











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkLas características estructurales de los ecosistemas acuáticos tropicales tienen influencia sobre la composición de las comunidades ícticas. La estructura de las comunidades ícticas en los ríos, se han relacionado principalmente con el tamaño del río, la velocidad, el caudal, la profundidad, la depredación y la competencia (Zaret & Rand, 1971; Bussing & López, 1977; Angermeier & Karr, 1984; Greenberg, 1991; Harding, Burky, & Way, 1998; Martin-Smith, 1998; Stewart, Ibarra, & Barriga-Salazar, 2002), variables que cambian a nivel espacial y temporal.
Adicionalmente, la variabilidad temporal y la heterogeneidad del hábitat (como la composición del sustrato, ancho de la cobertura vegetal, complejidad estructural y la disponibilidad de nichos), influyen en los patrones de distribución de las comunidades de peces (Vannote, Minshall, Cummins, Sedell, & Cushing, 1980); por tal razón, los hábitats de las comunidades ícticas son variables y espacialmente diversos (Frissell, Liss, Warren, & Hurley, 1986).
Las comunidades de peces son, entonces, un indicador primario de la salud e integridad de un río, de tal forma que la declinación en la calidad de las comunidades es un indicador de una influencia humana negativa para el sistema (Mueller & Pyron, 2010) como la disminución considerable del caudal debido a la creación de presas, o sequías producidas por la desviación del cauce para el riego de cultivos; o procesos físicos y biológicos que actúan como agentes de perturbación, como tormentas, inundaciones y sequías (Sousa, 1984). Para comprender estas comunidades es necesario conocer su abundancia y distribución, así como su relación con factores bióticos y abióticos que afectan sus cambios poblacionales (Oliveira & Pessanha, 2014).
Algunas variables físicas de los ríos se relacionan directamente con parámetros fisicoquímicos, como ejemplo de ello, el caudal y el flujo del agua se asocian a la temperatura y pH, las cuales pueden limitar la distribución y abundancia de las especies (Resh et al., 1988; Power, Sun, Parker, Dietrich, & Wootton, 1995), y pueden fluctuar a nivel temporal y espacial.
En el departamento del Tolima los estudios ícticos han estado encaminados a evaluar aspectos de ecología reproductiva y trófica de las familias Trichomycteridae, Characidae, Sternopygidae, Cichlidae, Astroblepidae y Loricariidae en las microcuencas de los ríos de Coello y Prado (Villa-Navarro & Losada-Prado, 1999; Villa-Navarro & Losada-Prado, 2004; Briñez-Vásquez, Villa-Navarro, Ortega-Lara, Reinoso-Flórez, & García-Melo, 2005; García-Melo, Villa-Navarro, Reinoso-Flórez, & Ortega-Lara, 2005; Zúñiga-Upegui, Villa-Navarro, Ortega-Lara, & Reinoso-Flórez, 2005), y en las microcuencas de los ríos Coello, Totare, Alvarado y Opia se han realizado estudios de diversidad, composición y estructura de las comunidades ícticas (CORTOLIMA, 2004; CORTOLIMA, 2007; López-Delgado, 2013; Albornoz-Garzón & Conde-Saldaña, 2014); no obstante, en la microcuenca del río Anchique no se han realizado estudios ícticos. Por lo anterior el objetivo del presente estudio fue describir la estructura y composición de la comunidad íctica a lo largo del río Anchique y probar la hipótesis de que estas varían a nivel espacial y temporal.
Materiales y métodos
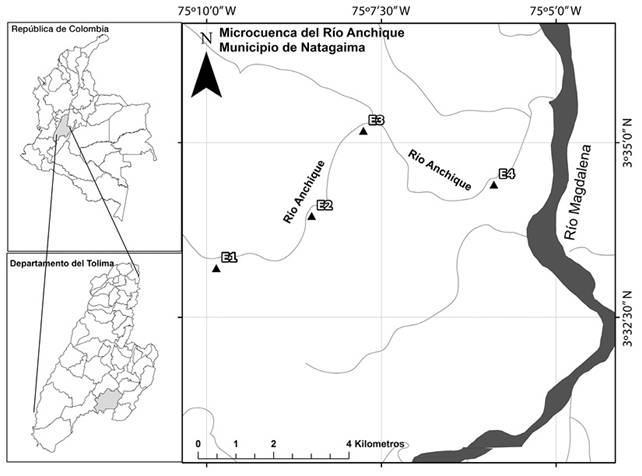
Área de estudio: El estudio se desarrolló en la cuenca del río Anchique, el cual tributa directamente al río Magdalena, y se localiza en la zona de vida bosque seco tropical (bs-T), departamento de Tolima, Municipalidad de Natagaima, Colombia. Su cuenca drena un área de 25 828.4 has y tiene una longitud de cauce de 47.4 km.
Fase de campo: Se efectuaron dos muestreos en el 2014, uno en temporada de lluvias altas (febrero) y otro en lluvias bajas (abril). Se establecieron cuatro estaciones de muestreo a lo largo del cauce principal del río. La primera estación (E1) se ubicó cerca al nacimiento del río, a una altitud de 415 m (3°33’11.7” N - 75°09’49.7” W); la segunda (E2) se ubicó a 346 m de altitud (3°33’56.3” N - 75°08’43.3” W); la tercera (E3) se ubicó a 340 m (3°35’10.1” N - 75°07’54.2” W); y la cuarta (E4) se ubicó a 3 km de su desembocadura en el río Magdalena, a 333 m de altitud (3°34’28.7” N - 75°05’59.1” W), la distancia entre E1 y E4 fue de 13 km (Figura 1).
Las capturas se realizaron empleando un equipo de electropesca, en un transecto longitudinal de 100 m de largo y a 2 m de la orilla, durante 60 min. Adicionalmente, entre dos personas fue utilizada una red de arrastre (10 x 1 m, con un ojo de malla de 0.05 m) con la cual se realizaron tres arrastres sobre las orillas en cada estación de muestreo, durante dos horas.
La electropesca es el método más adecuado para las condiciones que presentan los cuerpos de agua andinos, torrentosos y pedregosos (Maldonado-Ocampo et al., 2005); es ventajosa debido a que no es selectiva, y puede capturar peces de diferentes tamaños y no causa la muerte a los organismos, la electropesca está determinada principalmente por factores biológicos como la talla del pez, y por factores físicos como la conductividad y temperatura del agua (Guerrero-Kommritz, 1997).
Los peces capturados se fijaron en una solución de formol al 10 %, posteriormente fueron determinados taxonómicamente (Eigenmann, 1922; Dahl, 1971; Harold & Vari, 1994; Buitrago, 1995; de Santana & Maldonado-Ocampo, 2005; Maldonado-Ocampo et al., 2005) en el Laboratorio de Investigación en Zoología de la Universidad del Tolima (GIZ). Una vez Identificados y cuantificados, los individuos fueron depositados en la Colección Zoológica sección Ictiología (CZUT-IC) de la Universidad del Tolima; los individuos depositados en la Colección Zoológica van desde el lote CZUT-IC 11 527 hasta 11 915 en el primer muestreo, y CZUT-IC 11 864 hasta 11 929 en el segundo muestreo.
Paralelamente, se tomaron in situ algunos parámetros ambientales, tales como: temperatura superficial del agua empleando un termómetro de mercurio; velocidad (m / s) del agua con un flujómetro (General Oceanics - modelo 2030R y 2030R6); caudal (m3 / s), profundidad (m) y ancho (m) del cauce. La profundidad media del agua en cada estación de muestreo fue calculada con base en medidas efectuadas con una varilla graduada, en intervalos de 1 m en cada una de las estaciones del río. Estas medidas sirvieron igualmente para calcular el ancho del río, posteriormente, el caudal del río, fue calculado utilizando el área y la velocidad.
Adicionalmente, se colectaron muestras de agua superficial y contra corriente para su evaluación en el Laboratorio de Servicios de Extensión en análisis químico (LASEREX) de la Universidad del Tolima. Así, se determinó el pH (unidades de pH), oxígeno disuelto (mgO2 / L), conductividad eléctrica (µS / CM), alcalinidad (HCO3 -), dureza (mgCaCO3 / L) y sólidos totales (mg / L).
La toma de las muestras de agua se realizó siguiendo los procedimientos recomendados por el Órgano de Control Ambiental, cumpliendo con la normatividad vigente. Las muestras poseían la bitácora y los controles de calidad establecidos para la toma, rotulación, preservación, embalaje y entrega al laboratorio LASEREX acreditado ante el IDEAM, de acuerdo con lo establecido por el decreto 1600 del 27 de julio de 1994 expedido por el Ministerio de Ambiente y Desarrollo sostenible.
Variables ambientales: Para conocer la composición del sustrato y la complejidad estructural del hábitat fueron creadas dos matrices, las cuales fueron construidas teniendo en cuenta la presencia y ausencia de algunos componentes en cada estación de muestreo. Para la composición del sustrato se tuvo en cuenta la presencia y ausencia de roca, arena, hojarasca, rocas grandes, guijarro, grava y lodo; y, para la complejidad estructural del hábitat, se tuvo en cuenta la presencia y ausencia de hojarasca, vegetación enraizada sumergida, perifiton, vegetación riparia arbórea y arbustiva.
Posteriormente, se hallaron los índices de composición del sustrato (S) y complejidad estructural del hábitat (S), siguiendo la metodología propuesta por Winemiller, López-Fernández, Taphorn, Nico and Duque (2008), los cuales estandarizaron el índice (S) por medio de la siguiente fórmula:
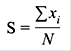
Donde, S son los índices de composición del sustrato y complejidad estructural del hábitat, xi representa cada uno de los componentes del sustrato y estructura del hábitat por cada estación de muestreo, y N representa el número de componentes combinados del sustrato o estructura del hábitat observados en las estaciones de muestreo; los valores cercanos a 0 indican mínima complejidad y cercanos a 1 máxima complejidad (Winemiller et al., 2008).
Para determinar la existencia de diferencias estadísticamente significativas entre variables físicas y fisicoquímicas de las estaciones de muestreo a nivel espacial y temporal, se realizó una prueba no paramétrica de una vía de Kruskal-Wallis. Se realizó esta prueba debido a que los datos no cumplieron los supuestos de normalidad y homogeneidad de varianza, estos fueron evaluados con la prueba de Shapiro-Wilk y la prueba de Bartlett respectivamente. Se utilizó la función Kruskal test de la librería Stats del programa estadístico R versión 3.2.5 (R Development Core Team, 2016).
Con el objetivo de identificar el régimen pluviométrico en la zona de estudio, fueron utilizados los datos de precipitaciones anuales y mensuales de los últimos diez años, con el fin de realizar un diagrama de precipitación con la media mensual multianual, y la media mensual del año de estudio (2014). Estos datos fueron obtenidos de estaciones pluviométricas cercanas a la zona de estudio del Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia (IDEAM).
Estructura y composición: Para determinar la representatividad de los muestreos, se calcularon los estimadores de diversidad Chao 1, ACE y Cole, para esto se utilizó el paquete estadístico EstimateS versión 8.2 (Colwell, 2009).
La abundancia relativa se calculó a partir del número de individuos colectados de cada especie y su relación con el número total de individuos de la muestra. Se calculó el Índice de Constancia (C), el cual indica que las especies presentes en más del 50 % de las muestras pueden ser consideradas como residentes, entre el 25 y 50 % son accesorias y, menos del 25 % son accidentales (Ferreira, 2007).
Además, fueron calculados los modelos de distribución de abundancia los cuales incorporan información sobre la riqueza, la distribución y, en algunas ocasiones, el número de especies no observadas en una comunidad (McGill et al., 2007). Estos son una herramienta útil para describir la estructura de la comunidad y pueden ser indicadores de algún tipo de disturbio (Hill & Hamer, 1998).
Para determinar la estructura de la comunidad íctica, se evaluaron las abundancias proporcionales de cada una de las especies, desarrollando los modelos serie geométrica, logarítmica, log-normal y vara quebrada de MacArthur (Moreno, 2001; Magurran, 2004); se empleó la prueba de bondad de ajuste Kolmogorov-Smirnov (Magurran, 2004) para establecer cuál de ellos explica mejor la distribución de los datos.
Para medir la diversidad a nivel espacial, se hallaron los números efectivos de especies o números de Hill, a través de la ecuación denominada qD (Jost, 2006).
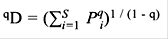
Dónde qD es la diversidad. El exponente q determina la sensibilidad del índice a las abundancias relativas de las especies. Tiene tres componentes, la diversidad de orden cero (q = 0) el cual es insensible a las abundancias de las especies, el valor obtenido es el equivalente a la riqueza de especies; la diversidad de orden 1 (q = 1) incluye a todas las especies con un peso exactamente proporcional a su abundancia en la comunidad, y tiene en cuenta las especies comunes; y, los valores de q mayores a 1 (q = 2), tienen más en cuenta las especies dominantes (Hill, 1973). Todo lo anterior se realizó haciendo uso del programa estadístico R versión 3.2.5 y la librería Vegan (R Development Core Team, 2016).
Asimismo, se realizó un análisis de rarefacción y extrapolación usando el número efectivo de especies, mediante los órdenes de diversidad q = 0, q = 1 y q = 2 (números de Hill), con el fin de identificar diferencias en la diversidad de especies a nivel temporal y espacial (Chao et al., 2014), usando el programa estadístico R versión 3.2.5 y la librería iNEXT. (R Development Core Team, 2016).
Para definir si la composición de la comunidad de peces es similar dentro de cada estación de muestreo, se realizó un análisis de escalamiento multidimensional no métrico (EMNM), usando la distancia de Bray-Curtis; el análisis se consideró robusto cuando el valor de estrés fue menor a 0.2. Se utilizó programa estadístico R versión 3.2.5 y la librería Vegan (R Development Core Team, 2016). Posterior al ordenamiento, se realizó un análisis de similitud (ANOSIM) (Chapman & Underwood, 1999) con el fin de determinar diferencias significativas entre dos o más grupos. Para este caso, se determinaron grupos a nivel espacial y temporal. Este análisis se desarrolló usando la librería Vegan del programa estadístico R versión 3.2.5 (R Development Core Team, 2016).
Influencia de los factores ambientales: Con el fin de determinar si las variables ambientales (incluyendo los valores del índice de composición del sustrato y complejidad estructural del hábitat) influyeron en la estructura de la comunidad de peces, se realizó un análisis de ordenación. El método utilizado fue el de correspondencia canónica (ACC) el cual es ampliamente utilizado para matrices que poseen datos de abundancia ya que este método utiliza la distancia de X 2 (Chi-cuadrado).
Después de realizar el análisis de ordenación, se procedió a realizar la selección de las variables, con el fin de determinar cuál de estas influyó de manera significativa en la estructura de la comunidad. Para esto, fue realizado un análisis de permutación de Monte Carlo (999 permutaciones). El ACC y la prueba de Monte Carlo fueron realizados utilizando el programa CANOCO 4.5 (Braa, Ter, & Smilauer, 2009).
Resultados
Variables ambientales: La caracterización física de las estaciones en altas lluvias mostró que el cauce fue más ancho en E2 (20 m) y menos ancho en E1 (4 m); el caudal osciló entre 2.35 m3 / s (E2) y 0.43 m3 / s (E4); la profundidad fue mayor en E1 (0.37 m) y menor en E4 (0.23 m); y finalmente, la velocidad fue mayor en E3 y E4 (0.377 m / s) (Cuadro 1). En bajas lluvias, el ancho fue mayor en E2 (18.8 m) y menor en E1 (4.1 m); el caudal osciló entre 3.86 m3 / s (E2) y 0.65 m3 / s (E1); la profundidad fue mayor en E1 (0.52 m) y menor en E4 (0.18 m); y finalmente, la velocidad fue mayor en E2 y E3 (0.69 m / s) y menor en E1 (0.30 m / s). No se encontraron diferencias estadísticamente significativas entre las variables físicas del río entre periodos climáticos (Kruskal-Wallis, profundidad p = 0.08326; ancho p = 0.1489; velocidad p = 0.2454; caudal p = 1).
Cuadro 1: Variables físicas y químicas por estación de muestreo de la microcuenca del río Anchique, Tolima, Colombia Table 1: Physical and chemical variables by sampling sites in the Anchique river sub-basin
| Variables físicas y químicas | Estación 1 | Estación 2 | Estación 3 | Estación 4 | ||||
| Altas lluvias | Bajas lluvias | Altas lluvias | Bajas lluvias | Altas lluvias | Bajas lluvias | Altas lluvias | Bajas lluvias | |
| Profundidad (m) | 0.372 | 0.52 | 0.33 | 4.12 | 0.24 | 0.3 | 0.23 | 0.65 |
| Ancho (m) | 4 | 0.29 | 20 | 18.8 | 14 | 0.69 | 5 | 3.86 |
| Velocidad (m / s) | 0.3 | 0.26 | 0.35 | 8 | 0.37 | 0.69 | 0.37 | 1.46 |
| Caudal (m3 / s) | 0.45 | 0.18 | 2.35 | 4.1 | 1.27 | 0.82 | 0.43 | 1.05 |
| pH | 7.62 | 7.68 | 7.15 | 7.44 | 7.46 | 7.12 | 7.26 | 7.56 |
| Dureza (mg CaCO3 / L) | 124.2 | 111.8 | 1 | 3 | 4.4 | 0.9 | 6.1 | 6.4 |
| Alcalinidad HCO3- | 182 | 162.4 | 42 | 34.6 | 41 | 40.3 | 45 | 48.1 |
| Conductividad eléctrica | 379 | 388 | 82.5 | 75.4 | 87.8 | 78.2 | 89.6 | 89.6 |
| Oxígeno disuelto | 6.3 | 5.55 | 6.85 | 6.29 | 7.57 | 6.36 | 7.59 | 6.33 |
| Sólidos totales | 268 | 300 | 86 | 182 | 80 | 161 | 80 | 154 |
| Temperatura del agua (ºC) | 24 | 26 | 25 | 26 | 28 | 30 | 30 | 30 |
Respecto a la caracterización fisicoquímica, en altas lluvias E1 presentó los valores más altos de pH (7.6), dureza (124.2 mg CaCO3 / L) y sólidos totales (268); contrario a esto, los valores más bajos de estas variables se hallaron en E2. La temperatura del agua fue mayor en E4 (30 °C) y menor en E1 (24 °C). En bajas lluvias, los valores de las variables de pH (7.6), dureza (111.8 mg CaCO3/ L), conductividad eléctrica (388 µs / cm) y sólidos totales (300) fueron también mayores en E1. El oxígeno disuelto osciló entre 6.33 mg O2 / L (E4) y 5.55 mgO2 / L (E1); la temperatura del agua entre 30 °C (E3 y E4) y 26 °C (E1). No se encontraron diferencias estadísticamente significativas entre las variables fisicoquímicas del río entre periodos climáticos (Kruskal-Wallis, pH p = 0.7728; dureza p = 0.7728; alcalinidad p = 0.5637, conductividad eléctrica p = 0.6631, oxígeno disuelto p = 0.08326; sólidos totales p = 0.1465 y temperatura del agua p = 0.3719).
A nivel general, el cálculo del índice complejidad estructural del hábitat (S) indicó que E4 presentó el mayor valor (0.8), seguida de E1 (0.6), mientras que E2 y E3 fueron las menos complejas (0.4); en cuanto al índice de composición del sustrato E4 presentó el mayor valor (0.72), seguida de E2 (0.43) y E3 (0.43), y finalmente, E1 obtuvo el valor más bajo (0.29).
Estructura y composición: La curva de acumulación de especies indica que el número total de especies observadas (Smean = 36) en el río Anchique corresponde al 99.31 % del valor esperado por el estimador Chao 1, 98.46 % para ACE y 100 % para Cole, indicando una buena representatividad.
Se recolectaron un total de 625 individuos, pertenecientes a cinco órdenes, 17 familias, 31 géneros y 36 especies. En lluvias altas se recolectaron 325 pertenecientes a 32 especies, mientras que para lluvias bajas se recolectaron e identificaron 300 individuos pertenecientes a 31 especies (Cuadro 2).
Cuadro 2: Lista de especies ícticas por estaciones de muestreo y época climática e Índice de Constancia de la microcuenca del río Anchique Table 2: Fish species by sampling stations, climatic period and Constancy Index of the Anchique river sub-basin
| Taxón | Estación 1 | Estación 2 | Estación 3 | Estación 4 | Índice de constancia | ||||||
| Altas lluvias | Bajas lluvias | Altas lluvias | Bajas lluvias | Altas lluvias | Bajas lluvias | Altas lluvias | Bajas lluvias | - | |||
| Characiformes | - | - | - | - | - | - | - | - | |||
| Parodontidae | - | - | - | - | - | - | - | - | |||
| Parodon magdalenesis | 0 | 0 | 0 | 0 | 5 | 1 | 0 | 0 | Accesoria | ||
| Crenuchidae | - | - | - | - | - | - | - | - | |||
| Characidium phoxocephalum | 8 | 3 | 0 | 0 | 2 | 0 | 0 | 0 | Accesoria | ||
| Characidae | - | - | - | - | - | - | - | - | |||
| Argopleura magdalenensis | 0 | 0 | 4 | 2 | 0 | 0 | 0 | 1 | Accesoria | ||
| Astyanax fasciatus | 0 | 0 | 1 | 3 | 5 | 3 | 2 | 15 | Residente | ||
| Astyanax magdalenae | 0 | 0 | 0 | 2 | Accidental | ||||||
| Creagrutus affinis | 0 | 15 | 3 | 0 | 37 | 29 | 3 | 15 | Residente | ||
| Gephyrocharax melanocheir | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 5 | Accesoria | ||
| Hemibrycon raqueliae | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 2 | Accesoria | ||
| Microgenys minuta | 0 | 0 | 1 | 6 | 8 | 7 | 0 | 0 | Accesoria | ||
| Saccoderma hastata | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 5 | Accesoria | ||
| Anostomidae | - | - | - | - | - | - | - | - | |||
| Leporinus muyscorum | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | Accidental | ||
| Erithrinidae | - | - | - | - | - | - | - | - | |||
| Hoplias malabaricus | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | Accidental | ||
| Gasteropelecidae | - | - | - | - | - | - | - | - | |||
| Gasteropelecus maculatus | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | Accidental | ||
| Siluriformes | - | - | - | - | - | - | - | - | |||
| Pimelodidae | - | - | - | - | - | - | - | - | |||
| Pimelodus blochii | 0 | 0 | 0 | 0 | 0 | 0 | 12 | 2 | Accesoria | ||
| Rhamdia guatemalensis | 0 | 0 | 0 | 0 | 2 | 0 | 1 | 0 | Accesoria | ||
| Heptapteridae | - | - | - | - | - | - | - | - | |||
| Cetopsorhamdia molinae | 0 | 0 | 0 | 1 | 16 | 11 | 0 | 0 | Accesoria | ||
| Pimelodella chagresi | 0 | 0 | 0 | 0 | 3 | 1 | 0 | 0 | Accesoria | ||
| Heptapteridae sp. | 0 | 0 | 0 | 0 | 8 | 9 | 0 | 1 | Accesoria | ||
| Cetopsidae | - | - | - | - | - | - | - | - | |||
| Cetopsis othonops | 0 | 0 | 0 | 2 | 1 | 2 | 0 | 0 | Accesoria | ||
| Trichomycteridae | - | - | - | - | - | - | - | - | |||
| Trichomycterus retropinnis | 4 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | Accesoria | ||
| Trichomycterus banneaui | 4 | 5 | 8 | 7 | 43 | 14 | 0 | 1 | Residente | ||
| Loricariidae | - | - | - | - | - | - | - | - | |||
| Chaetostoma milesi | 0 | 0 | 23 | 25 | 42 | 0 | 0 | 0 | Accesoria | ||
| Chaetostoma thomsoni | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | Accidental | ||
| Chaetostoma sp. | 0 | 0 | 0 | 0 | 0 | 34 | 0 | 1 | Accesoria | ||
| Lasiancistrus caucanus | 0 | 0 | 0 | 2 | 0 | 2 | 0 | 1 | Accesoria | ||
| Sturisomatichthys leightoni | 0 | 0 | 0 | 0 | 6 | 1 | 0 | 2 | Accesoria | ||
| Spatuloricaria gymnogaster | 0 | 0 | 0 | 0 | 2 | 3 | 0 | 1 | Accesoria | ||
| Astroblepidae | - | - | - | - | - | - | - | - | |||
| Astroblepus homodon | 1 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | Accesoria | ||
| Gymnotiformes | - | - | - | - | - | - | - | - | |||
| Sternopygidae | - | - | - | - | - | - | - | - | |||
| Eigenmaniia virescens | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 4 | Accesoria | ||
| Sternopygus aequilabiatus | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | Accesoria | ||
| Apteronotidae | - | - | - | - | - | - | - | - | |||
| Apteronotus eschmeyeri | 0 | 0 | 1 | 5 | 7 | 2 | 7 | 3 | Residente | ||
| Cyprinodontiformes | - | - | - | - | - | - | - | - | |||
| Cynolebiidae | - | - | - | - | - | - | - | - | |||
| Cynodonichthys magdalenae | 16 | 13 | 0 | 0 | 0 | 0 | 0 | 0 | Accesoria | ||
| Poeciliidae | - | - | - | - | - | - | - | - | |||
| Poecilia caucana | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 5 | Accesoria | ||
| Cichliformes | - | - | - | - | - | - | - | - | |||
| Cichlidae | - | - | - | - | - | - | - | - | |||
| Andinoacara latifrons | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 5 | Accesoria | ||
| Kronoheros umbrifer | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | Accidental | ||
| Geophagus steindachneri | 15 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | Accesoria | ||
En la cuenca del río Anchique el orden Siluriformes fue el más dominante (51 %), seguido de Characiformes (32 %). Gymnotiformes, Cyprinidontiformes y Cichliforrmes tuvieron una representación entre el 5 % y 6 %. A nivel de familias, las más abundantes fueron Characidae (28.3 %; 9 spp.), Loricariidae (23.4 %; 8 spp.) y Trichomycteridae (15.0 %; 2 spp.). Las especies más abundantes fueron Creagrutus affinis (16.3 %), Chaetostoma milesi (14.4 %) y Trichomycterus banneaui (13.1 %), que representaron más del 42.8 % de la abundancia total.
Para las dos épocas muestreadas, los órdenes con mayor abundancia relativa fueron Siluriformes y Characiformes que aportaron el 80 % de la abundancia, aproximadamente. En cuanto a familias, Characidae y Loricariidae aportaron cerca del 55 % de la abundancia en lluvias altas y bajas.
En lluvias altas, Chaetostoma milesi (20 %), Trichomycterus banneaui (16.9 %) y Creagrutus affinis (13.2 %) fueron las especies más abundantes; mientras que Leporinus muyscorum, Hoplias malabaricus, Gasteropelecus maculatus, Rhamdia guatemalensis y Chaetostoma thomsoni sólo se registraron en este periodo. Durante lluvias bajas, C. affinis (19.6 %) y Chaetostoma sp. (11.6 %) fueron las especies más abundantes; Astyanax magdalenae, Chaetostoma sp. y Lasiancistrus caucanus únicamente se registraron en esta temporada.
En general, el Índice de Constancia muestra a Astyanax fasciatus, C. affinis, T. banneaui y Apteronotus eschemeyeri como residentes (C = 11.1 %), mientras que A. magdalenae, Leporinus muyscorum, Hoplias malabaricus, Gasteropelecus maculatus, Chaetostoma thomsoni y Kronoheros umbrifer (C = 16.7 %) se consideran accidentales, y las 26 especies restantes fueron accesorias para el río (C = 72.2 %) (Cuadro 2).
En altas lluvias las especies residentes no variaron (C = 12.5 %), pero el número de especies accesorias aumentó a 28 (C = 87.5 %); en bajas lluvias nuevamente se registraron las mismas especies residentes, pero se adicionó L. caucanus (16.1 %) y bajó a 26 el número de accesorias (C = 83.8 %).
La distribución de especies del río Anchique, a nivel general, se ajustó al modelo de serie logarítmica (D36 = 0.2194; p > 0.05), el cual no varió entre las épocas evaluadas (altas lluvias, D32 = 0.0812; p > 0.05) (bajas lluvias, D31 = 0.2419; p > 0.05).
A partir de la extrapolación del número de especies (0D) a nivel espacial, se determinó que E4 (24 spp.) y E3 (20 spp.) concentran la mayor riqueza; en contraste, E2 (12 spp.) y E1 (7 spp.) fueron las estaciones con menor riqueza de especies.
A partir del número de especies típicas o comunes (1D) se evidenció que E4 (14 spp.) y E3 (10 spp.) obtuvieron los mayores valores, y los más bajos se observan en E1 (6 spp.) y E2 (5 spp.). Con respecto al número de especies dominantes (2D), E4 (11 spp.) y E3 (8 spp.) obtuvieron los valores más altos, y finalmente, los valores más bajos se observan en E1 (5 spp.) y E2 (3 spp.).
Finalmente, el número efectivo de especies en los órdenes 0 (q = 0), 1 (q = 1) y 2 (q = 2) no mostraron diferencias entre las épocas de altas y bajas lluvias evaluadas.
El análisis EMNM (stress = 0.04) muestra que la composición de la comunidad fue diferente en E1 y E4 (Figura 2). La comunidad íctica en E1 estuvo conformado por Characidium phoxocephalum, C. affinis, Trichomycterus retropinnis, T. banneaui, Astroblepus homodon, Cynodonichthys magdalenae y Geophagus steindachneri; compartiendo a C. affinis y T. banneaui con las demás estaciones, y C. phoxocephalum con E3.
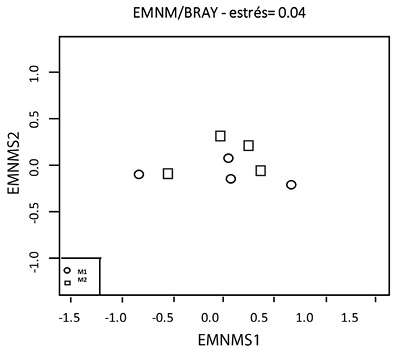
Figura 2 Escalamiento Multidimensional No Métrico (EMNM, estrés = 0.04) de la composición de la comunidad íctica en las dos épocas de muestreo evaluadas a lo largo de la microcuenca del río Anchique.
En la estación E4 se registró el mayor número de especies, mientras que en la estación E1 se presentó el menor número de especies. En este orden, la comunidad de la estación E4 se caracterizó por la presencia de A. magdalenae, Saccoderma hastata, L. muyscorum, H. malabaricus, G. maculatus, Pimelodus blochii, Eigenmannia virescens, Poecilia caucana, Andinoacara latifrons y C. umbrifera, estas especies se registraron únicamente en E4.
Las 12 especies presentes en E2 son compartidas con una u otra estación, mientras que E3, con 20 especies, tiene a Parodon magdalenensis, Pimelodella chagresi y Chaetostoma thomsoni como especies no compartidas con ninguna otra estación. Finalmente, se detectaron diferencias en las comunidades de peces espacialmente (ANOSIM, p < 0.012), no siendo así entre periodos climáticos (ANOSIM, p > 0.742).
Influencia de los factores ambientales: El análisis de correspondencia canónica, indicó que el componente 1 explicó el 33 % de la variación de las especies y el componente 2 un 27 %. El ACC y la prueba de Monte Carlo (999 permutaciones), muestran que la composición del sustrato (p = 0.022; F = 2.39), la complejidad estructural del hábitat (p = 0.012; F = 3.20) y el ancho del cauce (p = 0.052; F = 2.39) influyeron significativamente en la composición y distribución de la fauna íctica; mientras que las variables fisicoquímicas no tuvieron ninguna influencia en la composición y distribución de la fauna a lo largo del río Anchique.
La composición del sustrato y la complejidad estructural del hábitat fueron los factores que más influyeron en la comunidad de E4, en los dos periodos climáticos, asociándose con A. magdalenae, G. melanocheir, S. hastata, L. muyscorum, H. malabaricus, G. maculatus, P. blochii, R. guatemalensis, Chaetostoma sp., S. leightoni, E. virescens, S. aequilabiatus, P. caucana, A. latifrons, A. eschmeyeri y K. umbrifer (Figura 3).
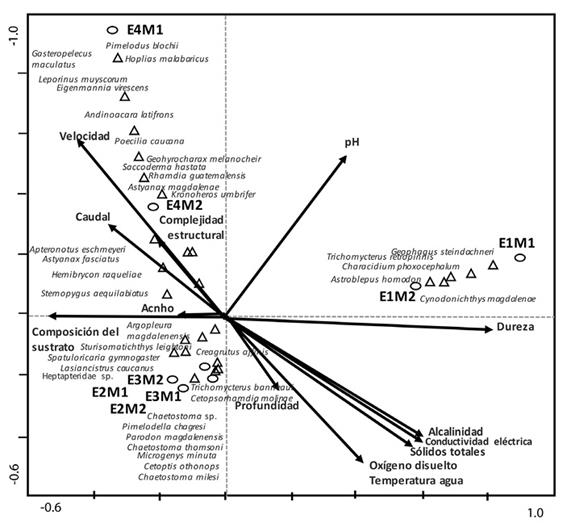
Figura 3 Análisis de Correspondencia Canónica de las especies, variables ambientales y estaciones evaluadas en la microcuenca del río Anchique.
El caudal y ancho del cauce determinaron la comunidad de peces en E2 y E3, donde se registran a A. magdalenensis, A. fasciatus, C. affinis, H. raqueliae, M. minuta, C. molinae, C. othonops, P. magdalenesis, S. gymnogaster, P. chagresi, T. banneaui, Ch. milesi y L. caucanus.
El diagrama de ordenación del ACC concuerda con el diagrama de ordenación del EMNM, en que las estaciones más similares por composición de especies, estructura de la comunidad y variables ambientales son E2 y E3 (Fig. 3), ya que se encuentran agrupadas en el mismo cuadrante del diagrama y comparten un número similar especies.
Discusión
La mayor representación de los órdenes Siluriformes y Characiformes encontrada en el presente estudio, es un patrón general no solo para la ictiofauna neotropical (Anderson & Maldonado-Ocampo, 2010) sino también para la región del Alto Magdalena, en las microcuencas de los ríos Coello (Siluriformes 54.1 %, Characiformes 34.2 %), Totare (Siluriformes 55 %, Characiformes 41 %), Opia (Characiformes 73.45 %, Siluriformes 22.1 %) y Alvarado (Siluriformes 52.8 %, Characiformes 35.3 %) (CORTOLIMA, 2004; CORTOLIMA, 2007; López-Delgado, 2013; Albornoz-Garzón & Conde-Saldaña, 2014).
El número de especies identificadas (36 especies), representan el 27 % de las especies reportadas para el Alto Magdalena (Villa-Navarro et al., 2006) y un 17 % de la cuenca del río Magdalena (Barletta et al., 2010). Adicionalmente, el número de especies registradas en el río Anchique es igual a lo reportado en ríos de mayor envergadura como el río Alvarado (36 spp.) (Albornoz-Garzón & Conde-Saldaña, 2014), y menor a lo reportado en el río Opia (38 spp.) (López-Delgado, 2013).
La comunidad íctica del río Anchique se ajustó al modelo de distribución serie logarítmica, el cual sugiere una comunidad con abundancias altamente desiguales (Hill & Hammer, 1998; He & Tang, 2008), además el modelo puede ser resultado de efectos de muestreo y efectos estocásticos o definitivos (Boswell & Patil, 1971; Hughes, 1986). Esto se ve reflejado en un tramo del río antes de E3, donde el efecto de la extracción de agua para riego (CORTOLIMA, 2007) podría generar una disminución de recursos disponibles para los peces al intervenir en los patrones hidrológicos de la cuenca y modificar los atributos físicos del hábitat, lo que podría actuar como un efecto definitivo y generando cambios en la comunidad íctica (Whittaker, 1972; Bazzaz, 1975; Gray, 1981; Nummerlin & Kaitala, 2004; He & Tang, 2008; Martins-Dias & Tejerina-Garro, 2010). Además, durante el periodo de muestreo se presentó un año atípico en el ciclo hidrológico, como efecto del fenómeno del Niño el cual actuó como un evento estocástico en la comunidad íctica del río Anchique.
Adicionalmente, el régimen pluvial bimodal del bs-T genera variaciones hidrológicas que se reflejarían en las comunidades de peces por la intermitencia en el flujo del agua, el ancho del río y conectividad entre la cabecera y los sectores medio y bajo (Winemiller, 1996; Herbert & Gelwick, 2003). Sin embargo, los resultados del presente estudio no soportan esta teoría ya que no se observaron diferencias en la composición y estructura de la comunidad íctica entre las épocas evaluadas.
La inexistencia de variación temporal entre las comunidades de peces podría explicarse por la alteración en el régimen bimodal del primer semestre de 2014. En este orden, febrero se comportó como el segundo mes más lluvioso (media = 49.3 mm, 2014 = 73.5 mm), y abril no fue el mes de mayor precipitación (media = 60.7 mm, 2014 = 49.8 mm). Lo anterior sugiere un efecto de la escala de trabajo, ya que, la fuerte relación entre la estructura de las comunidades y el régimen hidrológico no siempre puede ser verificado a escalas temporales ecológicas y espaciales locales (Herbert & Gelwick, 2003), por lo tanto, no se cumple a nivel temporal la hipótesis planteada en el presente estudio. Sin embargo, muchas especies de peces se mueven en respuesta a la variación espacial de un recurso disponible (Winemiller, 1989), lo que podría explicar las diferencias encontradas en las comunidades a nivel espacial a lo largo del río.
A lo anterior, es importante considerar que las características estructurales de los ambientes tropicales ejercen efectos en la composición de las comunidades ícticas (Mendoça, Magnuson, & Zuanon, 2005), de tal forma que la heterogeneidad del hábitat, incluyendo variaciones en el sustrato (E1, E2 y E3 con fondos de roca, guijarro y arena, y E4 compuesto principalmente de hojarasca y lodo), los cuales influyen en los patrones de distribución de las comunidades de peces tropicales (Motta-Bührnheim & Cox-Fernandes, 2003; Mueller & Pyron, 2010).
La persistencia de Creagrutus affinis y Trichomycterus banneaui podría explicarse por su capacidad de colonización de quebradas y ríos, así como la estrategia de reproducción continúa en C. affinis (Harold & Vari, 1994), y a la presencia de odontodes en T. banneaui que le permiten resistir y ascender corrientes fuertes, colonizando hábitats con fondos de roca, guijarro y corrientes fuertes (García-Melo et al., 2005, Rodríguez, 2008).
Characidium phoxocephalum, C. affinis, Trichomycterus retropinnis, T. banneaui, Astroblepus homodon, Cynodonichthys magdalenae y Geophagus steindachneri conformaron la comunidad característica de la estación E1. Se destaca que esta estación se ubica en la cabecera del río y se caracteriza por registrar un escaso número de especies, de ahí el hecho de haber presentado la menor diversidad según los números efectivos de especies.
Según las teorías del Concepto del Río Continuo (Vannote et al., 1980) y del Pulso de inundación (Junk et al., 1989), en las cabeceras y zonas altas de los ríos se presenta menor diversidad y riqueza de especies, todo esto debido a cambios estacionales abruptos que se presentan en estas zonas, este patrón se observó en el río Anchique (E1) un río corto con un gradiente altitudinal no tan pronunciado. Este hecho se puede asociar a que el río no tiene tributarios que le aporten un mayor número de especies en su parte alta.
Adicionalmente, en E1 predominaron los fondos rocosos y arenosos, con perifiton, vegetación riparia arbórea, hojarasca y detritus, siendo un hábitat o ambiente adecuado para estas especies ícticas (Briñez-Vásquez et al., 2005; Castro-Roa et al., 2007), ya que, estos elementos sirven como sustrato para el desarrollo de alevines, zonas de alimentación, refugio y reproducción (Angermeier & Karr, 1983), sin embargo, en E1 se encuentra una barrera (caída de agua) que limita el desplazamiento de las poblaciones de ictiofauna.
La riqueza local y la composición de las comunidades ícticas, tienden a incrementarse de forma directa con el ancho y volumen en los cuerpos de agua (Toepfer, Williams, Martínez, & Fisher, 1998; Grenouillet, Pont, & Herisse, 2004). Los análisis multivariados como el ACC, son consistentes con el aumento en la complejidad estructural aguas abajo, generada por la presencia de vegetación riparia, así como hojarasca y raíces sumergidas (Mueller & Pyron, 2010).
Lo anterior explicaría, en parte, la presencia E. virescens, S. aequilabiatus y A. eschmeyeri en la parte baja del río (E4), donde la vegetación enraizada sumergida puede ser utilizada para su refugio y alimentación (Ferreira & Casatti, 2006; Barreto-Teresa & Mei-Romero, 2010). Adicionalmente, E4 se encuentra ubicada más cerca a la desembocadura con el río Magdalena, lo cual facilita la migración de especies como L. muyscorum, S. leightoni y P. blochii al río Anchique, siendo uno de los factores que incrementa la diversidad en pequeños tributarios cercanos a grandes ríos (Gorman, 1986; Fernandes, Podos, & Lundberg, 2004).
El uso de números efectivos permite una mejor interpretación de la diversidad de las comunidades a nivel espacial y temporal, de acuerdo a esto, E4 registro el valor más alto de riqueza (0D = 24 spp.) y diversidad (1D = 14 spp.), lo cual se puede atribuir a la complejidad estructural del hábitat y composición del sustrato ofertado, estas características podrían ofertar más refugios y proporcionar condiciones óptimas para la colonización de un mayor número de especies ícticas (Cala, 2001). Asimismo, algunos componentes de la complejidad estructural como hojas y ramas sumergidas, sirven como sustrato para el desarrollo de microorganismos, proveen zonas de alimentación para macroinvertebrados acuáticos y peces, y ofrecen refugios contra depredadores a la ictiofauna (Angermeier & Karr, 1983; Casatti et al., 2012).
Finalmente, la comunidad de peces de la microcuenca del río Anchique no presenta variación a nivel temporal, pero sí a nivel espacial, lo cual podría deberse a que cada estación de muestreo presentó características estructurales muy disimiles entre sí, como la complejidad estructural, composición del sustrato y el ancho del río.

