











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkEntender la causa específica de muerte y supervivencia en las poblaciones de borrego cimarrón (Ovis canadensis; especie sujeta a protección especial en la normatividad mexicana (NOM-059-SEMARNAT-2010; Secretaría de Medio Ambiente y Recursos Naturales [SEMARNAT], 2010), es de utilidad al desarrollar estrategias de manejo para la protección y translocaciones dentro de su hábitat histórico (Schaefer, Torres, & Bleich, 2000). Reciente evidencia ha sugerido que la depredación tiene un impacto en la dinámica poblacional en plantas y animales (Courchamp, Clutton-Brock, & Grenfell, 1999). Sin embargo, los escasos estudios de mortalidad por depredación limitan nuestro entendimiento del papel de los depredadores y el impacto que tienen sobre las poblaciones presa.
El borrego cimarrón prefiere hábitats que limiten la capacidad del puma para atacarlos, paisajes relativamente libres de vegetación y terreno accidentado que les permiten ver y evitar a los pumas (Shaefer et al., 2000; Logan & Sweanor, 2001). Sin embargo, el decline de las poblaciones de borrego cimarrón debido a la depredación por puma ocurre donde el ámbito hogareño del borrego cimarrón se traslapa con el del venado bura (Odocoileus hemionus) en densidades adecuadas para proveer presas al puma (Shaefer et al., 2000; Logan & Sweanor, 2001; Rominger, 2018). Esto debido a que el venado bura es la principal presa del puma en ecosistemas desérticos (Ackerman et al., 1986; Leopold & Krausman, 1986; Cashman, Peirce, & Krausman, 1992; Logan & Sweanor, 2001; Villepique, Pierce, Bleich, & Bowyer, 2011) y se sospecha que la abundancia de venado bura en rangos de montaña sirve como atrayente para los pumas (Wakeling et al. 2009). Por lo que Rominger (2018) menciona como una de las primeras causas del incremento de la depredación por puma en borrego cimarrón en las últimas cuatro décadas, el incremento de la presencia del felino en hábitats donde históricamente era ausente o raro, en respuesta a la extensiva conversión de ecosistemas abiertos a matorral por la alteración de los ciclos del fuego seguido de la expansión territorial del venado bura. Además, argumenta que el puma que caza en hábitat de borrego cimarrón en baja elevación o pie de montaña difícilmente selecciona venado.
Hallazgos similares se hicieron en un estudio realizado por Schaefer et al. (2000) que evaluaron la causa específica de muerte y supervivencia en una población simpátrica de borrego cimarrón y buras en San Bernardino, California. En donde 26 borrego cimarrón (10 machos y 16 hembras) y 34 venado bura (nueve machos y 25 hembras) fueron dotados de radio collares. Reportaron un 75 % (9/12) de muertes de borrego por puma con una tasa de supervivencia de 0.80, y para los venados buras encontró un 55 % (5/9) de mortalidad debido a depredación por puma con una tasa de supervivencia de 0.81. Logan & Sweanor (2001) estudiaron una población de venado bura y población remanente de borrego cimarrón en San Andrés, Nuevo México, E.U.A. Monitorearon 175 venados con radio collares (91 machos y 84 hembras) durante los años 1987 a 1994 y a 43 borregos con radio collares de telemetría (16 machos y 27 hembras) entre los años de 1985 a 1995, registrando para venado bura 61 % (62/101) muertes por puma (35 machos y 27 hembras) y para borrego cimarrón 38 % (10/26) bajas a causa de puma (4 machos y 6 hembra). En este estudio concluyen que la depredación por puma fue la mayor causa de muerte para ambas especies.
En 1949, se dio por extirpado al borrego cimarrón de la Sierra Maderas del Carmen, Coahuila, debido a factores como la minería, competencia con borrego doméstico (Ovis aries) y cabras (Capra spp.), fauna exótica como el Berberisco (Ammotragus lervia), enfermedades, cacería no controlada, cercos borregueros, sequía y depredación (Baker 1956; Sánchez 2005). En 2000, CEMEX (Cementos Mexicanos) y la organización no gubernamental Unidos para la Conservación iniciaron un programa de reintroducción del borrego cimarrón en su hábitat histórico en Sierra Maderas del Carmen al norte de Coahuila. El programa empezó con la construcción de un encierro de 5 000 ha libre de depredadores, en Pilares, Ocampo, Coahuila. Durante 2001 y 2002 se capturaron a 48 borregos en Sonora, en la reserva del Yaquí, Punta Cirios, San Ramón e Isla Tiburón para liberarlos en el encierro de Pilares. En 2004, se capturó 32 borregos más de la reserva del Yaquí, para liberarlos a estado silvestre en la Sierra Maderas del Carmen (McKinney & Villalobos, 2005).
Bajo el contexto de esta reintroducción, el objetivo de este estudio es evaluar la tasa de depredación por puma sobre el borrego cimarrón y el venado bura, para analizar la selección entre ambas presas del puma en un mismo hábitat, así como el impacto en las poblaciones. Como hipótesis se plantea que el puma es un depredador oportunista que no discrimina entre especies de ungulados simpátricos generando mayor impacto a la especie de menor densidad. El conocer cómo la depredación por pumas afecta las poblaciones del borrego cimarrón y venado bura en la Sierra Maderas del Carmen es importante para entender la relación depredador-presa y diseñar estrategias efectivas de reintroducción y conservación de estos ungulados.
Materiales y métodos
Área de estudio: La Sierra Maderas del Carmen se ubica al noroeste del estado de Coahuila, México, entre (28°42’18.28” - 29°21’29.42” N & 102°22’04.58” - 102°55’04.03” W). La parte norte y noroeste del área está limitada por el río Bravo, donde colinda con el Parque Nacional Big Bend; al oeste y al sur limita con por la carretera Melchor Múzquiz - Boquillas del Carmen y al este con el camino de terracería El Melón- La Linda. Políticamente forma parte de los municipios de Ocampo, Acuña y Múzquiz. El rango de elevación va desde los 500 m en los márgenes del río Bravo hasta los 2 720 msnm en los picos más altos de la sierra. La precipitación se presenta principalmente en verano que varía de 100-200 mm en las colinas bajas hasta 200-300 mm en las partes más altas. La temperatura promedio anual es de 22 °C, con temperaturas en invierno por debajo de los 0 °C. Las comunidades vegetales presentes son matorral desértico chihuahuense (micrófilo, rosetófilo, halófilo y gypsófilo), matorral submontano, zacatal y bosque de montaña (pino, encino y oyamel) que se presentan de acuerdo al gradiente altitudinal (Secretaría del Medio Ambiente, Recursos Naturales [SEMARNAT], 2013).
Los borregos y venados fueron capturados con la técnica de rifle de red desde un helicóptero (Wehausen, 1996; Kamler, Lee, de Vos, Balard, & Whitlaw, 2002; Rominger, Whitlaw, Weybright, Dunn, & Ballard, 2004; McKinney, Smith, & de Vos, 2006a), del criadero de vida silvestre de Los Pilares, Ocampo, Coahuila en noviembre del 2014 (permiso DGVS/09/04067). A todo los individuos se les cubrieron los ojos durante el manejo para reducir el estrés de captura, una vez contenidos, médicos veterinarios tomaron muestras de sangre, se les inoculó antibióticos, antiparásitos, des-inflamatorios, vitaminas, un reforzamiento del sistema inmunológico y aretes de identificación. Finalmente, individuos mayores de tres años de edad se equiparon con radio collares de telemetría modelo MOD-600 con frecuencia VHF de seis dígitos y sensor de mortalidad de cuatro horas de activación (Telonics Inc., Mesa AZ., E.U.A.). Previo a la liberación en estado silvestre y con el fin de reducir el estrés de captura y la dispersión en el grupo de borrego cimarrón, se construyó, al pie de la Sierra Maderas del Carmen, un corral de adaptación de 1 ha con un cerco de 2.4 m de alto con doble línea eléctrica para evitar el ingreso de depredadores. El grupo de borregos se confinó por seis días de climatización, suplementados con agua y alfalfa. Los venados buras se mantuvieron en cautiverio cinco días en otro corral de climatización de 4 ha con un cerco de 2.4 m de alto ubicado a 10 km del corral de los borregos en el valle oeste de la Sierra Maderas del Carmen, también se les brindó agua y alfalfa, y se liberaron sin presión humana. El monitoreo de los individuos reubicados se realizó con ayuda de un receptor TR-4 (Telonics Inc., Mesa AZ., E.U.A.) durante los primeros seis meses después de su liberación con una intensidad de tres a cinco veces por semana.
Una vez en vida silvestre, al recibir señal de mortalidad se localizó al individuo lo antes posible para investigar las causas de muerte. La mortalidad a causa de puma se determinó por las características únicas de esta especie en su ataque que incluye: arrastrar y cubrir a su presa con material vegetal, arena, piedras o el mismo pelo de la presa, así como marcas de colmillo profundas en cuello o nuca, extracción del rumen, huellas de patas, excretas, marcas en suelo, y cuando termina de alimentarse de los órganos y carne, continua con las costillas, huesos de la cabeza y fémur (Krausman & Ables, 1981; McKinney, 1996; Rominger et al., 2004).
Se calculó la tasa de supervivencia y depredación por puma por medio de micromort (Heisey & Fuller, 1985; Haynes, Rubin, Jorgensen, Botta, & Boyce 2000; Logan & Sweanor, 2001; Kamler et al., 2002; Rominger et al., 2004). Este método se utiliza para estimaciones de tasas de supervivencia y causas específicas de muerte en animales con radio trasmisores, donde las tasas son estimadas por el número de días trasmitidos, el número de mortalidades debido a una causa en particular, y el número de días en intervalo de tiempo. La causa específica de muerte por puma se obtuvo de dividir el número de mortalidades por puma entre el total de mortalidades durante el estudio (Heisey & Fuller, 1985).
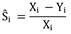
Dónde: Ŝᵢ es La tasa de supervivencia, Xᵢ total de número de días transmitidos y Yᵢ número total de muertes en intervalos de tiempo i.
Una vez obtenido el cálculo de las tasas mensuales de depredación por puma en los ungulados y con base en la prueba de normalidad Kolmogorov-Smirnov se utilizó una prueba no paramétrica de Wilcoxon 95 % I.C. en el programa estadístico PAST, donde Ho = no hay diferencia en selección de presa y Ha = sí hay diferencia en selección de presa.
Previo a la translocación y con el fin de demostrar la hipótesis, se estimaron las densidades de los ungulados antes de la liberación en Sierra Maderas del Carmen, en el mes de octubre 2014. Con base en el comportamiento, ciclo biológico de cada especie y el acceso al terreno, se empleó diferente metodología para cada especie. El censo de borrego cimarrón se realizó a través de conteo físico (Simmons & Hansen, 1980; Ross, Jalkotzy, Festa-Bianchet, 1997). Se fijaron ocho estaciones de observación en áreas representativas para el borrego cimarrón, los cuales se recorrieron con una cuatrimoto por la mañana (07:00 h-10:00 h) y por la tarde (16:00 h-19:00 h), localizando a los individuos por medio de telemetría y observación directa con ayuda de unos binoculares Zeiss© 8x42 y monocular Sawarovski© modelo ATX 20-60x85, se contó el total de borregos observados y se dividió entre el máximo ámbito hogareño 59.05 km² (Velázquez, 2012). En el caso del venado bura, se realizaron conteos nocturnos con ayuda de luz artificial en tres transectos fijos de 400 m aproximadamente de ancho, que en total suman 16.8 km² aproximadamente (Villarreal, 1999; Lancia, Kendall, Pollock, & Nichols, 2005).
Resultados
A 12 borregos (10 hembras y 2 machos) y a 10 venados bura hembras se les incorporaron collares de telemetría. De noviembre 2014 hasta abril 2015 se registraron 1 241 días transmitidos en los 12 borregos y 1 328 días transmitidos en las 10 venadas bura. La causa específica de muerte por puma en borrego fue 88 % (8/9), la tasa promedio diaria de supervivencia en borrego cimarrón fue de 0.9914 ± 0.0016 SD, la tasa promedio mensual de supervivencia fue de 0.79 y en la tasa mensual por depredación por puma tuvo rangos de 0.17 a 0.30 (Cuadro 1). En venado bura, la causa específica de muerte por puma fue 83 % (5/6). La tasa media de supervivencia diaria en venado bura fue de 0.9944 ± 0.0043 SD, la tasa media mensual de supervivencia fue de 0.86 y la tasa de depredación mensual por puma tuvo rangos de 0.10 a 0.17 (Cuadro 1). En las densidades, se estimó 0.77 borrego/km² y 7.0 venado/km². En el análisis de comparación entre las tasas de depredación por puma usando la prueba de Wilcoxon 95 % I.C., encontramos una diferencia significativa (Z = 1.826; df = 6; P = 0.05) siendo el borrego cimarrón la presa más selecta y de menor densidad.
Cuadro 1 Tasas de supervivencia y mortalidad en Ovis canadensis y Odocoileus hemionus con radio collares en Sierra Maderas del Carmen
| Mes/año | Días transmitidos | Tasas diarias en Ovis canadensis y Odocoileus hemionus | Tasas mensuales en Ovis canadensis y Odocoileus hemionus | ||
|---|---|---|---|---|---|
| Supervivencia | Depredación por puma | Otra causa de muerte | Supervivencia | Depredación por puma | Otra causa de muerte |
| Nov/2014 | 360 300 | 0.994 0.996 0.005 0.0034 0 0 | 0.83 0.90 0.17 0.10 0 0 | ||
| Dic/2014 | 310 279 | 0.990 0.996 0.009 0.0034 0 0 | 0.70 0.88 0.30 0.12 0 0 | ||
| Ene/2015 | 217 248 | 0.990 0.991 0.009 0.0080 0 0 | 0.71 0.75 0.29 0.25 0 0 | ||
| Feb/2015 | 140 196 | 0.992 0.988 0.007 0.006 0 0.006 | 0.80 0.66 0.20 0.17 0 0.17 | ||
| Mar/2015 | 124 155 | 0.991 1 0 0 0.008 0 | 0.75 1 0 0 0.25 0 | ||
| Abr/2015 | 90 150 | 1 1 0 0 0 0 | 1 1 0 0 0 0 |
Discusión
Se encontró que las tasas de supervivencia y causa de muerte por puma tienen valores parecidos entre el borrego cimarrón y venado bura, por lo que se demostró que el puma no discrimina entre especies de ungulados que comparten el mismo hábitat. Logan & Sweanor (2001) mencionan que el puma no premedita la selección de presa, sino que, como depredador oportunista, toma ventaja de la vulnerabilidad y densidad de la presa. Sin embargo, al comparar la depredación mensual entre estas especies se observó una diferencia significativa, siendo el borrego cimarrón la especie más selecta y de menor densidad, impactando en el establecimiento del grupo. La estrategia anti-depredador del borrego cimarrón es más efectiva al acoso por lobos que al asecho por pumas, por lo que la expansión de los pumas debido a la extirpación del lobo ha resultado en una trampa ecológica para los borregos donde los cambios abruptos inducidos por el hombre sobrepasan el tiempo de adaptación evolutiva de la especie (Rominger, 2018). La depredación por puma en especies de baja densidad también ha sido reportada en la Patagonia Chilena por Elbroch & Wittmer (2013), donde identificaron pumas que seleccionan especies raras y de menor abundancia en un sistema multi-presa impactando la recuperación de la población de las presas menos abundantes. Resultados similares fueron reportados por Schaefer et al. (2000), donde demostraron que la depredación por puma ha disminuido poblaciones nativas de borrego cimarrón (taxón en vías de extinción; Villepique et al., 2011; Romiger, 2018) en San Bernardino, California y también por Logan & Sweanor (2001), quienes encontraron que el mayor factor limitante en la población de borrego cimarrón de San Andres, Nuevo México, fue la depredación por puma, al grado que en 1997 fue declarado biológicamente extirpada por causas de depredación por puma, en ambos lugares los borregos cimarrones y los venados buras son simpátricos.
Algunos autores como Leopold & Krausman (1986), Kamler et al. (2002), Rosas-Rosas, Valdez, Bender, & Daniel (2003), Holl, Bleich, & Torres (2004) y McKinney et al. (2006a) sugieren que el puma cambia a presa alternativa (borrego cimarrón) cuando su presa principal (venado bura) disminuye en abundancia. Sin embargo, en el caso de este estudio, la población de venado bura es 9x mayor que la del borrego cimarrón y se demostró que las causas específicas de muerte por puma son cercanamente similares, por lo que el puma no necesita una disminución de su presa principal para cambiar a presa alternativa.
En el primer día de la liberación se observó al grupo de borregos liberado unirse con algunos individuos del grupo fundador en su rango de invierno que es de baja elevación o pie de montaña (Wehausen, 1996) hábitat que comparten con los venados bura en Sierra Maderas del Carmen. El puma como depredador oportunista, toma ventaja de la densidad y vulnerabilidad de la presa (Logan & Sweanor, 2001) y se asume que, como resultado inmediato de la translocación, se aumentó el número de presas en el área que resultaron más vulnerables por la desorientación en su nuevo hábitat, lo que favoreció a los pumas, ya que aparte de las ocho mortalidades por puma del grupo del 2014 se encontraron tres hembras depredadas del grupo fundador. Resultados semejantes fueron encontrados en Sierra Catalina, Arizona donde una primer translocación de borrego cimarrón en el 2013 se reportaron 30 mortalidades de las cuales 15 fueron a causa de depredación por puma, en los primeros cuatro meses de la liberación (Rominger 2018). La liberación de adaptación cumplió el objetivo de mantener al grupo unido ya que el tamaño del grupo se asocia con un menor riesgo de depredación (Mooring, 2004) debido a que los borregos cimarrones viven y se alimentan en conjunto como comportamiento anti-depredador (FestaBianchet, 1991; Ross et al., 1997; Logan & Sweanor, 2001). Sin embargo, no redujo la depredación por puma; Dickens, Delehanty, & Romero (2010) mencionan que después de una captura, manejo y transportación el siguiente proceso es frecuentemente el cautiverio previo a la liberación y esta última fase en algunos animales silvestres puede causar estrés crónico afectando significativamente la habilidad de escapar del depredador en los siguientes días de la liberación. En diferentes translocaciones de borregos cimarrón los pumas representan uno de los principales factores de decesos al poco tiempo después de la liberación (Chow, 1991; Rominger et al., 2004; McKinney et al., 2006b; Janke, 2015.
En uno de los sitios de depredación de borregos por puma, se encontraron los restos de una hembra con radio collar junto a un macho de la misma liberación identificado por el arete y a 1 km al norte aproximadamente fueron localizadas dos hembras más, depredadas por puma en un mismo cañón a pocos metros una de la otra. Por la intensidad, el área y el tiempo de depredación se asume que se trata de un puma especialista. Registros de pumas especialistas han sido reportados por Hoban (1990), Ross et al. (1997), Logan & Sweanor (2001), Ernest, Rubin, & Boyce (2002).
La abundancia de venado bura se debe considerar como un factor potencial en la influencia de depredación por puma sobre los borregos cimarrones (McKinney et al., 2006b) a que las poblaciones de puma no persisten en lugares donde las densidades de venado son menores a 1 venado/km² (Cashman et al., 1992), se sugiere en el futuro, realizar translocaciones de borrego cimarrón en áreas de bajas densidades de venado bura y por ende baja presencia de puma, ya que el decline de las poblaciones de borrego cimarrón, debido a depredación por puma ha sido reportada en casi todos los estados y provincias donde estas especies coexisten (Rominger, 2017). Esto podría dar una posible solución a la hora de seleccionar el hábitat potencial para futuras translocaciones de borrego cimarrón, debido a que, la depredación por puma puede generar el efecto Allee (dependencia inversa de densidad; Mooring, 2004; Bourbeau-Lemieux, Festa-Bianchet, Gillard, & Pelletier, 2011). Esto afecta la dinámica de la población, haciendo al grupo susceptible al colapso por la temprana depresión en las tasas de crecimiento, reduciendo la posibilidad del establecimiento de la población de borregos, incluyendo hábitat histórico, calidad de hábitat y terreno de escape, ya que se sospecha que la abundancia de venado bura en rangos de montaña sirve como atrayente para los pumas (Wakeling et al., 2009) y puede permanecer en áreas donde el borrego cimarrón es simpátrico con el venado bura en densidades abundantes (Schaefer et al., 2000; Wakeling et al., 2009).

