukk, Kuparinen, Järv, Gross, & Vasemägi, 2013).
The equal proportion of males and females in H. mexicana in Belize was similar to findings in the majority of tropical Holothuroidea species (e.g. Guzman et al., 2003; Foglietta, Camejo, Gallardo, & Herrera, 2004; AbdelRazek et al., 2005; Muthiga, 2006; LeiteCastro et al., 2016). However, some uneven male:female sex ratios have been reported, such as 1.7:1 in H. fuscogilva in the Maldives (Reichenbach, 1999), 4:1 in H. nobilis in Micronesia (Amesbury et al., 1996) and 31:1 in S. chloronotus in La Reunion (Conand, Uthicke, & Hoareau, 2002), all corresponding to heavily fished populations. As hypothesized previously by Birkeland & Dayton (2005) and Fenberg & Roy (2008), harvesting of large individuals can lead to depletion of one sex over the other (e.g. if a size difference exists between the sexes), thereby limiting the species reproductive potential. The 1:1 sex ratio found in the present study suggests that the maximum size of the heavily fished H. mexicana in Belize is similar in males and females and that both sexes were harvested equally. Moreover, the presence of hermaphroditic H. mexicana observed in Belize, also described in the sea cucumber I. fuscus in Mexico, may be linked to fishing pressure (Herrero-Pérezul, Reyes-Bonilla, & Garcia-Dominguez, 1998). It has been hypothesized that, in places where population density of a species is so low, and conspecifics are rare, selection may enhance hermaphroditism and even self-fertilization. Inversely, if population density increases, the presence of hermaphrodites will decrease (Ghiselin, 1969).
Gametogenic stages and GI suggested synchronous gonad development in males and females in both years studied. Although a distinct indication of early gametogenesis (in February 2014 and January-February 2015) and clear spawning seasons were identified in this study (March-April 2014 and 2015 and in July-August 2015), some sea cucumbers with mature gonad stages were found throughout the year (between March and June 2014 and March and May 2015). This is similar to other studies of holothuroid reproduction, including H. mexicana from other locations (Engstrom, 1980; Guzman et al., 2003), which suggests that not all individuals that reach gonad maturity are involved in spawning at the same time. Moreover, Leite-Castro et al. (2016) suggested that, although gametogenesis in H. grisea may be monthly at the population level, it may be asynchronous among individuals in that population since only a portion of the individuals will spawn each month, as was also demonstrated in I. fuscus in Ecuador (Mercier et al., 2007) and H. atra in Northwestern Sri Lanka (Dissanayake & Stefanssong, 2010).
Based on fluctuations in GI values and gametogenic stages, spawning was estimated to occur between March-April of both years. However, the spawning event in March-April 2015 was of smaller amplitude than the 2014 event and, despite clearly visible from the drop in GI and appearance of post-spawning stages, it was followed by a second weak spawning event in August-September 2015. Both spawning periods recorded in Belize in 2015 differ slightly from previous records for H. mexicana that placed the spawning period at the end of August in Florida, USA (Engstrom, 1980), in September and October in Curacao (van Veghel, 1993) and in May and July in Panama (Guzman et al., 2003), suggesting adaptation to local environmental conditions (see below). However, it is consistent with other high-latitude tropical sea cucumber species (Mercier & Hamel, 2009) showing a seasonal reproductive period. The spawning period in both years was followed by months of gametogenic inactivity during which GI values were minimum and gonads were mostly in the recovery stage, as described in several other tropical and temperate sea cucumbers (Mercier & Hamel, 2009).
The reproductive cycle of H. mexicana exhibited a seasonal pattern that was seemingly driven by environmental factors. Gametogenesis was initiated around February in both years (more defined in 2015), a period which corresponds to low rainfall, and increasing salinity, temperature and day length. Increases or decreases in temperature, day length and salinity were previously described as major factors influencing the initiation of gametogenesis in sea cucumbers (Mercier & Hamel, 2009). In benthic marine invertebrates, spawning seasons often correspond to periods that favor recruitment success (Reitzel, Miner, & McEdward, 2004). Spawning in H. mexicana clearly occurred during the most productive period of the year with maximum or highly fluctuating values of chlorophyll-a, increasing rainfall, increasing daylength and rapidly decreasing or increasing temperature. Rainfall in Southern Belize where H. mexicana is most abundant (Rogers, 2013) is the highest for the country with an annual average of about 4 000 mm (National Meteorological Service of Belize, 2016). Increasing rain and runoff of terrigenous material coming from numerous rivers found along the coast could explain the increase of primary production (chlorophyll-a) and the beginning of the spawning period. This timing may well coincide with the presence of larvae of H. mexicana that feed on planktonic food, as also suggested by Leite-Castro et al. (2016) for H. grisea and by Chao et al. (1995) for Actinopyga echinites, H. leucospilota and H. difficilis in southern Taiwan. Such a correlation between primary production and sea cucumber spawning was also observed in the temperate species Psolus fabricii (Hamel et al., 1993) and Cucumaria frondosa (Hamel & Mercier, 1995) and the tropical species H. grisea (Leite-Castro et al., 2016). Guzman et al. (2003) also suggested a relationship between GI values and temperature in H. mexicana in Panama. Here, both annual spawning periods coincided with the highest high tides and highest low tides, similar to the findings of Leite-Castro et al. (2016), suggesting that spawning dates could be determined by the tidal cycle and the related water level. High tides acting as spawning cues were also reported for A. echinites (Desurmont, 1996) and Bohadschia similis (Desurmont, 2004), and believed to facilitate the dispersion of the propagules with currents. Finally, the period characterized by the lowest GI values and a dominance of gonads in recovery stages in the present study was between August and January, corresponding to the period of shortest daylengths, coldest temperatures, stable salinity and minimal rainfall.
The results presented here call for additional studies that seek to understand the adaptive strategies in H. mexicana and the role that adults play in coral reef communities. Investigations of nursery habitats are also needed to fully understand the life cycle and early life-history strategies of H. mexicana. While efforts are being deployed to learn more about the species, it is critical to protect its habitat and restrict fishing around the spawning months because the resource has already been heavily exploited.

