











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkTourism development is an important driver of forest fragmentation in some countries in tropical areas, in addition to the historic clearing for cattle and agriculture (Bierregaard & Stouffer, 1997; Lambin, Geist, & Lepers, 2003). Construction of tourism developments and associated infrastructure (golf courses, residential zones, recreational parks, roads, etc.) result in fragmentation of forest habitats (Fahrig, 1997; Christ, Hillel, Matus, & Sweeting, 2003; White et al., 2012), leaving many different shapes and sizes of forest remnants. Further, selective extraction of native vegetation and introduction of exotic species to increase the value of tourism complexes (Chettri, Chandra, Sharma, & Jackson, 2005; Schlaepfer, Sax, & Olden, 2011), modify plant species composition, and forest structure and complexity (vertical stratification and plant species composition). Altogether, these environmental modifications reduce the availability of habitats with suitable attributes (e.g., food resources and shelter) to forest-dependent wild fauna, including bird communities (McGarigal & McComb, 1995; Newsome, Moore, & Dowling, 2002; Buckley, 2004).
In addition, if the number of remnants increases, distance between them increases and the exposed edge becomes larger (Fahrig, 1997; Sodhi, 2002; Sekercioglu, 2007), resulting as plausible scenario a higher mortality of bird species by high nest predation as well lower food availability near to the edge of remnants with respect to their interior (e.g., Whyte, Didham, & Briskie, 2005; Newmark & Stanley, 2011). However, these effects depend on the attributes of avian community such as: migratory status, feeding guilds, species richness, and abundance (Stouffer & Bierregaard, 1995; Bierregaard & Stouffer, 1997); as well as forest type and the local threats facing each of them. Nevertheless, some bird species are able to use forest remnants surrounded by secondary growth, in a matrix with pasture and crops and other land uses, with stable population sizes and even experiencing significant increases in their populations (Hughes, Daily, & Ehrlich, 2002; Sekercioglu, Loarie, Oviedo, Ehrlich, & Daily 2007). Thus, this biodiversity corresponds to species generalists or species associated with anthropogenic activities (Krauss et al., 2010). Forest-interior bird species (i.e., specialist species) abilities to use the matrix of modified habitats surrounding forest fragments may affect their vulnerability in fragmented landscapes i.e. species that avoid the matrix tend to decline or disappear in fragments, while those (i.e., generalist bird species) that tolerate or exploit the matrix often remain stable or increase. However, it is not known what happens in a tourism development where forest remnants are interspersed by residential buildings and tourism activities, which are increasing across the tropical forest in Latin American.
During the period 2000-2010, world tropical forest deforestation was 62 % (Keenan et al., 2015), resulting in 6.5 million hectares lost per year. However, in Mexico showed the largest deforestation rates, with 197 651 hectares lost from the 2001 to 2015 period (see details in http://www.globalforestwatch.org/country/ MEX). Tropical forest originally covered about 8 % of the country, being considered a world “hotspot” because of its high biodiversity and endemism (Myers, Mittermeier, Mittermeier, Da Fonseca, & Kent, 2000). Unfortunately, this ecosystem has experienced high deforestation rates, particularly since the early 1970´s, because of conversion to pastures and crops, and the establishment of tourism development. Nonetheless, it is still possible to find considerable amount of tropical forest in the Yucatan Peninsula. However, these tropical forest area consist of forest remnants surrounded by mosaics of agricultural land, tourism development and secondary growth. Therefore, it is very important to know the characteristics and extension of these remnants of tropical forest and evaluate if it possible to conserve bird diversity and richness compared to other areas with different land uses. In order to know if the tropical forest remnants are functioning as bird diversity shelters within a matrix dominated by tourism development in one of the most important tourism area in Mexico, our goal was to better understand differences in bird species richness among natural and modified habitats in Akumal region in Quintana Roo, Mexico. In addition, to investigate the role of the different habitat types in a matrix dominated by tourism development. This study aims to provide a general understanding of how bird communities are affected by tourism development. We expected to find a lower species richness and a distinctive bird species composition in modified environments compared with natural environments (mature tropical forest and tropical forest remnants).
Materials and methods
Study area: The present study was carried out in Akumal, an area with several tourism developments (covering approximately 143 km2) located in the Yucatan Peninsula between 20°30’ N - 87°12’ W & 20°10’ N - 87°26’ W (Fig. 1), at the municipality of Tulum in Quintana Roo, Mexico. This site ranges in elevation from 0 to 20 masl, climate of warm subhumid type with abundant rainfalls in summer. Annual average temperature ranges from 25 to 28 °C, and annual precipitation between 1 300 and 1 500 mm. Dominant natural vegetation in the area is tropical semideciduous forest, tropical deciduous forest, and tropical flooded forest associated with secondary growth; as well as relicts of dunes coast vegetation and mangrove. Common tree species in the study area included Brosimum alicastrum, Bursera simaruba, Manilkara zapota, Talisia olivaeformis, Metopium brownei, Caesalpinia gaumeri, Thrinax radiata, Coccothrinax readi and Pseudophoenix sargentü. The mangrove relicts founded are dominated by Rhizophora mangle and Laguncularia racemose. From the tide line, where the sand accumulates and the soil is very unstable plants, are established Sesuvium portulacastrum, trailblazers like Ambrosia hispida, Salicornia and Hymenocallis littoralis bigelavii. This vegetation is the limit to stable dunes where there is a thicket forming shrub species complex as Cocoloba uvifera, Ipomoea pescaprae, Camavelia rosea, Sophora tomentosa, and Ernodea littoralis, among others (Miranda, 1959; Rzedowski, 1978).
Fig. 1 Map show the location of study area in Yucatan Peninsula, Mexico. The location of study area is found in the eastern Yucatan Peninsula, and it’s delimited by a square that it’s shown in detail in the right panel.
Habitat classification: Habitat classification was based on main vegetation cover, land uses, and the pattern of utilization by settlements as follow: (a) mature tropical forest: tropical semideciduous and tropical deciduous forest >2 ha with mature trees >10 years with canopy height 8 to 15 m and, diameters >20 cm; (b) tropical forest remnants: tropical semideciduous and tropical deciduous forest remnants <2 ha with mature trees <10 years, with canopy height 4 to 8 m and, diameters <20 cm within golf courses and residential zones; (c) modified environments by tourism developments, that include golf course and artificial water bodies in golf course; hotels zones and residential with natural and introduced vegetation; (e) modified environments by urban developments, crops and livestock, that include urban zone with natural and introduced vegetation, cattle pastures and agricultural fields; (f) coast dunes, beach zone and small remnants mangrove.
Bird surveys: Point counts surveys were conducted along transects in the different habitat types (see above; Hutto, Pletschet, & Hendricks, 1986) from April 2009 to November 2010, for a total of 412 point counts in 96 days. Points were randomly selected to represent different types of natural vegetation and land uses in the area (107 km2). Distance between sampling points were at least 250 m to avoid double-counting of highly local species (Hutto et al., 1986; Ralph, Saber, & Droege, 1995). Observation time by point was 20 min, as proposed for tropical environments (Vielliard, 2000). Points were located in both edge and interior of the forest remnants. Sampling was conducted monthly mostly in the morning (06:00 to 11:30 h) and in the afternoon (15:30 to 20:00 h), additional to nocturnal observations. Birds were identified by sight and sound (mostly), excluding birds that overflew the sampling points. Sampling was avoided on rainy days. Species richness was expressed as the total number of species recorded in each habitat, because effort was approximately equal at all habitats (21 days of sampling effort per habitat, with exception of coast dunes, beach zone and small remnants mangrove, which was 12 days). For species identification, Peterson and Chalif (1989), and Howell and Webb (1995) guides were used, and nomenclature and taxonomic status followed AOU (2017), as well as some supplements.
Bird attributes: Birds were categorized as resident or migratory species according to their presence during the study period and complemented with Howell and Webb (1995). Feeding habits were categorized according to which the species was feeding most frequently, which was complemented with literature sources (Peterson & Chalif, 1989; Howell & Webb, 1995) and field observations: omnivores, nectarivores, carnivores, frugivores, granivores, and insectivores (included aquatic invertebrates as well as bark insectivores, aerial insectivores, trunk insectivores, generalist insectivores, ground insectivores, and leaf insectivores). Habitat use preferences were categorized based on Blair (1996), and based on main cover vegetation of the land uses (see above).
Statistical analyses: Species richness was calculated as the cumulative number of species observed in the study area. EstimateS v.9 was used to compute species accumulation curves for the species detected by survey (number of sampling days) (Colwell, 2013). Species accumulation curves estimate the number of species expected in the study area and to compare qualitatively avian richness among habitat types, based on randomized re-sampling from all pooled samples. Asymptote from species accumulation curves was constructed by MichaelisMenton species richness estimation function using EstimateS v.9 (Colwell & Coddington, 1994). This method estimates of total species richness based on successively larger numbers of samples from the data set. Non-parametric estimator Jackknife 2 was selected based for having the slightest bias in the accuracy data (Walther & Moore, 2005; Hortal, Borges, & Gaspar, 2006). The Shannon diversity index (H´) and Simpson´s index (D) were obtained to estimate diversity among habitats (Krebs, 2000). Point Abundance Index (PAI) was calculated by dividing the number of detections for each species by the total number of point’s sampled (Blondel, Ferry, & Frochot, 1970). To understand how community composition differs, and what species are present and how the habitats differ in the mix of species they have, we conducted a hybrid multidimensional scaling ordination (HMDS), using the Bray-Curtis dissimilarity index on untransformed species abundance. The hybrid MDS was introduced by Faith, Michin and Belbin (1987) and combines both the PCoA (principal coordinate analysis or classical MDS) and the non-metric MDS (NMDS). It has the advantage of assuming a linear relationship between the ecological distances obtained by the ordination and the dissimilarity measures where it is most often straight (the PCoA part), and only monotonicity where ecological distances (in the ordination space) are too high to be accurately measured (the NMDS part; Faith et al., 1987). Differences between natural environments and modified environments (see above) were tested using a permutation multivariate analysis of variance (PERMANOVA; Anderson, 2005). Data of the coast dunes, beach zone and mangrove were not included given the low number of sampling points made in those areas. All analyses were conducted using Minitab (see details http:// www.minitab.com/).
Results
We recorded a total of 1 914 bird sightings during the study period, with a bird density of 54.3 individuals/observation-hour. A total of 160 species and 50 families was recorded, from which only five species are considered endemic, and 10 species were most frequently recorded (Appendix). Accumulation curves for sampling by census reached an asymptote (Fig. 2A) in the value of 170 species. In this context, Jackknife´s 2 estimator resulted in a value of 177 species, indicating that the probability of encountering more species increasing sampling effort is very low (Fig. 2B). From all detected bird species, 99 were permanent residents, 47 were winter visitors, 11 were transients, and three introduced (Appendix). The avian community in the study have a predominance of insectivore species (N= 97, see Appendix).
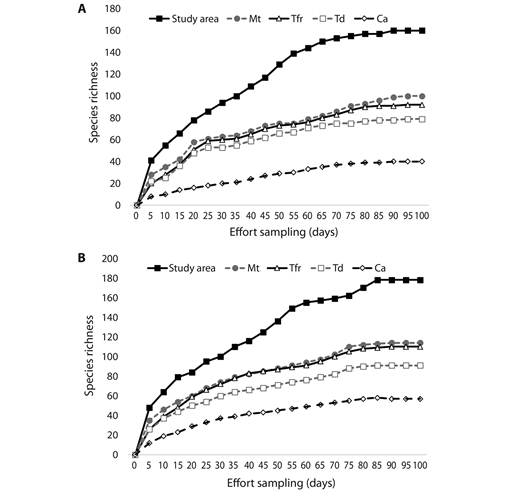
Fig. 2 Species accumulation curve for birds sampled by census in the study area. Observed species richness (a) and expected (b) in the study area and habitat type: Mature tropical forest (Mt), Tropical forest remnants (Tfr), Modified environments by tourism developments (Td), and Modified environments by urban developments, crops and livestock (Ca).
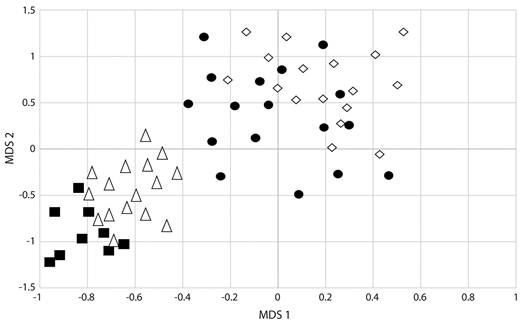
Bird community attributes by habitats: A total of 96 species were found in mature tropical forest, 92 in tropical forest remnants, 79 in modified environments by tourism developments and, 40 in modified environments by urban developments, crops and livestock (Appendix). Accumulation curves showed that the expected species richness present in mature tropical forest had the highest bird richness (Jackknife 2 = 114), followed by tropical forest remnants with expected species (Jackknife 2 = 110), modified environments by tourism developments (Jackknife 2 = 91), and (Jackknife 2 = 57; Fig. 2B). Only 17 species were exclusively found in mature tropical forest, three in tropical forest remnants, and the rest was shared, and four in modified environments by tourism developments and two in modified environments by urban development, crops and livestock, while the rest were found in two to four habitat types (Appendix). The species richness and diversity values were highest in mature tropical forest (96 species, H‘ = 3.78±0.006, D= 0.93±0.010) and tropical forest remnants (94 species, H‘= 3.32±0.008, D= 0.90±0.010); while, modified environments by tourism developments (72 species, H‘= 2.89±0.014, D= 0.73±0.030), and modified environments by urban developments, crops and livestock (40 species, H‘= 2.73±0.012, D= 0.69±0.029) presented the lowest species richness and diversity values (Fig. 3). Bird species richness and diversity values (H‘, D) varied significantly among habitats (Fig. 3; P< 0.001), with few species detected in modified environments compared with mature tropical forest and tropical forest remnants. This was supported also by the HMDS ordination explained 55 % of the variation in species composition among habitats. Clear gradients in community composition were observed along both axes, with the centroids for mature tropical forest sites and tropical forest remnants having negative values on both axes and the centroids for modified environments (by tourism developments and by urban developments, crops and livestock) having positive values (Fig. 4). Mature tropical forest and tropical forest remnants sites were clearly separated from modified environments, and grouping of same land-use sites was significant (F2 = 28.632, R2 = 0.412, P< 0.05) showing that bird richness differ significantly between the four analyzed habitats (Fig. 4). Similarly, with migratory status, the highest species richness of resident and migratory species was recorded in mature tropical forest (69 resident species and 26 migratory species) and tropical forest remnants (62 resident species and 35 migratory species) while modified environments by tourism developments (45 resident species, 26 migratory species, and one introduced species) and modified environments by urban developments, crops and livestock (30 resident species, eight migratory species, and two introduced species) recorded the lowest species richness; but not significant difference depending on migratory status, both migratory and resident species respond the same way. Insectivore species were better represented in the mature tropical forest, and tropical forest remnants (>16). Frugivores and nectarivores species were slightly higher and abundant in mature tropical forest and, tropical forest remnants (with six species in each habitat). Carnivores (18), granivores (10) and omnivores (10) species were better represented in modified environments (particularly in cattle pastures and agricultural fields), and insectivore species were better represented in mature tropical forest (65 species) and, tropical forest remnants (59; Appendix). Results in this study are consistent with respect to that modified environments (i.e., agricultural and livestock areas) had a higher proportion of carnivores and granivores species in comparison with tropical forest.
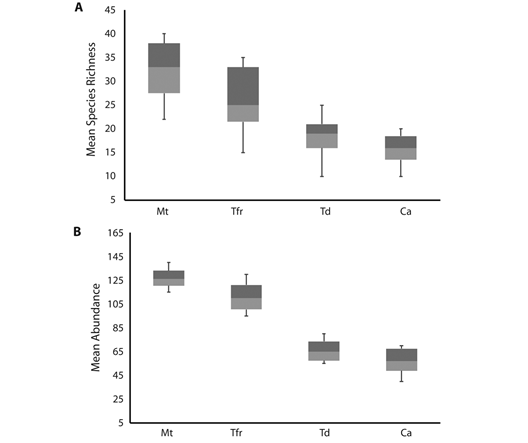
Fig. 3 Boxplots of mean species richness (a), mean abundance (b), (c) Shannon diversity index, and (d) Simpson´s diversity index of bird species at Akumal region, Quintana Roo, Mexico: Mature tropical forest (Mt), Tropical forest remnants (Tfr), Modified environments by tourism developments (Td), and Modified environments by urban developments, crops and livestock (Ca). Lines represent minimum, first quartile, median, third quartile, and maximum.
Discussion
Our study revealed that mature and tropical forest remnants in Akumal region had higher bird species diversity that the modified environments, which is expected because modified environments lack suitable vegetative remnants, shrubs and canopy cover that limits food density and diversity, nest placement, and predator avoidance. Above mentioned reveals the importance of tropical forest remnants for bird diversity conservation in a tourism area, as an important shelters to the bird community. According with our results, Bennet and Saunders (2010) mentioned that the forest remnants are very important in terms of shelter, feeding and nesting areas, particularly to birds that depend on native vegetation.
Bird species recorded accounted for 32 % of all species reported for Quintana Roo by Correa and MacKinnon (2011), being the order Passeriformes the most representatives with 52 % (83 species) from the total recorded. Abundance index values (PAI) showed a large number of species with low PAI, as well as few species with intermediate to high PAI compared to the pattern observed in other surveys (Aleixo & Vielliard, 1995; Lyra-Neves, Martins, Mendes, Rodrigues, & Lacerda, 2004). Bird species richness in the study was similar to other tropical forest areas with a predominance of insectivore species (e.g., Estrada, CoatesEstrada, & Meritt, 1997; Blake & Loiselle, 2001). Omnivore species abundance can be directly related to the variety of available resources for change in land use and declining native resources like fruits. However, the presence of frugivore species, also some bark insectivore species indicate that the study area is relatively well conserved (Blake & Loiselle, 2015). Others signs of relative adequate habitat conditions included the occurrence of mixedspecies flocks (Stotz, Fitzpatrick, Parker, & Moskovits, 1996), and trunk insectivores. Frequency and structure of mixed-species flocks also suggests habitat conditions at the study area were adequate for many common in tropical forest bird species according to Stotz et al. (1996). Most bird species recorded in this study were dependent on forest edge, these results suggest that the sensitivities of bird species to vegetation are associated with their dependence of food resources as availability of native fruit (Hasiu, Gomes, & Silva, 2007).
The differences in the species richness and diversity found in this study indicated that the mature tropical forest and tropical forest remnants present greater diversity and richness compared with modified environments. This accords with other studies in tropical environments, and indicates that the loss of original habitats directly influences the presence, abundance and persistence of species (Kattan, Álvarez-López, & Giraldo, 1994; Laurance & Bierregaard, 1997; Rocha, Virtanen, & Cabeza, 2015). The higher avian diversity found in tropical forest may be due to high numbers of individuals and mature vegetation that provide many different microhabitats, which promote varieties of bird species compared with habitats with different land covers (e.g., with human infrastructure or tourism development). However, others studies have found highest richness in modified environments than natural environments (Petit, Petit, Christian, & Powell, 1999; Martin, Viano, Ratsimisetra, Laloë, & Carrière, 2012), but this may be due to the environmental heterogeneity that can get to present the area.
Tropical forest remnants had a significant contribution to the bird species richness and diversity in the study area which is consistent with those reported by Estrada et al. (1997) in Los Tuxtlas region in Veracruz, Mexico. On the other hand, bird composition in terms of the feeding guilds is related to vegetation structure (Laurance & Bierregaard, 1997). Different groups of bird species were found that respond differently to the conversion of forest to modified environments. Not surprisingly, tropical forest assemblages were characterized by a high proportion of forest-associated species, whereas modified habitats were dominated by generalists and open habitat specialists. However, modified environments by urban developments, crops and livestock are very important to a lot of carnivores, granivores and insectivores species because of temporarily or permanently provide such resources depending on their phenology and seasonality (Loiselle & Blake, 1994).
In general, the tropical forest remnants that presents the study area appears to contribute to the relatively high species richness, especially considering the number of species occurring in mature tropical forest. Results of this study showed evidence that tropical forest remnants are significantly important in tourism zones as an available habitat for birds. The continuing expansion of tourism complex, particularly large-scale, will likely result in the simplification and loss of bird diversity. That is particularly important in tourism zones from Quintana Roo because these remnants representing shelters, feeding or nesting areas for birds dependent from natural environments; as well as responsible for maintaining an important proportion of regional avian diversity. The importance of tropical forest remnants provides important habitats for many species of resident and migrant birds in Yucatan Peninsula. Our results confirm the great need for conservation (preserved areas), restoration with native vegetation, and ecological studies of tropical forests remnants, because represent the first step to take actions for conservation of regional avian diversity in the Yucatan Peninsula subjected to anthropogenic activities. An added potential value to this tourist area to attract other tourism type (as birdwatchers) as an alternative to preserve and promote ecological tourism. Furthermore, create incentives for protection and preservation on natural areas and, native biota, which allow preserve these tropical forest remnants.

