











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEchinodermata present a high regenerative potential expressed at different anatomical levels in both adults and larvae (Candia-Carnevali & Bonasoro, 2001; Candia-Carnevali, 2006). This process allows echinoderms to survive to sub-lethal predation by losing some part of the body (Lawrence & Vasquez, 1996; Lawrence, Byrne, Harris, Keegan, Freeman & Cowell, 1999; Dupont & Thorndyke, 2006). It also increases their numbers rapidly through asexual reproduction and subsequent regeneration (Emson & Wilkie, 1980; Alves, Pereira & Ventura, 2002; Rubilar, Pastor de Ward & Díaz de Vivar 2005). Regeneration in this phylum is a process which may be quite rapid, depending on the structure lost or the taxa (Dupont & Thorndyke, 2006).
Brittle stars are usually subject to sublethal predation and to mechanical stress. Therefore, is common in the field to find a high percentage of individuals with several arms in process of regeneration (Pomory & Lawrence, 1999; 2001; Rodríguez 2009). The arms are used for feeding, locomotion, reproduction, social interaction (Lawrence, 1987), as well for the processes of photoreception (Dupont & Thorndyke, 2006).
In ophiuroids, the regenerative phenomena are well known and also remarkably well documented in fossils of extinct species (Aronson, 1987; 1991). However, there is a substantial gap in the understanding of regenerative mechanisms at the cellular and molecular level (Candia-Carnevali & Bonasoro, 2001; CandiaCarnevali, 2006). Arm regeneration cellular process in Ophiuroidea has been studied in two other species (Biressi et al., 2010) However, it is not clear if the regenerative mechanism at the cellular level is common for the entire class.
Ophioplocus januarii (Lütken, 1856) is one of the largest ophiuroids present in the Argentinean waters (Brogger et al., 2013). This species is found in shallow waters in both rocky and sandy bottoms (Tommasi, 1968; 1970; Thomas, 1975; Bartsch, 1982). In some coastal areas, up to 82% of the individuals usually show at least one regenerating arm. This percentage is higher than that recorded in other species (Rodríguez, 2009).
The aim of this study was to describe in detail the histological and morphological changes occurring at the surface of an arm in the ophiuroid Ophioplocus januarii after traumatic amputation.
Materials and methods
Study area and collection site
Nonregenerating individuals (n = 33) of Ophioplocus januarii with similar size (15-20 mm in disk diameter) were collected during the summer 2010 by Scuba diving at Villarino beach (42º23´S - 64º18´W), San José gulf, Argentina, at 6 m depth with a water temperature of 16ºC. Individuals were transported to a wet laboratory and acclimatized for one week. The experiment was conducted by using 501 × 40 × 50 cm (approx. 100 l). The aquarium had a constant supply of filtered seawater with a salinity of 35 ‰. An air-driven box filter containing filter floss and activated charcoal was used to maximize and filtration. The experiment was conducted under a 12 h light: 12 h dark schedule at constant temperature (15 ºC).
Experimental design for morphology observation
For the morphology observations 6 individuals were used. Experimental arm amputation was done following to Dupont and Thorndyke (2006). The arm into the right of madreporite was used during the experiment. Autotomy was done about 10 mm from the arm tip after relaxation with MgCl2 5 % (Rubilar, 2010). After amputation, the individuals were returned to the aquarium. Experimental individuals were kept in the aquaria and were monitored at 2, 4, 6, 8, 11 and 16 days post amputation (dpa). At each time, one individual was randomly selected and 15 mm of the regenerating arm was amputated and used for morphology observations. For SEM (Scanning Electronic Microscope) observations on whole mount were made as follow: the tissue was fixed in paraformaldehyde (PF) 4 % in phosphate buffer solution (PBS) (0.01 M, pH = 7.4) + glutaraldehyde (GA) 2 % and then the tissue was dehydrated in ethanol series and dried to critical point using Hexamethyldisilazne (Sigma). Each arm was extended and mounted with a double-sided adhesive tape on a labelled slide. Arms were observed under a JEOL JSM-6460 LV microscope using the back-scattered electron (BSE) technique at low vacuum (15 - 20 Pa). Digital images were taken and specifications were recorded (magnification, plate inclination and rotation, scale, resolution, etc.).
Experimental design for histological observation
27 individuals were used for histological observations. Amputation procedure was performed as explained above. After amputation, the individuals were returned to the aquarium. Three individuals were used as controls (C1). Control individuals were not subject to amputation and were kept in the aquarium until completion of the experiment. Experimental individuals were kept in the aquarium and were monitored at 2, 4, 6, 8, 11, 16, 20 and 30 days post amputation (dpa). At each time, three individuals were randomly selected and 15 mm of the regenerating arm was amputated and used for histological observations by fixing the arm tip in PF 4% in PBS (0.01 M, pH = 7.4). After fixation the tissue was rinse with PBS (0.01 M, pH = 7.4) and then decalcified according to Mashanov, Zueva, Heinzeller and Dolmatov (2006) for a week. The tissue was dehydrated in ethanol series and embedded in paraffin. Sections of 7 µm were stained with Masson´s Trichrome. Slides were observed and analyzed using a Zeiss AxioImager Z1 microscope. Images were taken with an AxiocamHRc digital camera attached to the microscope.
Cell count and statistical analysis
Slides were observed and divided by group. Different cell types were counted, measured and identified by using the classification by Smith (1981), Byrne (1994), Candia-Carnevali, Bonasoro, Patruno and Thorndyke (1998) and Candia-Carnevali and Bonasoro (2001). Differential interference contrast was used to improve the cell identification at different days post amputation. The type and number of cells observed were counted in three fields in each section by using a 100x objective. The cell density (cell mm-2) for each group was calculated throughout the regenerative process. A two-way ANOVA analysis was used to test for differences between the type of cells during the regeneration process and between the different types of cells in each days post amputation. Normality was tested using Shapiro Wilk´s test and homogeneity of variances was tested using Bartlet´s test. The data was transformed with the square root (Sokal & Rohlf, 1979).
Results
Macroscopic observation of the regeneration process
At two days post amputation (dpa) a contraction of the tip of the stump was observed. The wound of the surface of the stump presented a concave shape and was partly covered by the external skeletal shields and a thin epithelium (Fig. 1A). The wound healing was in progress. Fibrous material and tissue debris formed a largely incomplete cicatricle layer. At 4 dpa the wound surface acquired a flattened shape. However, the tissue did not show a defined structure. There were areas with calcium carbonate deposits, characterized by their white coloration and hexagonal shape (Fig. 1B). After 6 dpa the surface of the wound was more regular and continuous. The skeleton was more developed, evidenced by a greater amount of calcium carbonate deposits (Fig. 1C). At 8 dpa the wound surface and the concave shape was lost. The skeleton and surrounding tissues were more organized and the complete re-epithelialization of the wound was observed. The stump surface was covered by a thin new epithelium (Fig. 1D). At 11 and 16 dps the shape of the tip of the arm changed. Without differentiation of any structure (Fig. 1E). This stage of the regenerative process was characterized by the formation of a large number of ossicles (Fig. 1F).
Types of the cell found during histological observations of the regeneration process
Different cells types were identified during the experiment: coelomocytes, sclerocytes, granule cells, presumptive blastemic cells, phagocytes, cells in mitosis, cells with cytoplasmic expansions, amoebocytes and cells in differentiation (Fig. 2). The coelomocytes (3.0 ± 0.5 μ) were observed in the coelomic channels, these cells were characterized by a central nucleus and a small cytoplasm (Fig. 2A). Sclerocytes (5.0 ± 1.0 μ) showed a rounded appearance, acidophilic cytoplasm and lot of granules in the cytoplasm (Fig. 2B). Granule cells (2.0 ± 0.5 μ) were characterized by their large chromatophilic bodies (Fig. 2C). Blastemic cells (= undifferentiated, 8.0 ± 1.0 μ) showed a globular appearance, circular nucleus and translucent pink cytoplasm (Fig. 2D). Phagocytes were distinguished by their large size (14.0 ± 4.0 μ) and the residual bodies of different sizes inside their cytoplasm (Fig. 2E). Cells in mitosis (13.0 ± 4.0 μ) were characterized by the presence of two nuclei located at both ends of a large cytoplasm (Fig. 2F). Cells with cytoplasmic expansions (5.0 ± 0.7 μ) were characterized by the expansion of their cytoplasm and an ovalshaped cytoplasm (Fig. 2G). The amebocytes (19.0 ± 3.0 μ) were characterized by their large size (Fig. 2H). Cells in differentiation were distinguished by their small size (1.0 ± 0.3 μ) and their acidophilic circular-shaped cytoplasm (Fig. 2I). The mean cell density did not vary during the wound healing. However, if the types of cells varied significantly during the experiment (ANOVA, F5,8 = 14.25, p < 0.01). The sclerocytes were abundant throughout the study period, even though the density decreased. The amebocytes were the lesser abundant cell type.
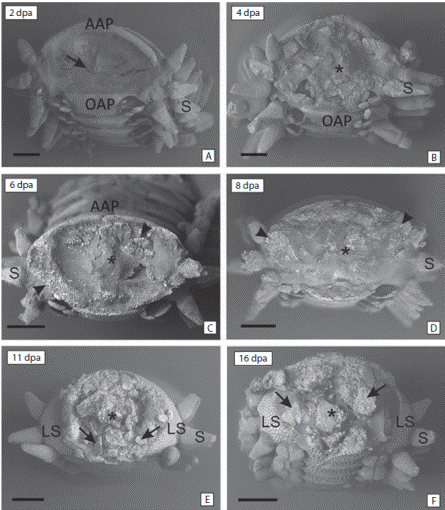
Fig. 1 Scanning Electronic Microscope images. Frontal view of the amputated arm tip at different times post amputation (dpa). (A) 2 dpa, denote the strongly concave shape (arrow) covered by the external skeletal shields and a thin epithelium. (B) 4 dpa, denote the flattened shape and the disorganized tissue (asterisk). (C) 6 dpa and (D) 8 dpa, denote the loss of the concave shape and the organized tissue (asterisk) and the calcium deposits (arrowhead). (E) 11 dpa and (F) 16 dpa, denote the ossicles in formation (arrows) and the growth of the arm (asterisk). AAP aboral arm plate, LS lateral shield, OAP oral arm plate, P podia, S spine. Scale bars: 500 µm.
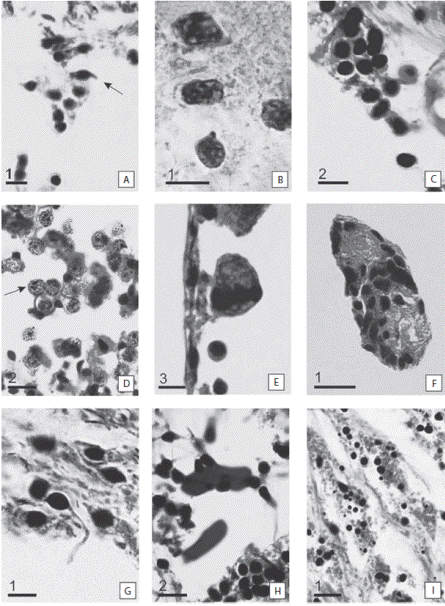
Fig. 2 Detail of cells involved in the regeneration process. (A) Coelomocytes (arrow). (B) Sclerocytes. (C) Granule cells. (D) Presumptive blastemic cells. Note the two nuclei of the cell (arrow). (E) Phagocytes. (F) Cells in mitosis. (G) Cells with cytoplasmic expansions. (H) Amoebocytes. (I) Cell in differentiation. Scale bars: 1: 5 μm; 2: 10 μm; 3: 20 μm.
Histological observation of the regeneration process
At 2 dpa (Fig. 3A, B, C) the wound was closed. The injured area acquired a concave shape. The amputation surface was covered by a complete and thin cicatricial layer (Fig. 3A). This tissue was characterized by a heterogeneous appearance, with no apparent structure. The tissue was discontinuous and had unclear boundaries (Fig. 3B). Beneath the cicatricial layer, the connective tissue was loose and without a clear tissue structure. During this period, the sclerocytes were the more abundant cells, followed by granule cells and coelomocytes. Phagocytes and amoebocytes were also observed (Fig. 3B, C).
At 4 dpa the wound surface showed a smooth concave shape (Fig. 3D, E, F). The amputation surface was covered by a welldeveloped cicatricial layer (Fig. 3E). The sclerocytes started to secrete collagen to form the endoskeleton. The ends of the coelomic cavity, epineural oral canal, the hyponeuralcanal and radial nerve cord were sealed by a large number of cells (Fig. 3F). The sclerocytes continued to have the highest density followed by the granule cells. Coelomocytes and phagocytes were observed. Cells with cytoplasmic expansions and amebocytes were also present in a low number.
At 6 dpa the amputation surface was no longer concave and acquired a more regular appearance at both ends and in the center of the arm (Fig. 3G, H, I). There were a large number of cells, primarily cells in differentiation, in the scar surface (Fig. 3H). The epithelium was continuous and thick (Fig. 3I). The coelomic cavity, the radial water canal and the sinuses surrounding the nerve remain sealed by primarily sclerocytes. The connective tissue appeared to have a denser compact appearance. The sclerocytes had the highest cell density, followed by phagocytes, cells in differentiation and granular cells.
At 8 dpa the wound was completely covered (Fig. 4A, B, C). Beneath the wound surface there was an intense cellular activity in the connective tissue (Fig. 4B). There were a large number of cells at the end of the nerve (Fig. 4C). At 16 dpa below the epidermis layer, the connective tissue presented loose fibers; there were a degree of rearrangement (Fig. 4D, E, F). The sclerocytes were close to each other; however, trabeculae were forming (Fig. 4E). The sclerocytes were the most abundant cells. However, cells in differentiation were important in this stage. Phagocytes, granular cells and coelomocytes were also present.
At 20 dpa the shape of the tip of the arm changed (Fig. 4G, H, I). A protrusion was observed on the amputation surface. The regenerative regrowth of the coelomic canals and the brachial nerve is outlined by a marked cellular flux towards the bud region. The outline of the presumptive blastema formation was observed as accumulation of a large number of cells (Fig. 4H). Trabeculae were already formed at both sides of the wound surface (Fig. 4I). At 30 dpa a presumptive blastema appeared as a small button, slightly emerging in the central-oral area of the autotomy surface was observed (Fig. 5A). The stalked presumptive blastema was formed mostly by presumptive blastemic cells. However, there were also cells in differentiation and sclerocytes (Fig. 5B, C, D).
Discussion
Amputation in Ophioplocus januarii triggers a number of processes. First, the wound is repaired. A thin cicatrizal tissue is formed to cover the wound surface to isolate the arm from the outside environment in order to prevent loss of coelomic fluid and infection. This tissue is very thin and formed mainly by sclerocytes and fibers. After this, the tissue starts to rearrange and the internal channels are sealed. The scleorocytes are the most important cells in this stage since they are responsible for secreting the trabeculae and skeleton growth (Byrne, 1994). Sclerocytes also function as nutrient storage cells (Byrne, 1994), which are needed to regenerate the lost tissue. Phagocytes are also important during this stage, as observed in Ophioderma longicauda, were they play an important role to prevent infections (Märkel & Röser, 1985). Coelomocytes and amoebocytes were also observed during this stage; these cells migrate to the site of the amputation and are important in the repair and regeneration processes (Candia-Carnevali et al., 1998). After the wound is closed (around 4 dpa) an epithelium can be observed over the wound surface. The skeleton starts to form, evidenced by new the trabeculae. A stalked presumptive-blastema indicating the beginning of the regeneration process appears at 30 dpa. This structure is closely associated to the radial nerve cord and it is formed mainly by presumptive blastemic cells. However, there are also two types of cells: sclerocytes and cells in differentiation. The scleorocytes will be responsible for the formation of the skeleton during regeneration of structures and the cells in differentiation are crucial to regeneration since, in the presumptive blastema-like structure the most important process is morphogenesis and differentiation of cells (Biressi et al., 2010).
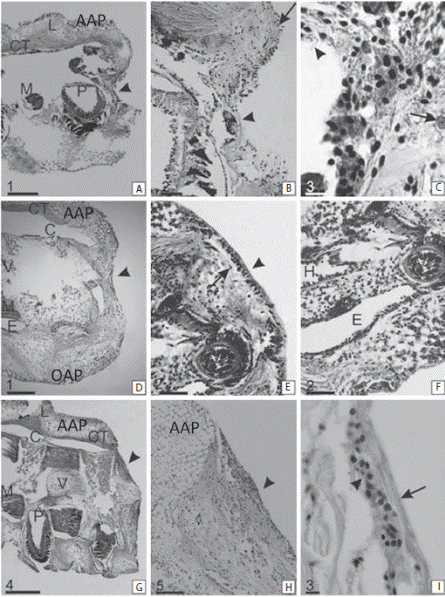
Fig. 3 Sagittal sections of the regenerating arm tip of Ophioplocus januarii. A-C 2 dpa. (A) Overview of the arm tip in regeneration. Denote the concave shape of the wound surface (arrowhead). (B) Detail of a. Denote the thin epithelium covering the wound. The central area is disorganized and discontinuous (arrowhead). The peripheral regions are more organized (arrow). (C) Detail of b. denotes the presence of various cell types; cells with cytoplasmic expansions (arrowhead) and phagocytes (arrow). D-F 4 dpa. (D) Overview of the arm tip in regeneration. Denote the smooth concave shape of the wound surface (arrowhead). (E) Detail of d. Denote the scar epithelium, is more organized and thicker (arrowhead). Under the epithelium there is a large amount of secreted fibers (arrow). (F) Detail of e. Denote the distal end of the channels sealed by sclerocytes. G-I 6 dpa. (G) Overview of the arm tip in regeneration. Denote the wound surface (arrowhead). (H) Detail of g, there are areas with high cellular density (arrowhead). (I) Detail of h. Denote the shape of the epithelium (arrow) and the more abundant cells (sclerocytes) (arrowhead). AAP aboral arms plate, C coelomic cavity, CT connective tissue, E epineural sinus, H hyponeural sinus, L interosciculars ligaments, M muscle, OAP oral arm plate, P podia, V vertebral ossicle. Scale bars: 1: 200 μm, 2: 50 μm, 3: 10 μm, 4: 500 μm; 5: 100 μm.
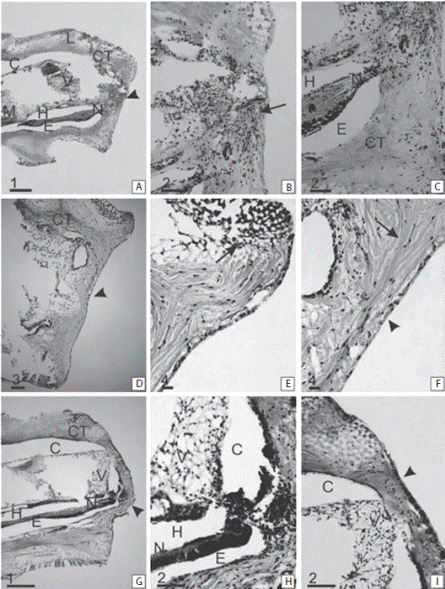
Fig. 4 Sagittal section of the regenerating arm tip of Ophioplocus januarii. A-C 8 dpa. (A) Overview of the arm tip in regeneration. Denote the straight shape of the wound surface (arrowhead). (B) Detail of c. Denote the injured surface with intense cellular activity, immersed in a dense disorganized connective tissue (arrow). (C) Detail of b. Denote the large cell accumulation in close association with the nerve. D-F 16 dpa. (D) Overview of the arm tip in regeneration. Denote the flattened shape of the wound surface (arrowhead). (E) Detail of d. Denote the formation of new sclerocytes at the aboral extremity of the wound (arrow). (F) Detail of e. Denote the organized epithelium (arrowhead) and the loose connective tissue fibers below it (arrow). G-I 20 dpa. (G) Overview of the arm tip in regeneration. Denote a protrusion in the oral surface (arrowhead). (H) Detail of g. Denote an accumulation of cells proximal to the nerve. (I) Detail of h. Denote the organized area formed by sclerocytes (arrowhead). AAP aboral arm plate, C coelomic cavity, CT connective tissue, E epineural sinus, G ganglion, H hyponeural sinus, L interosciculars ligaments, M muscle, N nerve, OAP oral arm plate, P podia, V vertebral ossicle. Scale bars: 1: 200 μm; 2: 50 μm; 3: 100 μm; 4: 20 μm.
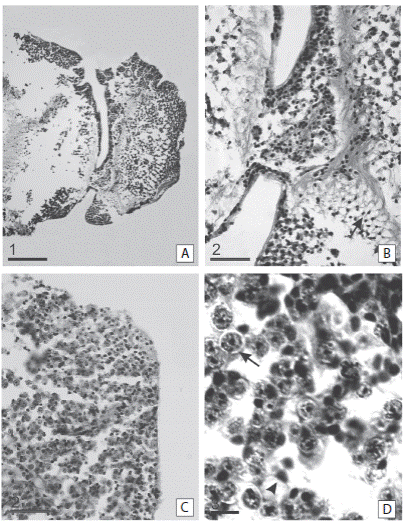
Fig. 5 Sagittal sections of the regenerating arm tip of Ophioplocus januarii. A-D 30 dpa. (A) Overview of the arm tip in regeneration. Denote the stalked presumptive blastema at the end of the arm, characterized by the presence of undifferentiated cells. (B) Detail of the stalked presumptive blastema. Denote the formation of trabeculae in the blastema area (arrowhead). (C) Detail of a, denote the abundance of presumptive blastemal cells (arrow) but also there are fibers of the connective tissue (arrowhead). (D) Detail of the stalked presumptive blastema. Denote presumptive blastemic cells (arrow) and sclerocytes (arrowhead). Scale bars: 1: 200 μm; 2: 50 μm; 3: 10 μm.
The presence of a blastema-like structure indicates that O. januarii regenerates through an epimorphic process as was found in other echinoderm species (Dupont & Thorndyke, 2006; Clark et al., 2007; Biressi et al., 2010). However, prior to the formation of the presumptive blastema there is a rearrangement of preexistent tissue and cells, indicating a morphollactic process. The co-existing of these two regenerative processes has been described previously in ophiuroids (Biressi et al., 2010). However, it also has been described in other classes of echinoderms (García Arrarás et al., 1998; Mashanov & Dolmatov, 2000; Candelaria, Murray, File & García Arrarás, 2006) and in vertebrates regeneration (Sánchez Alvarado & Tsonis, 2006). The blastema-like structure observed in echinoderms is equivalent to the blastema described in other phyla (Sánchez Alvarado & Tsonis, 2006; Yokoyama, 2008). However, it differs from the histological point of view; the presumptive blastema of echinoderms is not composed only of blastemic-like cells, there are also sclerocytes (Candia-Carnevali & Bonasoro, 2001; Biressi et al., 2010; this work). In ophiuroids, regeneration of the arm requires the formation of new ossicles to form shields, vertebrae, spines and other components of the skeleton. In fact, the skeleton is the most abundant tissue of the arm. Immediately after amputation the formation of new ossicles, even without the presence of the presumptive blastema, can be observed.
The identity of the cells forming the presumptive blastema in echinoderms is still unknown. It was postulated that in crinoids and ophiuroids the blastema-like structure is formed by coelomocytes that migrate from the coelomic channel. These cells dedifferentiate and generate new ones (Candia-Carnevali & Bonasoro 2001; Biressi et al. 2010). Although we were not able to visualized migrating cells toward the tip of the amputated arm, we cannot rule out this possibility.
Biressi et al. (2010) described the regeneration process of two other brittle stars (Amphiura filiformis and Ophioderma longicauda). The processes described of wound healing are quite similar to the one observed in this paper for O. januarii. Although, the regeneration rates in A. filiformis and O. longicauda are quite different (Dupont & Thorndyke, 2006), the presumptive blastema structure appears at similar times in these species and is coincident with O. januarii. Even though these three species have different feeding habits, habitat, depth range and size (Dupont & Thorndyke, 2006; Brogger, 2010), the wound healing process is quite similar. Arm tip regeneration in ophiuroids has been well documented in the fossil records (Aronson, 1987; 1991) and in living species (Pomory & Lawrence, 1999; 2001; Dupont & Thorndyke, 2006; Rodríguez, 2009). Since arms are crucial for feeding, locomotion, reproduction, social interaction (Lawrence, 1987), the process of regeneration it is probably a common feature among species. Future studies with focus on different species of Ophiuroidea may support this idea.

