











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkEl género PrestoniaSchaus, 1920 es monotípico y endémico de México; por su distribución restringida y rareza ha sido objeto de escasos estudios. Se conocen pocos ejemplares en colecciones y su distribución comprende tres pequeñas áreas disyuntas del Pacífico mexicano: Mazatlán, Sinaloa (localidad tipo), región del Istmo en Oaxaca y en el presente trabajo ampliamos su área al estado de Michoacán (Luis-Martínez, Llorente-Bousquets, & Vargas-Fernández, 2005; Llorente-Bousquets, Luis-Martínez, & Vargas-Fernández, 2006). Hasta el momento se desconoce su planta de alimentación larval, pero se supone que puede ser una Mimosaceae (Leguminosae), pues todas las especies de géneros próximos a Prestonia se alimentan de especies de plantas de esta familia (Murillo-Ramos, Hernández-Mejía, & Llorente-Bousquets, 2016).
Vázquez (1956) analizó el estatus taxonómico de Prestonia clarki, con base en ejemplares de la Mixtequilla, Oaxaca; también transcribió las descripciones originales a nivel genérico y específico de Schaus (1920), ubicando al género en la tribu Rhodocerini sensu Klots (=Coliadinae). Lo diferenció del género Phoebis Hübner, [1819] por su vena humeral muy corta, pues en Prestonia no existe; además de otras diferencias genitales, siguiendo a Klots (1931-1933): en Phoebis el saccus está curvado suavemente, el tegumen tiene los procesos articulares largos y delgados, las valvas presentan un lóbulo interno setífero redondeado o bien una espina dorsal fuertemente quitinizada o un proceso dentado; el anellus es delgado y es ausente el subscaphium. Mientras que en Prestonia el saccus presenta dos curvaturas fuertemente marcadas, el tegumen es largo y grueso, en las valvas no existen lóbulos, espinas ni procesos dentados; el anellus es grueso y presenta subscaphium. No obstante, situó a Prestonia como un subgénero de Phoebis, dentro de un total de cuatro subgéneros: Prestonia, Phoebis, Aphrissa Butler, 1873 y Rhabdodryas Godman & Salvin, 1889. Así mismo, redescribió con detalle a Prestonia clarki, e ilustró los genitales masculinos, femeninos y su venación alar. Dicha autora mencionó, como característica propia de Rhodocerini, al tercer artejo de los palpos como muy corto; basándose a su vez en la descripción original de Prestonia (Schaus, 1920), quien había destacado que era cónico y con muchas escamas. De acuerdo con Vázquez (1956), los palpos de machos están cubiertos de escamas de color amarillo estroncio, con el tercer artejo de igual color café que en la cabeza; del género Phoebis indicó que el tercer artejo es corto y oval.
Maza (1975) describió la forma ‘semimaculata’, en la que ambos sexos presentan diferencias menores en color, bajo su reverso alar. Miller y Brown (1975), sin referir el trabajo de Vázquez (1956), también efectuaron una redescripción de P. clarki, incluyendo los genitales masculinos; ellos hicieron énfasis en las similitudes y diferencias con respecto a los subgéneros de Klots (1931-1933), ubicando a Prestonia -por sus caracteres genitales- en un estatus intermedio. Su análisis los llevó a las mismas conclusiones que obtuvo Vázquez, en relación con su categorización subgenérica dentro del género Phoebis. Lamas (2004) reconoció a Prestonia como un género monotípico de Coliadinae.
Braby, Vila y Pierce (2006) en su análisis molecular de la familia Pieridae, con base en cuatro genes (COI, EF-1α, 28S, y wingless), no consideraron a Prestonia como uno de los géneros de Colianidae que estudiaron. De acuerdo con Murillo-Ramos et al. (2016), lo consideraron implícitamente en sus resultados dentro del género Phoebis. Hernández, Flores-Gallardo y Llorente-Bousquets (2014), al estudiar el corion de P. clarki bajo las Coliadinae, encontraron que Prestonia exhibe estados de carácter coriónico notablemente plesiomórficos. Murillo-Ramos et al. (2016), en su análisis sobre las relaciones del género Aphrissa confirmaron que el género Prestonia es el grupo hermano del clado (Aphrissa + (Rhabdodryas + Phoebis)), con base en caracteres morfológicos (alares, coriónicos y de genitales).
Excepto el trabajo de Vázquez (1956), ninguno de los autores mencionó los palpos labiales de Prestonia. No obstante, en diversos grupos de lepidópteros tales estructuras han sido estudiadas a nivel ultraestructural desde los años 1970s; aunque se han abordado pocos taxones de mariposas diurnas durante este lapso. Los estudios se circunscriben a morfometría, ontogenia, electrofisiología y respuestas de las sensilas u organitos relacionados con la quimiorecepción de los palpos labiales, principalmente el órgano Palp pit (ver adelante).
Los palpos labiales son apéndices muy conspicuos de las partes bucales de adultos de Lepidoptera; están cubiertos con numerosas escamas y contienen gran número de sensilas (Krenn, 2010). La presencia de sensilas dentro de estructuras llamadas ‘Palp pit Labial’ (LPO), se caracteriza por poseer superficies ranuradas con poros o túbulos porosos en su pared, que es típico de receptores. Estos Palp pit pueden variar desde pequeñas depresiones con pocas sensilas hasta estructuras especializadas con gran número de estas (Kent, Harrow, Quartararo, & Hildebrand, 1986).
De acuerdo con Lee, Seizer y Altner (1985), los palpos labiales de Papilionoidea se componen de tres artejos: el basal, unido al labium, el mesial y el distal. Es en el último donde se localiza el Palp pit. La mayoría de autores se refieren a las partes que componen los palpos como ‘segmentos’, algunos como Chen y Hua (2016) y Kawahara, Ortiz-Acevedo y Marshall (2012) los denominan ‘palpómeros’, pero nosotros empleamos el término más general artejos, por tratarse de partes articuladas de apéndices bucales.
Reuter (1896) propuso un esquema jerárquico entre diferentes familias de mariposas, definido por la morfología de los palpos labiales. Para Pieridae describió 31 géneros, mencionando las dimensiones y describiendo con detalle las formas y curvaturas de cada artejo, así como las proporciones entre cada uno de ellos; además de la ‘mancha basal’, los ‘conos’ y las escamas en la superficie. Un detalle importante es que todos los géneros de piéridas estudiadas exhiben tres artejos, excepto Pseudopontia paradoxa Staudinger, 1884 (Pseudopontiinae), que solo exhibe dos.
Entonces, dos estructuras u organitos en los palpos están bien definidos y pueden ser de importancia taxonómica: mancha de Reuter y órgano Palp pit. La primera es mencionada e ilustrada en estudios microestructurales comparados por autores como Bilotta (1992) para varias especies de Morphini. Mielke, Mielke y Casagrande (2004) la estudiaron también en Charaxinae: Zaretis itys itylus (Westwood, 1850) y Agrias claudina anneta (Gray, 1832). Otros autores, además de esquemas, incluyeron micrografías electrónicas de barrido y breves descripciones, v. gr. Miller (1987); en Papilionidae, Duarte, Casagrande y Mielke (2001) en Hemiargus hanno (Stoll, 1790) (Polyommatini); Paluch, Casagrande y Mielke (2008) en Actinote melanisans Oberthür, 1917 (Acraeinae); Dias, Casagrande y Mielke (2010) en Memphis moruus stheno (Prittwitz, 1865) (Charaxinae); Leite, Casagrande y Mielke (2010) en Heraclides anchisiades capys (Hübner, [1809]) (Papilionidae), y Sourakov, Duehl y Sourakov (2012) en Morpho helenor (Cramer, 1776) (Morphini).
Recientemente se estudió el órgano Palp pit a nivel ultraestructural, en especies consideradas plagas de cuatro familias de Lepidoptera: Noctuidae, Tortricidae, Pyralidae y Carposinidae. Todos utilizaron microscopios electrónicos, el MEB y/o el de transmisión, describiendo los tipos encontrados, el número de sensilas y su ordenamiento. Sin embargo, ninguno de estos trató con especies de mariposas (Zhao et al., 2013; Dong et al., 2014; Barcaba & Krenn, 2015; Chen & Hua, 2016; Song, Sun, & Wu 2016).
Antes los estudios sobre palpos labiales en Pieridae se centraron en algunas especies de Pierinae de importancia económica, entre ellas están: Pieris rapae (Linnaeus, 1758) (Lee, 1987; Lee & Altner, 1986a, b; Lee et al., 1985; Lee, Kim, & Altner, 1988; Lee, Altner, Kim, & Kim, 1989) y Pieris brassicae (Linnaeus, 1758) (Lee & Altner, 1986b; Bogner, 1990); pero también en varias especies de Heliconiinae (Nymphalidae) (Krenn & Penz, 1998). En otras Nymphalidae, los palpos labiales se estudiaron con propósitos filogenéticos y evolutivos (v. gr. Miller, 1968; Salazar, 2008), no solo estudios descriptivos.
Las sensilas son las unidades sensoriales que componen los organitos; su variedad y especialización se han determinado según la posición en el artejo o región donde se hallan. Respecto a los palpos se han encontrado sensilas celocónicas (Faucheaux, 1999; claviformes sensuLee et al., 1985; Zhao et al., 2013), campaniformes (Duarte et al., 2001; Barcaba & Krenn, 2015; Song et al., 2016), quéticas sin poros (Bogner, Boppré, Ernst, & Boeckh, 1986; Faucheux, 2008; Chen & Hua, 2016; Song et al., 2016), celocónicas multiporosas (Faucheux, 2008; Salazar, 2008; Barcaba & Krenn, 2015), escuamiformes (Chen & Hua, 2016; Song et al., 2016), aplanadas (Chen & Hua, 2016), tricoides y de forma de hoja (Barcaba & Krenn, 2015) y microtriquias (Bogner et al., 1986; Chen & Hua, 2016). En el Glosario (Apéndice) se describen estos tipos de sedas según los trabajos de Sellier (1977), Ljungberg y Hallberg (1992), Krenn y Penz (1998), Faucheux (1999), Faucheux, Kristensen y Yen (2006), Kristensen, Scoble y Karsholt (2007), Shields (2008), Krenn (2010), Faucheux (2013), Barcaba y Krenn (2015), Chen y Hua (2016), Ma, Chang, Lu, Lei y Yang (2016) y Song et al. (2016).
El presente estudio describe e ilustra a nivel ultraestructural los palpos labiales de imagos de Prestonia clarki, la organización de las sensilas en sus tres artejos, particularmente en la mancha sensitiva de Reuter y el órgano Palp pit.
Materiales y métodos
Disección y Fotografía: Se diseccionaron los palpos labiales de seis ejemplares (cuatro ♂♂ y dos ♀♀, todos en el Museo de Zoología, Facultad de Ciencias, UNAM, con los siguientes datos: MÉXICO. Oaxaca: Puente Chipehua, Santo Domingo Tehuantepec, 1♂ MZFC- LEP 372544, 31/07/2012, A. Luis, Bosque Tropical Caducifolio (BTC), 50 m. Oaxaca: Parque Nacional Huatulco, Santa María Huatulco, 100 m, BTC: 1♂ MZFC- LEP 375257, 16/03/2013, J. L. Salinas; 1 ♀ MZFC- LEP 373756, 22/10/2012, A. Arellano; 1 ♀ MZFC- LEP 375253, 16/03/2013, J. Llorente. Michoacán: Rancho Nuevo, Arteaga, MZFC- LEP sin/números, 1♂ 31/05/1998, 1♂ 19/01/1998) que pasaron en humefacción en una cámara o recipiente con papel absorbente y agua. La disección se efectuó bajo un microscopio estereoscópico Olympus SZX9 a 20x de magnificación; se desprendió primero la cabeza, por medio de pinzas finas Rubis Switzerland 5-SA y después la proboscis; luego se procedió a retirar los palpos ayudados con una jeringa de insulina (BD Ultra-fine U-100 0,3 mL, 31G X 6mm), a manera de aguja de disección fina y un pincel miniatura Rodin M27 como instrumento de sujeción. Los palpos se almacenaron en seco dentro de microviales rotulados con su clave de identificación en la tapa. Se eligió un ejemplar de cada sexo para limpiar sus palpos derechos; para lo cual se colocó cada palpo en una caja de Petri PYREX de 10 cm y se eliminaron las escamas en alcohol al 50 % por medio de pinzas Dumont Dumostarl (0209-4-PO, Style 4), cambiando varias veces el alcohol hasta donde fue posible quitar la mayoría de ellas. En ocasiones el palpo también se lavó con agua destilada y se sumergió en hipoclorito de sodio al 50 % durante 2-3 min, observándose bajo el microscopio hasta aclararse las escamas que no se pudieron retirar con las pinzas más finas. Nuevamente se lavaron y se fotografiaron sumergiendo el palpo limpio en alcohol en gel, para evitar el movimiento durante las tomas. Se efectuaron medidas con el ocular micrométrico calibrado del microscopio; los palpos sin escamas se fotografiaron con la cámara Olympus DP12 integrada a un microscopio SZX12, con el objeto de efectuar el esquema. Los palpos sin escamas se conservaron en microviales de tapa plana de 0.2 ml, en alcohol al 70 %, rotulados con su clave en la tapa. Las fotografías de la cabeza y palpos se tomaron en una cámara Nikon D700, con lente AF-S Micro Nikkor 105 mm.
Preparación y observación en Microscopio de Barrido: Para la parte ultraestructural se preparó el palpo de la siguiente manera: se desprendieron la cabeza y la proboscis del ejemplar seco o humedecido; observando al estereoscopio, poco a poco se retiraron las escamas, con pinzas muy finas del lado derecho y sujetando la cabeza con cuidado por el otro lado. El ejemplar se desecaba en el procedimiento, por lo que debía volver a colocarse intermitentemente en la cámara húmeda y alternar con otro ejemplar hasta retirar la mayoría de escamas de los tres artejos del palpo. Durante el procedimiento fue necesario hacer cortes del artejo distal para poder apreciar las sensilas contenidas en su interior; para ello se humedeció el palpo y, bajo el microscopio óptico, primero se hizo un corte transversal y luego se aisló ese artejo. Después se hizo el corte longitudinal sobre el portamuestras de aluminio de 15 mm, con una navaja para afeitar marca Gillete (super delgada), compuesta de acero inoxidable revestido con politetrafluoretileno.
Otra técnica adicional que se siguió fue la de punto crítico (Bozzola & Russell, 1992), para utilizar el palpo sin escamas que se conservó en alcohol. Las estructuras fueron montadas con pinzas finas en portamuestras de aluminio de 15 mm con cinta de carbono de doble cara (8 mm); se cubrieron con oro en una ionizadora Q150R. Luego se observaron en un Microscopio Electrónico de Barrido (MEB) Hitachi S3500 bajo el detector de electrones secundarios a diferentes magnificaciones, en el Museo de Zoología de la Facultad de Ciencias de la UNAM. Algunas micrografías se tomaron en un MEB FEI Apreo field emission, en la Smithsonian Institution. El formato de las micrografías es en tiff a 2560 pixeles. En la descripción de las sensilas se tomó en cuenta a Faucheux (1999) y Shields (2008).
Resultados
Microestructura de los palpos labiales: Los palpos labiales de Prestonia clarki constan de tres artejos diferentes en forma y dimensiones (Figura 1). Al ser los palpos estructuras articuladas tubulares dirigidas hacia la parte frontal (Figura 2, Figura 3), poco esclerosadas y comprimidas lateralmente, exhiben dos vistas o caras, la externa y la interna; esta última es la que se encuentra en contacto con la proboscis y además integra mayormente a las estructuras sensoriales. Aunque en la vista externa también se localizan series de sensilas, con un arreglo en forma de hileras longitudinales en cada artejo, implican otro tipo de diferenciación sensorial por el tipo de sedas.
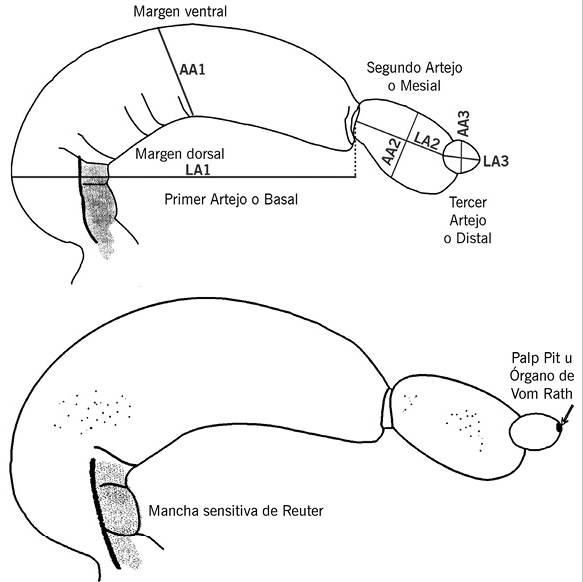
Figura 1 Vistas laterales internas del palpo labial derecho sin escamas. A. Hembra, se indican las claves de las distancias medidas. La nomenclatura utilizada es: AA1 = Anchura artejo basal, AA2 = Anchura artejo mesial, AA3 = Anchura artejo distal; LA1 = Longitud artejo basal, LA2 = Longitud artejo mesial, LA3 = Longitud artejo distal. B. Mancha sensitiva de Reuter, en artejo basal, y Palp pit, en artejo distal del macho.

Figura 2 Cabeza y palpos labiales. Vistas frontal y lateral. A, C. Macho; B, D. Hembra. (Fotografías de Arturo Arellano en cámara Nikon D700, con lente AF-S MICRO NIKKOR 105 mm).
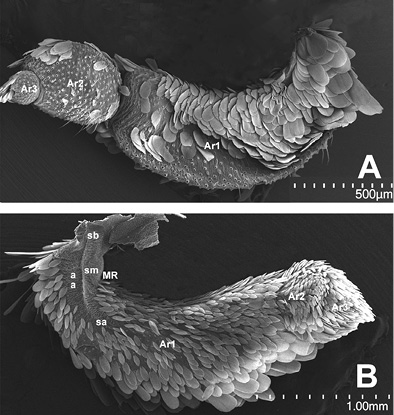
Figura 3 Palpos labiales. A. Vista externa de un palpo izquierdo completo de una hembra, al que se le han retirado la mayor parte de las escamas; B. Vista interna de un palpo izquierdo completo de un macho, con escamas. Ar1= Artejo basal, Ar2= Artejo mesial, Ar3= Artejo distal; Secciones basal (sb), media (sm) y apical (sa) de mancha sensitiva de Reuter (MR). Magnificaciones utilizadas: A - 60x y B - 42x.
La forma de los artejos basal y mesial, observados sin escamas, guardan diferentes proporciones de largo-ancho en los dos sexos. El macho tiene una anchura mayor (tomada en la parte media) y el primer cuarto basal del primer artejo se aprecia muy abultado. Ambos sexos muestran el artejo basal reni o faseoliforme, pero más alargado y arqueado en los machos. En cuanto al grosor de cada artejo se aprecian diferencias en toda su longitud: en el macho es más angosto que en la hembra. Una medida aproximada del grosor es: en machos < 200 μm y en hembras 250 μm.
En el artejo basal se observa la mancha sensitiva de Reuter y varios pliegues (Figura 3B).
El artejo mesial es elipsoidal. Este artejo asemeja una cuchara por la forma alargada y curveada hacia su interior.
El artejo distal es esferoide u elipsoide. En su interior, al microscopio estereoscópico, en el palpo sin escamas, puede apreciarse una estructura de color café en forma de botella, que ocupa aproximadamente la mitad del artejo. Esto constituye una estructura que algunos autores han llamado órgano de vom Rath y muchos otros como Palp pit, la cual se describirá en la parte de ultraestructura.
Los palpos in situ, en la mayor parte de su superficie, se encuentran cubiertos por escamas de varias formas y matices amarillos. Estas escamas se asemejan a plumas debido a su forma aplanada con ápice redondeado; por su distribución en cada artejo, se encuentran acomodadas en áreas conspicuamente definidas o circunscritas. Las vistas interna y externa del palpo se diferencian entre sí, principalmente por el tamaño y el color de las escamas.
El artejo basal ocupa la mayor parte del palpo, en una proporción de 2/3. Los artejos mesial y distal se aprecian como una sola estructura por la cobertura de sus escamas; juntos ocupan solo un tercio de la longitud total, como se muestra en la Figura 3. La vista externa del palpo mayormente presenta escamas de color amarillo; algunas de éstas combinadas con anaranjado hacia la parte apical de la escama. En su vista interna este artejo está cubierto de modo predominante por escamas alargadas y angostas de color crema, cuyo ápice es agudo (porciones media y distal); hacia el margen ventral las escamas son de mayor longitud y su ápice más romo. En el área próximo-dorsal, donde se encuentra la mancha sensitiva de Reuter, no existen escamas; en el estereoscopio (1.5 X 90 de magnificación) se reconocen proyecciones pequeñas de escala diminuta, que solo es posible observar bien a nivel ultraestructural.
La vista externa del artejo basal, en su sección proximal exhibe escamas brillantes de color blanco; algunas muy largas y angostas en su porción proximal, seguidas de otras también aplanadas, pero más anchas, del mismo color, pero con ápice romo cuadrangular. En el resto son escamas de diferente longitud, en ocasiones de ápice dentado, con tres a seis puntas u ondulaciones. En el margen dorsal del artejo, las escamas son más cortas y de matiz amarillo más claro, algunas blancas, y, conforme se acercan al margen ventral, se aprecian de mayor longitud y de color amarillo canario más intenso.
En su vista interna, la distribución y cobertura de escamas delimitan tres áreas principales: (1) la porción proximal dorsal con escamas pequeñas y cortas de color crema, que puede ocupar hasta la mitad de esta estructura, llamémosle compuesta; (2) la porción proximal-media ventral -de color amarillo, cuyas escamas se intensifican en tono conforme se acercan al margen dorsal; (3) la porción distal dorsal ocupa una franja que corre hacia el margen ventral; está teñida de escamas color café marrón, muchas de ellas alargadas y de ápice bífido. En esta sección se encuentra inmerso el tercer artejo.
En la vista externa de esta estructura se aprecian dos áreas de escamas, más o menos correspondientes a aquellas de la vista interna, solo diferenciándose porque el área proximal dorsal también es de color amarillo. En esta estructura las escamas presentan de cuatro a seis ‘denticiones’ en su ápice.
Ultraestructura de los palpos labiales:
Artejo basal. Mide sin escamas 1.5 mm de longitud (LA1), su base es de 200-250 μm, con una anchura AA1 de casi 500 (parte media); en su vista externa presenta varias series o hileras de aproximadamente ocho sensilas en sentido longitudinal, intercaladas con escamas; se calculan de 15 a 20 en total. Estas sensilas quéticas tipo 1 (sq T1) son tubulares alargadas de punta prominente, de 80-120 μm de longitud y 4 de diámetro en su base, colocadas en un alveolo o cavidad (sockets) semicirculares o dacrioides de 6 μm de diámetro (Figura 4B, Figura 4D). En nuestras preparaciones de palpos sin escamas, los alveolos de las sensilas quéticas se distinguen de aquellos de las escamas por la diferencia en la magnitud y forma del diámetro, que en el caso de las sensilas son más pequeños y la forma no es circular, como los de escamas (Figura 4B, Figura 4E). Se observan algunas más cortas, de 24 μm hacia el margen dorsal y hacia la parte distal. Estas sensilas tienen surcos longitudinales en su superficie y, por lo general, se arquean sobre sí mismas, en especial las más cortas.
La vista externa del palpo es convexa y la interna algo aplanada y ligeramente cóncava. Los alveolos de las escamas se aprecian en posición supina o inclinada y el tegumento en su parte media longitudinal es más o menos liso, mientras hacia sus márgenes dorsal y ventral se observan pequeños pliegues (Figura 4A, Figura 4B, Figura 4C, Figura 4D, Figura 4E). Además, en su superficie se aprecian algunos agujeros diminutos, de diámetro menor a una micra (Figura 4E).
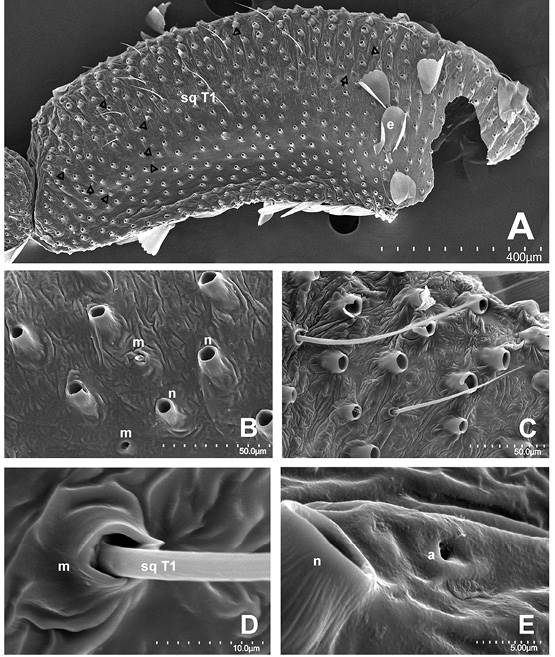
Figura 4 A. Vista externa del artejo basal del palpo derecho (puntas de flecha señalan sensilas desprendidas); sq = sensilas quéticas tipo 1, e = escama. B. Alveolos (Sockets) de sensilas (m) y alveolos de escamas (n); no se indican todas. C,D. Sensilas quéticas tipo 1 del artejo basal. E. Agujeros diminutos (poros) de la superficie del artejo basal (a). Magnificaciones utilizadas: A - 130x, B - 1000x, C - 700x, D - 5000x y E - 6000x.
La mancha de Reuter (Figura 5A, Figura 5B, Figura 5D, Figura 5F, Figura 5H) es un área alargada que integra cientos de proyecciones que hemos reconocido como microtriquias, se encuentra en la base del artejo en la vista interna con el margen dorsal, que ocupa casi un tercio de su longitud; está delimitada por un surco que corre longitudinalmente desde la base del palpo y se hace más superficial en su parte distal, hacia las 300-400 μm. Desde su parte más profunda este surco contiene proyecciones de 25-30 μm de longitud. En la mancha propiamente se observan tres secciones (basal, media y apical) y el área adyacente carente de escamas. La sección basal es semirectangular y de superficie ligeramente plana, contigua y continua con la sección media, que es abultada en su parte dorsal; cada sección mide cerca de 150-200 μm, juntas ocupan más de la mitad de la mancha. Estas dos secciones exhiben proyecciones lisas y abundantes de ápice puntiagudo o microtriquias, densamente imbricadas (Figura 5A, Figura 5B, Figura 5D); tales proyecciones del tegumento son de base circular (sin alveolo) de 2-5 μm, cuya longitud oscila desde unas pocas hasta 50 μm, ubicándose las más largas hacia la parte longitudinal media y las más cortas en la periferia. Hacia la sección apical, se intercalan escamas alargadas, elípticas y aplanadas, de ápice agudo o romo; las proyecciones que se exhiben aquí están más separadas entre sí y son más cortas conforme se acercan a la parte más distal de la mancha.
El área adyacente muestra proyecciones de longitud menor; en su mayoría se dirigen hacia el mismo sentido en el que se insertan las escamas del resto del palpo. Tales proyecciones se aprecian más espaciadas que en la superficie de la mancha, lo que permite ver la ‘topografía’ del tegumento, donde también se observan pliegues o surcos pequeños (Figura 5A, Figura 5H, Figura 5I).
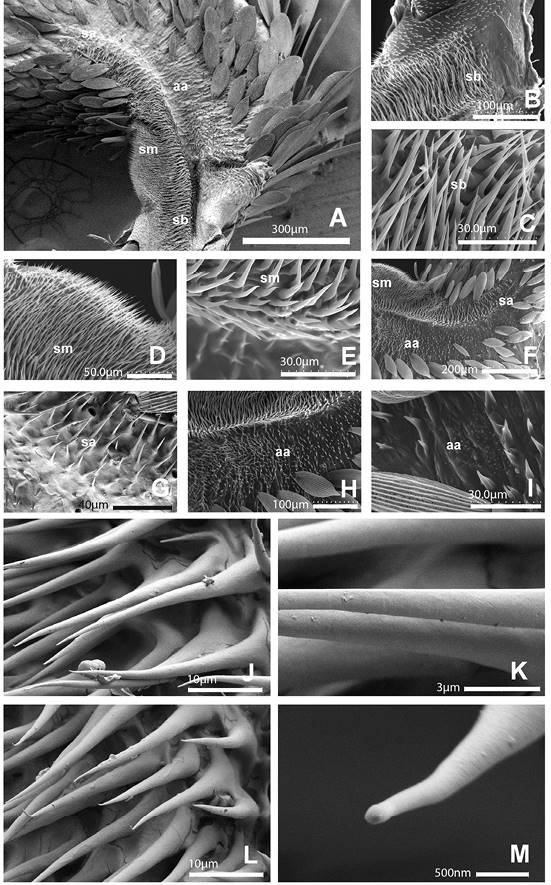
Figura 5 A-I. A. Mancha sensitiva de Reuter en vista interna del artejo basal. B. Sección basal. C. Magnificación de microtriquias, sección basal. D. Sección media. E. Magnificación de microtriquias, sección media. F. Sección apical. G. Magnificación de sección apical. H,I. Área adyacente. Secciones: basal (sb), media (sm), apical (sa) y área adyacente (aa). Magnificaciones utilizadas: A - 150x, B - 470x, C - 1900x, D - 650x, E,G - 1800x, F - 200x, H - 320x y J - 1700x. Micrografías A y G tomadas en SEM FEI Apreo field emission; el resto del Hitachi S3500. J-M. Magnificación de microtriquias de mancha sensitiva de Reuter, sección media. Magnificaciones utilizadas: J, L - 3500x, K - 12000x y M - 50000x. Micrografías tomadas en SEM FEI Apreo field emission.
Artejo mesial. Mide 0.5 mm de longitud (LA2), con ancho (AA2) de 0.35 mm. En su vista externa exhibe 5 a 6 hileras de sensilas. Cada una consta de cuatro a cinco sensilas quéticas T1 intercaladas con escamas (Figura 6A). En su vista interna se aprecia la superficie cóncava y en forma de cuchara.

Figura 6 A. Vista externa del artejo mesial del palpo izquierdo; sq T1= sensilas quéticas tipo 1. Las flechas pequeñas señalan alveolos con sensilas desprendidas. B-D, H. Sensilas quéticas con su alveolo, D y H se muestran rotas desde su base. E,F. Sensilas quéticas T1 cerca del margen y sensila ss. G. Sensila ss= escuamiforme, la cual se encuentra ex situ; se señala su posición en el artejo con flechas grandes en la fotografía A. Magnificaciones utilizadas: A - 200x, B, C - 6000x, D,H - 15000x, E - 1100x, F - 900x y G - 3200x.
En vista frontal, los alveolos donde se insertan las sensilas presentan forma dacrioide de aprox. 3-5 μm de altura y un diámetro de 4-5 (Figura 6B, Figura 6C, Figura 6D, Figura 6H).
Las sensilas quéticas T1 encontradas son de 3 μm de diámetro (Figura 6A, Figura 6E, Figura 6F).
No fue posible observar la vista interna del palpo totalmente sin escamas, ni saber con certeza si allí podemos encontrar sensilas.
Artejo distal y órgano Palp pit. El artejo distal de la hembra mide 120 µm de longitud (LA3), 150 de ancho (AA3) y una pared de 10. En el macho mide 170 de longitud (LA3). Presenta microtriquias o extensiones cuticulares del tegumento con surcos profundos, intercaladas con las escamas en sus dos vistas, entre las cuales no se aprecia diferenciación por tratarse de un artejo más redondeado y diminuto, pero en la base del artejo se aprecian más en las hembras, mientras los machos en todo el artejo. No se advirtieron muchas sensilas. Sin embargo en la zona contigua al orificio del Palp pit se aprecian varias sensilas quéticas T1 (Figura 7E, Figura 7F).
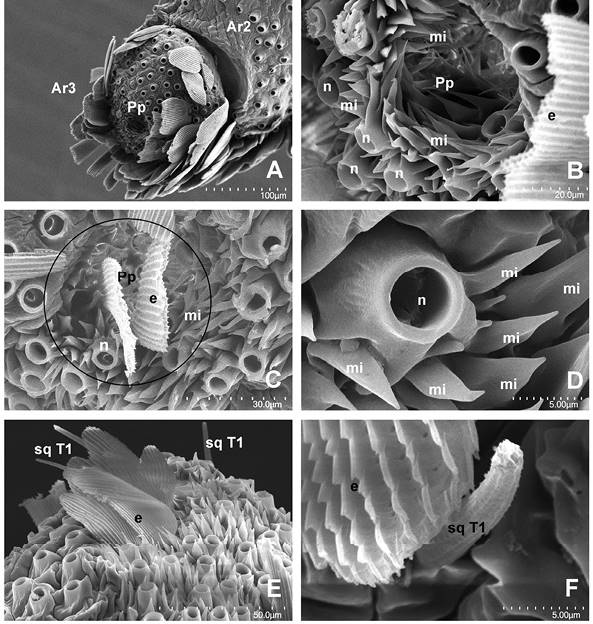
Figura 7 Artejo distal. A. Artejo completo casi sin escamas mostrando el orificio Palp pit, magnificación en B y C (círculo); Ar2= Artejo mesial, Ar3= Artejo distal, Pp= Palp pit. B, C. Vista exterior del órgano Palp pit mostrando las microtriquias (mi); n= alveolo de escama, mi= microtriquia, e = escama. D. Microtriquias y alveolo de escama cercanos al Palp pit. E. Porción superior del artejo distal; sq= sensilas quéticas tipo 1. F. Escama (e) y sensila quética T1, cercanos al Palp pit. Magnificaciones utilizadas: A - 350x, B - 2000x, C - 1600, D - 6000, E - 1100x y F - 700x.
Bajo corte longitudinal en su interior se observa el órgano Palp pit, en forma de botella con 95 μm de profundidad y 60-70 de diámetro, en su parte más amplia muestra una pared de 2 μm de grosor, cuyo orificio de entrada es de casi 20 μm (Figura 8E). Allí se encontraron dos tipos principales de estructuras: sensilas celocónicas (Faucheux, 1999; claviformes sensuZhao et al., 2013) y microtriquias (sensu Bogner et al., 1986 y Faucheux, 1999); las primeras con estrías en la cutícula, estas van desde la base hasta el ápice (Figura 8A, Figura 8C, Figura 8F). El diámetro basal de las celocónicas se observa de la misma o menor medida del alveolo semicircular donde se inserta (4-5 μm); se aprecia como una depresión en la cutícula, cuya profundidad es de aproximadamente 1-2 μm. La sensila se estrecha a 1.5-2.0 μm a nivel de la cutícula (diámetro basal) y su longitud total es de 20 μm, en promedio; todas se dirigen hacia el centro del órgano. La mayor parte de su superficie es estriada y con crestas aparentes, en su mayoría en la maza, que es cilíndrica o aplanada en el ápice y sus extremos curvados y extendidos hacia el mismo lado. De allí, en muchos casos, sobresalen dos o más proyecciones esferiformes de pocos nanómetros, sostenidas por un pedicelo de dos a tres veces el diámetro de la proyección. Este pedicelo puede encontrarse erecto o en ocasiones curvado. Las clavas presentan diferentes formas; algunas se aprecian de forma otoide (Sellier, 1977), otras algo menos aplanadas, con forma de cuchara en su ápice.
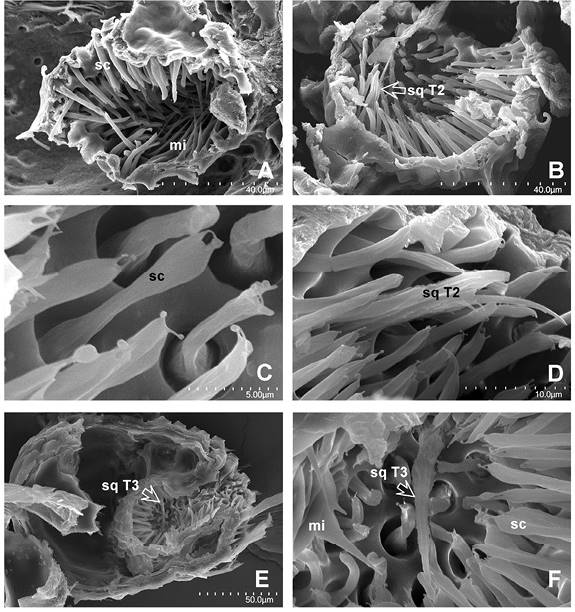
Figura 8 A-D. Corte oblicuo de un Palp pit que muestra, en A: la parte superior del Palp pit. B. Parte inferior o proximal del órgano. La flecha señala una sensila única quética T2, magnificada en D. C. Sensilas celocónicas (sc). D. Sensila quética sq T2, rodeada de sensilas celocónicas. E, F. Corte longitudinal del Artejo distal y del Palp pit (Ejemplar diferente a las fotos de A-D). Se observan tres tipos de sensilas contenidas y la forma de botella del órgano. E. Una sola sensila quética (T3) indicada con la flecha, la misma señalada en F. Magnificaciones utilizadas: A - 1300, B - 1400x, C - 8000x, D - 4500, E - 600x y F - 3500x.
Las microtriquias son alargadas, semiconiformes, con cutícula de superficie lisa y su longitud varía de 10-20 μm, con una base de 3-5 que carece de alveolo (Figura 8A); se encuentran inclinadas y se dirigen hacia la parte distal del mismo, esto es, la abertura u oquedad del artejo. La base es ancha con respecto a su parte media y su ápice muy aguzado. Las más cercanas a la superficie exterior del Palp pit se aprecian más cortas y presentan en su pared extensiones o surcos laterales longitudinales, que en corte transversal en su parte media es una cruz y no un elipsoide, como las localizadas en la parte media.
Se reconoció también un segundo tipo de sensila difícil de determinar que denominamos quética tipo 2 ó sq T2 (Figura 8B, Figura 8D); cuyo cuerpo es aprox. 50 % más amplio en su base. Posee crestas longitudinales paralelas que exhiben diminutas ornamentaciones espiniformes, las más largas son de 2 μm. En su porción distal presenta dos proyecciones o ramificaciones que terminan en punta. La base más tales ramificaciones miden casi 25 μm de longitud. Se encuentra insertada casi al centro del fondo del órgano Palp pit. Y un tercer tipo de sensila (quética tipo 3, sq T3), diferente de las anteriores y se ilustra en la figura 8E y en la figura 8F; en apariencia es semejante a la anterior, aunque de longitud mucho mayor, aproximadamente del doble de las celocónicas. Su ápice termina en clava, aunque no de la forma habitual que observamos en el resto.
La distribución de sensilas dentro del Palp pit presenta entonces el patrón siguiente: en el fondo del órgano (mitad proximal) están insertas las sensilas celocónicas, mientras en la mitad distal se hallan las microtriquias. Las sensilas celocónicas (Faucheux, 1999) se encuentran separadas entre sí por una distancia de 2-3 μm entre cada alveolo; la cantidad aproximada es difícil de calcular, aunque puede estar en el intervalo 150-200. En la parte apical exterior del Palp pit se observaron algunas sensilas quéticas sq T1 (Fig 7E, Figura 7F).
En la vista interna de los palpos, durante las disecciones al estereoscopio, se encontraron dos tipos de polen en más de uno de los ejemplares estudiados. Y un tercero en un solo ejemplar. Éstos pertenecen a plantas de las familias Acanthaceae, Polygonaceae (aff.) y Sapindaceae (Figura 9), visitadas por estos imagos que se adhirieron a los palpos posiblemente en el momento de alimentarse.
Otro organismo asociado fue el ácaro Proctolaelaps aff. antiquusTreat & Niederman (1967) (Mesostigmata: Melicharidae) (Figura 10), del que se encontró un ejemplar femenino en la vista interna del palpo de una hembra dePrestonia. Se han reportado varias especies asociadas a este género de ácaro en la proboscis de algunas noctuidas, lo que de acuerdo con Treat y Niederman (1967), sugiere que pueden habitar flores o frutos en descomposición.
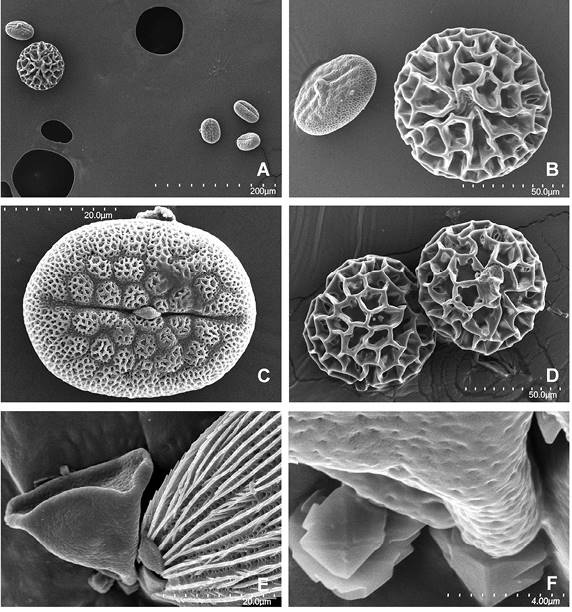
Figura 9 Granos de polen encontrados en la vista interna del palpo de Prestonia, en un ejemplar macho del estado de Michoacán, México, de las familias Acanthaceae (A, B y C), Polygonaceae (aff.) (A, B y D) y de la familia Sapindaceae (E y F), del estado de Oaxaca, México, C. Vista ecuatorial de polen diporado de cf. Justicia furcata (Acanthaceae). E, F. Vista polar de polen triporado del género Serjania. Magnificaciones utilizadas: A - 270x, B - 900x, C - 2500x, D - 850x, E - 2500x y E -1300x.

Figura 10 Ácaro hembra Proctolaelaps aff. antiquusTreat & Niederman (1967), (Mesostigmata: Melicharidae) encontrado en palpo de una hembra de Prestonia. (Fotografía de Ana I. Bieler en fotomicroscopio Olympus Provis con cámara Evolution MP de 5 Mp, captura digital Ax70).
Discusión
Con respecto al sistema propuesto por Reuter (1896), Viloria (2001, inéd.) hizo una crítica importante de sus opiniones, arguyendo que este autor contó con la suerte de haber trabajado con un número limitado de taxones, lo cual facilitó la generación de sus conclusiones. Aunque en Pieridae, la inclusión de 30 géneros es una muestra grande para tales estudios de microestructura.
Entonces si se toma en cuenta el trabajo de Reuter (1896), sabemos que la longitud del artejo mesial en los géneros de Pieridae va de 1/4 a 2/3 del basal y solo en un caso (Anthocharis Boisduval, Rambur & Graslin, [1833]) el mesial es más largo; el promedio en esta familia es casi de 1/2. Los palpos labiales en Prestonia guardan la misma proporción observada por este autor y por Miller (1987) para otras piéridas (Dismorphia Hübner, 1816, Colias Fabricius, 1807 y Pieris Schrank, 1801; con excepción de Euchloe Hübner, 1819, en este último autor) en cuanto a la longitud de sus artejos. En Prestonia el artejo mesial mide poco más de un tercio del basal y el distal un décimo. La proporción citada por Reuter (1896) en Prestonia se cumple para los dos sexos, aunque en hembras el artejo distal es un poco más pequeño y redondeado que en los machos.
Las proporciones del segundo y tercer artejo mesial, también se encontraron en 44 especies de Satyrinae (Miller, 1968). Este autor examinó varios caracteres de los palpos: a) la longitud de cada artejo; b) la relación entre la longitud del segundo y tercer artejos; c) la amplitud máxima del segundo, y d) la longitud máxima de las sedas de éste. Aunque encontró tendencias no obtuvo correlación filogenética aparente.
Miller (1987) encontró que en Troidini (Papilionidae, excepto Battus Scopoli, 1777 y Pharmacophagus antenor Haase, 1891), los palpos son bi-articulados, lo cual en su análisis definió la monofilia de este grupo. Este autor destaca que la mancha de Reuter se pierde en algunos grupos.
Salazar (2008) analizó y describió los palpos labiales de varios géneros de Charaxinae neotropicales, presentando dibujos de los palpos sin escamas y micrografías electrónicas, algunas con claridad insuficiente de las estructuras. También definió la mancha de Reuter como el área localizada en la cara interna de la unión basal cerca de la posgena, dispuesta como una placa provista de sedas erecto-pectinadas. En ocasiones describió más a detalle la forma y/o la composición de la mancha de Reuter en dos secciones y la diferenciación en el tamaño de sus sedas, que también llamó conos. Los palpos desnudos de Archaeoprepona Fruhstorfer, 1915, Zaretis Hübner, [1819] y Agrias Doubleday, [1845] que este autor ilustró muestran proporciones distintas en sus artejos respecto a los ilustrados por Miller (1987) para Papilionidae y Pieridae, donde el artejo basal es el de mayor longitud, como ocurre en Prestonia. En estos Charaxinae el artejo más largo es el mesial, y también es el de mayor curvatura. El artejo distal es el más corto. Aun con lo poco que se sabe de la mancha de Reuter, hay diferencias significativas en su composición de sedas, microtriquias y escamas que le rodean (Reuter, 1896; Miller, 1987; Duarte et al., 2001; Dias et al., 2010), además de las diferentes calidades en las tomas; por lo tanto, es difícil aún comparar al nivel famílico. Por lo tanto es recomendable estudiar un grupo de géneros próximos en Coliadinae (Pieridae), como lo hizo Miller (1987) en Papilionidae.
Como en Prestonia, en algunos casos existe dismorfismo sexual en las dimensiones de los artejos; así como ocurre en Carposina sasakii Matsumura (Carposinidae), en la que Chen y Hua (2016) encontraron que la longitud total de palpos fue significativamente mayor en hembras que en machos; el artejo mesial exhibió más del doble de longitud que en hembras (2.3 veces) y que, a su vez, es 4.63 veces mayor que el distal, y 5.04 veces que el basal. De acuerdo con Zacharuk (1985) puede pensarse que algunas diferencias entre los sexos están asociadas con diferencias conductuales, tales como el cortejo, la oviposición y el reconocimiento de su planta de alimentación larval.
El grosor de los artejos ha sido una característica poco estudiada y/o mencionada en la literatura. El artejo mesial en Prestonia es comprimido lateralmente, mientras que en algunas Heliconiinae estudiadas por Krenn y Penz (1998) es cilíndrico (Laparus Billberg, 1820, Neruda J.R.G. Turner, 1976 y Heliconius Kluk, 1780), pero en Eueides Hübner, 1816 es ligeramente ensanchado. En las Charaxinae estudiadas por Salazar (2008) también son comprimidos lateralmente y por lo tanto presentan dos caras o vistas. De acuerdo con Krenn y Penz (1998) la anatomía particular de los palpos labiales parece asociarse con los hábitos alimenticios del imago, pues mencionan que en especies frugívoras, ciertas escamas que sobresalen de los artejos basal y mesial ocultan la proboscis. Sus resultados indicaron que los palpos labiales son más cortos y angostos en especies de heliconinas que se alimentan de polen vs. las que no lo hacen. Faucheux (2013) mencionó también que la morfología de las sensilas varía de acuerdo con el tipo de alimento y el método de tomarlo. Todo esto nos sugiere que las dimensiones del palpo y/o de sus artejos no solo guardan relación con las sensilas que los componen (órganos de los sentidos involucrados), sino con lo que consumen y cómo lo hacen.
Las sensilas de la vista externa de los tres artejos en los palpos de Prestonia son quéticas tipo 1, sq T1 (Figura 4A, Figura 4D, Figura 6, Figura 7E). Chen y Hua (2016) encontraron en Carposina sasakii sensilas quéticas principalmente en el artejo distal y también los tipos escuamiforme (todo el palpo) y el campaniforme en el basal. Posiblemente las sensilas escuamiformes se encuentran en el mismo artejo de Prestonia, aunque nuestra técnica de limpieza no nos permitió diferenciarlas de las escamas; también se debió al tamaño diminuto del artejo (120 μm en hembras y 170 en machos) y la consiguiente dificultad de manipulación. Song et al. (2016) mencionaron la presencia de sensilas escuamiformes en el artejo mesial de Grapholita molesta (Busck) (Tortricidae), indicando que este tipo también se encuentra en antenas de otros grupos de polillas, pero su posible función aún se desconoce. Otros autores más antiguos (citado en Chen & Hua, 2016), refieren a este tipo de sensilas con función mecanorreceptora.
Las sensilas quéticas, de acuerdo con Shields (2008), y con Städler et al. (1974), Gnatzy y Tautz (1980) y French y Sanders (1981) (citado en Song et al., 2016) son mecanorreceptoras, aunque Altner, Sass y Altner (1977) las consideraron de función bimodal gusto-táctil. Faucheux (2013) las diferencia de las sensilas tricoides, que son multiporosas y de función olfativa, y menciona que a veces son difíciles de distinguir, por lo que algunos autores las confunden.
En el presente trabajo pudimos observar agujeros diminutos en la cutícula del artejo basal (Figura 4E), como los que Song et al. (2016) hallaron en el artejo mesial en palpos de Grapholita molesta; ellos lo mencionaron como algo novedoso y cuya función probable podría ser glandular; no se encuentran distribuidos de manera uniforme y son de un diámetro muy pequeño, menor a una micra.
Las sensilas encontradas en la base del Palp pit de Prestonia -que llamamos celocónicas, son semejantes a las que reportaron Lee et al. (1985) en Pieris rapae (Pierinae): con 20 μm de longitud, en un número de 80 y localizadas en la base del Palp pit. Al considerar que su longitud es similar en Prestonia, notamos que el diámetro basal de la sensila es mayor en Pieris (3.5 μm). Las dimensiones del Palp pit en Prestonia (95 de profundidad y 60-70 de diámetro y abertura de 20 μm) versus Pieris (100 profundidad) son similares; Lee et al. (1985) calcularon el diámetro de la abertura del Palp pit en 30 μm para Pieris, aunque ésta no se encuentra en su parte apical, sino subterminalmente por su cara ventral. También son similares las sensilas sin poros halladas por Bogner et al. (1986) en Rhodogastria Hübner, [1819] (Arctiidae; con cierta diferencia en las ornamentaciones o proyecciones apicales) y a las que Ljunberg y Hallberg (1992) estudiaron en palpos maxilares del tricóptero Apatania stigmatella Zetterstedt, 1840 (Limnephilidae), aunque con diferencia en el tamaño. La superficie de las sensilas en Prestonia -al igual que en Apatania, es estriada longitudinalmente; su sección apical puede exhibir una o varias proyecciones encontradas dentro de alveolos o pequeñas depresiones (Figura 8C, Figura 8D, Figura 8F). De acuerdo con Song et al. (2016), en Manduca sexta (Linnaeus) (Sphingidae) y algunos Tineidae, entre otros Lepidoptera, se han observado estas sensilas en el Palp pit.
Chen y Hua (2016, p. 46) reconocieron la presencia de sensilas aplanadas (flattened) de 7.5 μm de longitud en los dos tercios basales del Palp pit de Carposina sasakii, que se componen de una porción distal en forma de hoja y otra basal cilindriforme. Por sus ilustraciones pensamos que este es el mismo tipo hallado por Song et al. (2016, p. 64) en Grapholita molesta (10.5 μm de longitud). Nos parece apropiado para este tipo de sensila el término celocónica (Faucheux, 1999); posiblemente también se trata del mismo tipo estudiado en Mythima separata Walker (Noctuidae) por Dong et al. (2014), quienes la denominaron claviforme (> a 10 μm), aunque en sus figuras no se aprecian (superficie de la cutícula) las estrías tan marcadas como en los dos trabajos citados. Stange y Stowe (1999) presentaron micrografías de sensilas contenidas en el Palp pit de Cactoblastis cactorum (Berg, 1885) (Pyralidae), las que llamaron aplanadas (para nosotros, celocónicas) con su parte apical en forma de hoja; su longitud es 10 μm, similar a las descritas en otras especies; la longitud en Prestonia es casi el doble.
En Prestonia observamos sensilas celocónicas, en las que el extremo puede terminar en forma ligeramente plano y elíptico, un poco cóncavo, o bien presentan en su ápice una o más ornamentaciones o proyecciones semiesféricas (Figura 8C, Figura 8F). En algunos casos se asemejan a una cabeza de caracol terrestre, que en la literatura no encontramos algo similar, excepto en Bogner et al. (1986), quienes ilustraron y describieron un tipo semejante en Rhodogastria, sin denominarlo. Por las micrografías de sensilas con puntas bifurcadas podemos hacer esta comparación, aunque en Prestonia el ápice de la sensila es más amplio, en muchos casos aplanado y donde muestran las proyecciones en menor cantidad.
Barcaba y Krenn (2015) encontraron sensilas ‘en forma de hoja’ cerca del ápice de los palpos labiales de Plodia interpunctella (Hübner, 1813) (Pyralidae), en los bordes del Palp pit, además de sensilas tricoides. En sus ilustraciones no se aprecia alguna sensila ‘en forma de hoja’ completa pues ellos, al parecer, no efectuaron cortes longitudinales del Palp pit, pero también podríamos pensar que se trata de un tipo de sensila celocónica. Con respecto a las sensilas tricoides consideramos, al analizar sus figuras, que éste podría ser uno de los casos que Faucheux (2013) mencionó como una confusión del tipo de sensila y, en realidad, pueda tratarse de sensilas quéticas, como las del tipo 1, halladas en Prestonia (Figura 7E, Figura 7F), en el ápice del Palp pit.
De acuerdo con algunos autores como Bogner et al. (1986), Stange, Monro, Stowe y Osmond (1995), Stange y Stowe (1999) y Chen y Hua (2016) podemos sugerir las sensilas que llamamos celocónicas poseen ‘dendritas’ lameladas típicas, características de neuronas sensibles al CO2; estas sensilas poseen en su pared numerosos poros o receptores olfatorios que responden a cambios de CO2. Sobre el tema se han llevado a cabo numerosos estudios entre los que podemos mencionar: Bogner (1990), Stange (1992), Keil (1996) y otros.
Las estructuras que reconocemos como microtriquias en Prestonia (Figura 7B, Figura 8A, Figura 8E), se encontraron en la misma posición por Bogner et al. (1986) en Rhodogastria (Arctiidae) y por Chen y Hua (2016) en Carposina sasakii; esto es, localizadas en la parte superior o distal del Palp pit. Song et al. (2016) observaron en Grapholita molesta un tipo similar que también denominaron piliforme, pero algunos aparecen bifurcados en el tercio apical y las puntas no son tan aguzadas como en Prestonia, pero en nuestra opinión se trata de microtriquias. Estos últimos autores encontraron además otro tipo de estructuras en el Palp pit que llamaron mastoides, localizadas entre las microtriquias (para ellos, sensilas piliformes), cuyo diámetro fue de 0.27 μm y de forma redondeada. Zhao et al. (2013) en Helicoverpa armigera (Hübner), 1808 (Noctuidae) y Dong et al. (2014) en Mythimna separata ilustran y describen un tipo de sensila semejante que ellos también reconocen como piliforme, pero consideramos que se trata de las mismas microtriquias reportadas por Bogner et al. (1986) y Chen y Hua (2016) y que identificamos en Prestonia.
Con respecto a la mancha de Reuter, hasta el momento no tenemos descripciones ultraestructurales precisas para comparación. Reuter (1896) estudió y describió la estructura con detalle en varios taxones, refiriendo los componentes que la forman como ‘conos’. Posteriormente a este autor, algunos muestran ilustraciones y describen de manera breve o incluyen esquemas indicando su posición, forma y tamaño aproximados. Pero no se mencionan detalles del tipo de estructuras de que está compuesta la mancha; algunos términos que se utilizan son: ‘procesos espinosos’ (Duarte et al., 2001), ‘proyecciones espinosas’ (Paluch et al., 2008) o ‘pequeñas espinas’ (Moraes & Duarte, 2009); lo que si se menciona es que se trata un órgano sensitivo en su conjunto. Duarte et al. (2001) estudiaron a Hemiargus hanno y mostraron en una micrografía microtriquias lisas, largas y puntiagudas, como en Prestonia. La mancha es alargada y en su parte basal presenta las estructuras más imbricadas con otras más cortas y separadas. Paluch et al. (2008) ilustraron una micrografía de Actinote melanisans, en la que se aprecian estructuras que ellos llaman microproyecciones espinosas y espatuladas localizadas en la vista interna del artejo basal; en su mayoría son alargadas (aprox. 60-90 μm), de pared plana, bordes aserrados y ápice en punta, no muy agudo; algunas más cortas semitriangulares, en la periferia de la mancha, que son similares a Memphis moruus stheno ilustrado por Dias et al. (2010). Barcaba y Krenn (2015) describieron sensilas sin alveolo presentes en el canal alimentario de Plodia interpunctella, solo compuestas de conos sensoriales. Esta descripción podría coincidir con las estructuras que encontramos en la mancha de Reuter de los palpos de Prestonia, pero en su ilustración estos conos se observan insertados en pliegues o depresiones de la cutícula, colocados en hileras separadas. Sin embargo, coinciden en la carencia de alveolo pues se trata de microtriquias.
Las descripciones hechas por Duarte et al. (2001), Paluch et al. (2008), Moraes y Duarte (2009), Dias et al. (2010) y Leite et al. (2010) sobre la mancha o sus componentes indican que sus objetivos fueron más generales, ya que aunque se ha estudiado, su diferenciación y el contenido de sedas identificadas es escaso, y las figuras presentadas hasta ahora por estos autores, no permiten aún conclusiones más claras como referencia para comparación con el presente estudio. Sus trabajos no sugirieron una función específica para la estructura del conjunto ni de sus partes, excepto Hallberg, Hansson y Löfstedt (2003), quienes la describieron como un conjunto denso de pelos cuticulares (para nosotros microtriquias), los cuales no están inervados, y que, según mencionan, pueden presentar agregaciones de sensilas campaniformes y escuamiformes entre ellos; estos autores se basaron a su vez en Faucheux (1999) y en sus propias observaciones, hasta ese momento inéditas. Aunque podemos mencionar que, de acuerdo con las observaciones de DeVries (1979, 1987), la conducta que involucra el movimiento de los palpos labiales a través de los ojos es la de limpieza.
En Prestonia logramos identificar que no se trata de una estructura tan uniforme y por su posición creemos que se encuentra en contacto directo con la proboscis, aunque, por ahora no podemos determinar su función, hasta efectuar estudios morfoestructurales y fisiológicos más profundos.

