











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkLos arrecifes de coral constituyen uno de los ecosistemas marinos más complejos, con una alta riqueza y diversidad de especies y una comunidad íctica arrecifal particularmente conspicua (Rocha et al., 2005). Debido a su gran biodiversidad, están considerados dentro de los ecosistemas más importantes en el planeta, equiparándose con los bosques tropicales (Sale, 1977), lo cual es consecuencia de la amplia variedad de hábitats que crean diferentes espacios, los cuales pueden ser colonizados por numerosas especies; siendo los peces uno de los grupos o taxa más representativos (Rocha et al. 2005).
Los ensamblajes de peces arrecifales generalmente se encuentran ligados fuertemente a un sustrato que les brinde protección y alimento (Sale, 1991); sin embargo, cada especie muestra preferencias por hábitats muy precisos, (Allen & Robertson, 1994). De este modo, los peces arrecifales explotan dichas áreas coralinas mediante interacciones directas (i.e. depredación de los pólipos de coral), e indirectas(i.e. el forrajeo), con efectos inmediatos sobre el crecimiento y distribución de corales así como de la fauna asociada a escala local, (Sale, 1991). Debido a esto, la estructura de una comunidad de peces es resultado de la variabilidad regional de las especies presentes y de los procesos que influyen en la estabilidad y persistencia de los ensamblajes locales de esas especies (Sale, 1980); tales como competencia, depredación, reclutamiento y disturbios abióticos, los cuales determinan la predominancia de especies herbívoras, carnívoras, etc. (Ebeling & Hixon, 1991).
Otros habitantes importantes de los sistemas arrecifales y que coadyuvan a la estabilidad de los mismos, son los erizos del género Diadema, considerados “especie clave” por su función ecológica (Edmunds & Carpenter, 2001). Diversos estudios en campo y manipulaciones experimentales demuestran que las especies del género Diadema tiene efectos positivos como la disminución de la biomasa algal, aumento de la cobertura coralina, y aumento de la diversidad de especies, con influencia en la dinámica y estructuración de las comunidades costeras (Sammarco et al., 1974; Ogden & Lobel, 1978; Lessios et al., 2001; Tuya et al., 2004b). El efecto negativo de éstas especies se observa cuando existen altas densidades poblacionales, debido a que estos organismos pueden remover más carbonato de calcio de los arrecifes que cualquier otro habitante (Eakin, 2001; Herrera-Escalante et al., 2005). De esta manera, poblaciones excesivamente grandes de este ramoneador, pueden afectar de forma directa la abundancia de peces con los que compiten por alimento (McClanahan et al., 1996). Por ejemplo, en los arrecifes del Caribe, se ha observado una correlación inversa entre la densidad de Diadema antillarum y la densidad de peces herbívoros (Randall, 1961), sin embargo ambos mantienen una fuerte correlación en el ramoneo de las macroalgas (Tuya et al., 2004a), por lo tanto, los peces herbívoros y erizos tienen un efecto similar sobre las macroalgas (Sandin & McNamara, 2011). Además, se ha observado que los erizos del género Diadema, compiten directamente con especies de peces del género Stegastes y otros peces herbívoros (Williams, 1977; Sammarco & Williams, 1982, Eakin, 1988).
Por otra parte, desde hace algunas décadas se han reportado eventos de mortandades masivas de erizos, principalmente para zonas con grandes extensiones de arrecife como el Gran Caribe y la Gran Barrera Arrecifal Australiana. En el Gran Caribe, el evento se registró por primera vez en Panamá a principios de los años ochenta (Lessios et al., 1983; Lessios et al., 1984a, 1984b) e inmediatamente se realizaron numerosos estudios para evaluar los efectos en vertebrados e invertebrados (Bak et al., 1984; Liddell & Ohlhorst, 1986; Hughes et al., 1987; Lessios, 1988; Carpenter, 1988, 1990a, 1990b). En el Pacífico Mexicano, Benítez-Villalobos et al. (2009) reportaron por primera vez un evento de mortandad masiva del erizo negro Diadema mexicanum en La Entrega, Huatulco, México; los autores mencionaron una disminución severa en la densidad poblacional del erizo (de ~ 4.42 ind.m-2 a 0 ind.m-2) resultando en la desaparición de la población local de La Entrega y describieron características similares a las observadas en el Caribe para D. antillarum (mucosidad en las espinas, desprendimiento de espinas, perforación de las testas, lesiones en el peristoma).
La estructura poblacional de D. mexicanum depende de las interacciones competitivas con otros herbívoros (principalmente peces y algunos invertebrados) por recursos (alimento, sustrato, espacio), por lo que un cambio abrupto en la población del erizo provocado por un evento de mortandad masiva, modificaría la estructura de la comunidad íctica arrecifal, afectando principalmente a los gremios de los peces herbívoros y omnívoros, y en menor grado a otros niveles tróficos. En este trabajo se describen los efectos causados por la ausencia de D. mexicanum sobre la fauna íctica arrecifal de La Entrega a causa del evento de mortandad masiva de 2009
Materiales y métodos
Área de estudio
El área de estudio se ubica en las Bahías de Huatulco, las cuales se encuentran en la porción central de la costa de Oaxaca, entre la desembocadura de los ríos Copalita (15°45’48” N y 96°04’56” W) y Coyula (15°41’09” N - 96°14’45” W). Esta área es relevante por ser una de las zonas con mayor riqueza coralina (17 especies) en el Pacífico Mexicano (Glynn & Leyte-Morales, 1997; López-Pérez & Reyes-Bonilla, 2000).
En la zona se presentan dos temporadas climáticas bien marcadas correspondientes a la época de secas (noviembre-abril) y la época de lluvias (mayo-octubre) con una precipitación media de 817.7 mm. De noviembre a febrero se presentan vientos denominados localmente “nortes” o “tehuanos” provenientes del Golfo de México (Monreal-Gómez & Salas de León, 1998) que se originan por los frentes fríos atmosféricos que pasan por el Golfo de México. Esta actividad provoca que el agua superficial del Golfo de Tehuantepec se vuelva más fría por la recirculación del agua profunda, lo cual contribuye a la productividad biológica de la región (Lavín et al., 1992). La temperatura superficial del mar fluctúa entre los 26 y 28°C, la salinidad oscila entre 33.5 y 34.5 ppm y la profundidad de la termoclina es de 60 m durante la mayor parte del año (Fiedler, 1992).
La Entrega (15°44’34’’ N - 96°07’35’’ W) se caracteriza por ser una bahía semi-cerrada por puntas rocosas, las cuales protegen al arrecife coralino del oleaje directo. Dentro de la bahía se puede identificar la zona coralina con una extensión aproximada de 7 ha, conformada por siete especies de corales: Pocillopora verrucosa, P. capitata, P. damicornis, P. effusus., Pavona gigantea, P. varians y Porites panamensis, de las cuales P. damicornis es la más abundante (Glynn & Leyte-Morales, 1997; López-Pérez & Hernández-Ballesteros, 2004). El segundo componente de mayor importancia es el coral muerto (López-Pérez & Hernández-Ballesteros, 2004) y la máxima profundidad del arrecife es de 13.3 m (Glynn & Leyte-Morales, 1997). Durante la temporada de lluvias la comunidad arrecifal presenta una tasa de sedimentación alta (14.33 kg m-2 año-1), sin embargo, en comparación con otras comunidades arrecifales, se considera baja (GranjaFernández & López-Pérez, 2008), levemente por encima de la tasa ideal (10 kg m-2 año-1) propuesta por Rogers (1990).
Trabajo de campo
Para estimar la composición de especies, se utilizó el método de conteo puntual aleatorio de Bohnsack y Bannerot (1986), modificado por VillarrealCavazos et al. (2000). Este método es práctico, sencillo, puede ser repetitivo sin causar daño o destrucción en el arrecife y se ha utilizado anteriormente en la zona de estudio obteniéndose resultados satisfactorios (Ramírez-Gutiérrez et al., 2007; López-Pérez et al., 2010, 2013). El método consiste en contar todos los peces que ingresan a un cilindro imaginario que se extiende desde el fondo hasta la superficie, en un radio (φ) de 5 m durante un periodo de 5 min. El cilindro estuvo delimitado por una cadena de plástico de 5 m de largo (φ); en un extremo se colocó un plomo considerado como el centro del cilindro y posteriormente se fue rotando en una sola dirección y se anotaron todas las especies transitorias, altamente móviles y no territoriales observadas dentro del cilindro. Posteriormente, un tiempo y recorrido similar fue usado para contabilizar especies crípticas. Se realizaron muestreos mensuales de agosto 2009 a julio 2010 en los que se realizaron censos diurnos entre las 10:00 y las 15:00 h. En cada muestreo se obtuvieron diez cilindros, cinco en la zona somera (2 - 5 m) y cinco en la zona profunda (6 - 10 m), cubriendo un área de observación de 785 m2 por mes.
Trabajo de gabinete
Para poder identificar y analizar los cambio, se obtuvieron datos de la comunidad íctica arrecifal anteriores al evento de mortandad, los cuales provienen de muestreos que fueron realizados de febrero de 2006 a enero de 2007. La metodología que se empleó en el presente trabajo para los censos realizados posteriormente al evento de mortandad, es la misma que se utilizó en los muestreos de 2006-2007.
La identificación de las especies se realizó mediante guías ilustradas y base de datos online de especies de peces arrecifales del Pacífico Oriental Tropical (POT) como Allen y Robertson (1994), Robertson y Allen (2006) y Fishbase (2010). La validez de los nombres se corroboró en WORMS (World Register of Marine Species, http://www.marinespecies.org/ index.php), ITIS (Integrated Taxonomic Information System, http://www.itis.gov/), Robertson y Allen (2008, http://biogeodb.stri.si.edu/ sftep/taxon.php) y Fishbase (www.fishbase. org). El ordenamiento sistemático se realizó de acuerdo con Eschmeyer et al. (2014). El arreglo de los géneros y sus respectivas especies se presentan en orden alfabético.
Debido a que el nivel trófico muestra la posición en la que se encuentra ubicado un componente biótico dentro de la red trófica en un ecosistema y su clasificación depende de la corriente de energía y nutrientes que se establece entre las distintas especies, se organizó a las especies en los siguientes grupos alimenticios:Carnívoros (C), Herbívoros (H), Detritívoros (D), Planctívoros (P), Omnívoros (O) y Parasíticos (P) considerando sus hábitos alimenticios de acuerdo con Robertson y Allen (2006).
Descriptores ecológicos de la comunidad
La riqueza específica se consideró como el número total de especies de peces obtenido en los censos visuales en la comunidad. Para calcular la diversidad de especies de cada localidad se utilizó el Índice de Shannon. La equidad de Pielou se estimó para poder interpretar correctamente el índice de diversidad de Shannon. Para evaluar la dominancia, se utilizó el Índice de Simpson (Magurran, 1988).
Los descriptores ecológicos (riqueza, diversidad, equidad y dominancia) se calcularon mensualmente a partir de una matriz ensamblada de datos de abundancia de especies por unidad de esfuerzo (cilindro), usando el programa PRIMER 6 (V. 6.1.10). Posteriormente se aplicó un análisis de la varianza (Andeva-1 vía) para cada estimador ecológico, para detectar diferencias estadísticamente significativas entre estudios; así como una prueba de comparación múltiple a posteriori de Tukey (HSD) para determinar las diferencias obtenidas. En todos los casos se usó un nivel de significancia de α = 0.05.
Diferencias entre periodos
Se utilizó la técnica de ordenación espacial mediante escalamiento multidimensional no-métrico (MDS) para detectar gráficamente la existencia de algún patrón de agrupamiento o separación (Clarke & Warwick, 2001). El análisis se realizó utilizando una matriz de similitud basada en el índice de Bray-Curtis.
Para determinar si las diferencias eran estadísticamente significativas se realizó un análisis de varianza no paramétrico basado en permutaciones (PERMANOVA) de una vía, considerando el periodo (antes y después) como único factor. El PERMANOVA fue aplicado a una matriz de similitud basada en el índice de Bray-Curtis, con los datos de abundancia transformados mediante raíz cuarta y realizando 9 999 permutaciones. De manera complementaria, se realizó un análisis de importancia relativa de especies (SIMPER) con la finalidad de identificar a las especies que contribuyeron a la formación de grupos y determinaron la diferencia entre los periodos. Para este análisis también se utilizó una matriz de similitud de Bray-Curtis.
Resultados
Riqueza de especies y abundancia
En el estudio anterior al evento de mortandad se registraron 6 776 individuos pertenecientes a 61 especies. Las familias mejor representadas fueron Pomacentridae (8 especies), Labridae (7 especies), Serranidae (5 especies) y Haemulidae (4 especies). Las especies más abundantes durante todo el estudio fueron Thalassoma lucasanum (19 %), Haemulon maculicauda (18 %), Selar crumenophthalmus (15 %), Chromis atrilobata (14 %) y Stegastes acapulcoensis (13%). Posterior al evento se registraron 32 920 individuos pertenecientes a 53 especies. Las familias mejor representadas fueron Serranidae (6 especies), Labridae (5 especies), Pomacentriade (4 especies), Haemulidae, Lutjanidae y Muraenidae (3 especies cada una). Las especies más abundantes durante todo el estudio fueron H. maculicauda (29%), T. lucasanum (25 %), S. crumenophthalmus (14 %), S. acapulcoensis (13 %) y Prionurus punctatus (7 %). La riqueza de especies fue mayor antes del evento, sin embargo, la abundancia proporcional registrada fue significativamente mayor en el muestreo posterior al evento (Fig. 1).
En relación con la estructura trófica, antes del evento de mortandad, el arreglo trófico estuvo compuesto por un 46.66 % de especies carnívoras, 21.33 % de especies omnívoras, 21.33 % de especies planctívoras, 9.33 % de especies herbívoras y un 1.33 % de especies parasíticas. En el periodo posterior al evento, el arreglo trófico estuvo conformado por un 43.28 % de especies carnívoras, 14.93 % de especies omnívoras, 29.85 % planctívoras, 10.45 % de especies herbívoras y 1.49 % de especies de hábitos parasíticos (Fig. 2).
Analizando específicamente el gremio de peces herbívoros, se observaron diferencias estadísticamente significativas entre ambos estudios (Andeva F = 39.091, gl = 1, p < 0.05). Las familias más representativas en términos de especie/abundancia fueron Kyphosidae (2 especies), Scaridae (2 especies), Pomacentridae (3 especies) y Acanthuridae (3 especies). Los cambios más significativos se observaron en la familia Pomacentridae (F = 163.37, gl = 1, p = 0.000), siendo Stegastes acapulcoensis la especie herbívora que presentó un aumento significativo en la abundancia en el periodo posterior al evento de mortandad (Fig. 3).
Descriptores ecológicos comunitarios
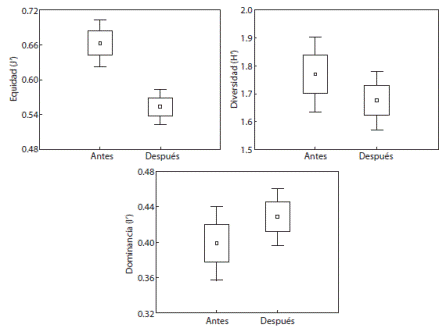
El valor promedio de los estimadores ecológicos durante todo el estudio previo al evento fue (media ± desv.est.): Diversidad de Shanon, H’ = 1.688 ± 0.289 bits / individuo; equidad de de Pielou J’ = 0.653 ± 0.055 y dominancia de Simpson I’ = 0.414 ± 0.08. En el periodo posterior al evento de mortandad, el valor promedio de los estimadores ecológicos fue: H’ = 1.675 ± 0.206 bits / individuo; J’ = 0.553 ± 0.044 e I’ = 0.429 ± 0.046. El Andeva mostró que no existieron diferencias estadísticamente significativas tanto para la diversidad (p = 0.277) como la dominancia (p = 0.258) entre ambos periodos; sin embargo, la equidad si mostró diferencias significativas (p < 0.01), indicando que fue mayor antes del fenómeno (Fig. 4).
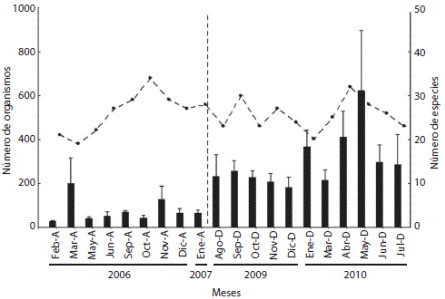
Fig. 1 Monthly average abundance (± S. E.) and number of species (line with dots) of fishes corresponding to the periods before and after the D. mexicanum mortality event. The vertical dashed line indicates the separation between periods.
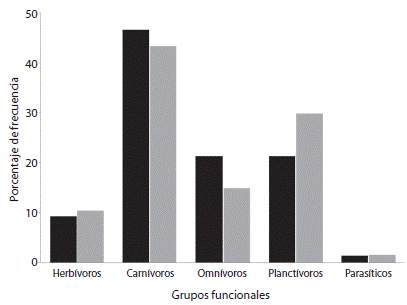
Fig. 2 Species percentage of each trophic level at La Entrega, on the periods before (black bars) and after (gray bars) the D. mexicanum mortality event.
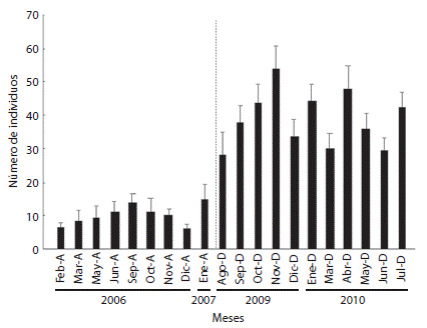
Fig. 3 Monthly average abundances (± S. E.) of Stegastes acapulcoensis corresponding to periods before and after the D. mexicanum mortality event. The vertical dashed line indicates the separation between periods.
Diferencias entre periodos
El análisis MDS con un nivel de stress de 0.18, mostró una separación clara entre los meses previos y posteriores al evento de mortandad de D. mexicanum (Fig. 5). El análisis PERMANOVA mostró diferencias estadísticamente significativas en el ensamblaje de peces en La Entrega con respecto a los periodos de antes y después del evento de mortandad (Cuadro 1), coincidiendo con la representación gráfica del MDS. El análisis SIMPER determinó que la similitud promedio antes del evento fue menor (42.62 %) que después (56.99 %). Las especies que contribuyeron a la formación del grupo antes del evento de mortandad fueron T. lucasanum (P) aportando el 37.52 %, S. acapulcoensis (H) con el 30.59 %, H. maculicauda (H) con el 7.97 % y S. flavilatus con el 5.08 %. Las especies que contribuyeron a la formación del grupo posterior al evento fueron H. maculicauda (36.03 %), T. lucasanum (32.38%), S. acapulcoensis (21.05 %) y P. punctatus (3.06 %). La disimilitud entre ambos estudios se debió principalmente a H. maculicauda (28.9%), T. lucasanum (22.2 %), S. crumenophthalmus (12.9 %), S. acapulcoensis (11.4 %) y P. punctatus (6.8 %).
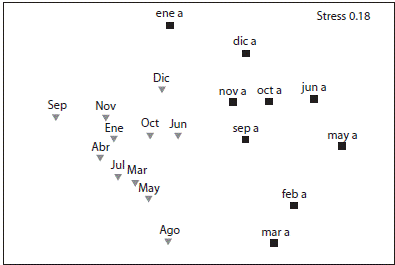
Fig. 5 Nonmetric Multidimensional Scaling (obtained with Bray-Curts similarity), of reef fishes at La Entrega. The months before (black squares) and after (gray triangles) the D. mexicanum mortality event are shown.
Cuadro 1 Análisis de varianza de una vía no paramétrico basado en permutaciones de los datos de abundancia. Los valores de P se obtuvieron usando 9 999 permutaciones
| Source | df | SS | MS | Pseudo-F | P(perm) | Unique perms | P(MC) |
|---|---|---|---|---|---|---|---|
| Ti | 1 | 3956.3 | 3956.3 | 5.7935 | 0.0001 | 9612 | 0.0001 |
| Res | 18 | 12292 | 682.89 | - | - | - | - |
| Total | 19 | 16248 | - | - | - | - | - |
Discusión
Tanto los datos de riqueza de especies como los indicadores ecológicos comunitarios obtenidos en este trabajo mostraron diferencias importantes en las estructura de la comunidad de peces antes y después del evento de mortandad de D. mexicanum en La Entrega. De manera general, los descriptores ecológicos mostraron en el periodo anterior al evento de mortandad altos valores de equidad y bajos valores de dominancia, lo que implica que estructuralmente se registraron pocas especies dominantes. Lo anterior es considerado como una característica típica de una comunidad estable (McCloskey, 1970); mientras que después del evento se observaron bajos valores de equidad y altos valores de dominancia, demostrando una inestabilidad aparente.
Los valores de diversidad obtenidos después de la mortandad del erizo, fueron inferiores a los reportados para La Entrega por Barrientos-Villalobos (2000) (51 especies en 26 familias) y Barranco-Servín (2003) (63 especies en 29 familias) quienes registraron valores del índice de Shannon entre 2.095 y 3.577 bits/individuo. A este respecto, Ravera (2001) menciona que valores bajos de diversidad son indicativos de que un ecosistema se encuentra sometido a situaciones de tensión y estrés, mientras que los valores altos se relacionan con comunidades estables (McCloskey, 1970).
Las variaciones observadas en la riqueza y abundancia de las especies podría deberse a variaciones temporales propias de la comunidad, asociadas a las fluctuaciones en las características oceanográficas de la zona, sin embargo en el trabajo de López-Pérez et al. (2013) realizado entre febrero de 2006 y septiembre de 2009 en la misma área, se indica que a pesar de las variaciones espacio-temporales de la comunidad íctica en los arrecifes de las bahías de Huatulco, como patrón general se observa que las comunidades con mayores abundancias y menor diversidad son aquellas relacionadas a localidades donde el componente mayor del sustrato son las rocas, mientras que una diversidad más alta se asocia a localidades con una mayor proporción de coral muerto y fragmentos de coral. A ese respecto, la comunidad íctica de La Entrega antes del evento de mortandad cumplía con ese patrón, ya que el componente principal del arrecife es el coral vivo y el segundo componente es el coral muerto y la comunidad de peces mostraba diversidad alta y abundancias moderadas, comparada con otras localidades, pero en el periodo posterior al evento de mortandad de D. mexicanum se observó un patrón similar a que si fuera una comunidad donde predominan las rocas y no el coral.
Los valores de equidad y dominancia, en los primeros meses posteriores a la mortandad de D. mexicanum mostraron valores cercanos y posteriormente comenzaron a mostrar diferencias más marcadas, lo cual sugiere que la abundancia de algunas de las especies registradas después del evento se incrementó de manera importante, aumentando de esta manera los valores de dominancia, sin embargo en este periodo inicial aún no se definían las especies dominantes de la comunidad y esto se reflejó en una equidad media que posteriormente disminuyó a medida que la dominancia se incrementaba.
Analizando los resultados obtenidos por el SIMPER, se observó que el primer grupo formado antes de la mortandad, y de acuerdo al porcentaje de contribución de cada especie, estuvo conformado por una especie omnívora (Thalassoma lucassanum), una herbívora (Stegastes acapulcoensis) y una carnívora (Haemulon maculicauda). Éste arreglo es típico de una comunidad íctica arrecifal tropical, en la que la diferencia en las abundancias relativas es muy característica debido a que los omnívoros son particularmente abundantes y conspicuos (Emery, 1968; Davis & Birdsong, 1973). El segundo grupo formado con especies registradas después del evento, presentó una conformación distinta, con un evidente incremento en las abundancias de los herbívoros y una disminución en las abundancias de los omnívoros. Nuevamente, lo anterior confirma una inestabilidad en la comunidad íctica arrecifal después del evento, sugiriendo que los carnívoros (H. maculicauda) y herbívoros (S. acapulcoensis) fueron más importantes en cuanto a la contribución en términos de su abundancia.
A partir de los análisis realizados para el gremio de los herbívoros, se obtuvo que Stegastes acapulcoensis fue la especie más afectada por la ausencia de Diadema mexicanum, ya que se registró un aumento significativo en su densidad poblacional en los meses posteriores al evento. Una posible explicación del aumento en la densidad de esta especie después de la mortandad, se fundamenta en los hábitos biológicos-ecológicos de la especie, ya que son peces que “cultivan” jardines de macroalgas, podándolas y protegiéndolas constantemente, para usarlas como alimento y/o zonas de anidación (Wellington, 1982; Wellington & Víctor, 1988). En ese sentido, un posible aumento en las macroalgas (datos sin publicar), debido a la ausencia de D. mexicanum significó un incremento importante en la disponibilidad de este recurso para S. acapulcoensis, al mismo tiempo que también disminuyó la competencia directa representada por el erizo. De esta manera, los organismos adyacentes a la bahía (provenientes de las zonas rocosas) pudieron haber emigrado hacia la plataforma arrecifal, ocupando aquellos espacios vacíos de D. mexicanum. La fisiografía de La Entrega, pudo haber facilitado la migración de una zona a otra, debido a que las distancias entre ambos puntos (arrecifal y rocoso) es relativamente insignificante.
Es importante mencionar que, interacciones competitivas entre esta especie y Diadema mexicanum han sido documentadas en Isla Uva, Panamá; donde se observó que S. acapulcoensis desempeña una importante función en los ecosistemas arrecifales, debido a que al excluir agresivamente al erizo y evitar su presencia en sus céspedes algales, protege alrededor del 10% del total de la zona arrecifal de los erizos, evitando la erosión causada por D. mexicanum (Eakin, 1988). Por lo tanto, de acuerdo con los resultados obtenidos, se propone que el evento de mortandad masiva de D. mexicanum benefició a S. acapulcoensis debido a características ecológicas, y hábitos alimenticios de esta especie. Por otra parte, es posible que la ausencia de D. mexicanum influyera en otros grupos de invertebrados epifíticos, quienes normalmente se alimentan por medio del ramoneo o utilizan las algas como protección. La abundancia de invertebrados como poliquetos, crustáceos y moluscos entre otros, pudo haberse incrementado debido a una alta disponibilidad de recurso (macroalgas) y a la disminución competitiva de D. mexicanum tanto por alimento como por espacio para asentamiento. Este efecto de liberación ecológica podría explicar el aumento en la abundancia de peces de hábitos carnívoros, los cuales al desaparecer la actividad competitiva de D. mexicanum, utilizaron el recurso disponible generando un aumento en su densidad.
Las especies herbívoras secundariamente dominantes después del evento fueron Prionurus punctatus (Acanthuridae) y Scarus ghobban (Scaridae), los cuales también influyen en otras especies, ya que se ha observado en campo que generalmente Arothron meleagris, Microspathodon bairdii y Pomacanthus zonipectus se asocian a los cardúmenes de P. punctatus, alimentándose en las zonas donde el sustrato ha sido removido por esta especie.
Actualmente existe un importante debate acerca de las consecuencias de los disturbios físicos y biológicos sobre la comunidad íctica arrecifal, pero aún se conoce muy poco sobre los efectos que estos puedan ocasionar. De hecho, estos eventos modifican la estructura del hábitat, alterando a la comunidad íctica en función del tipo de disturbio, así como la magnitud e intensidad del mismo (Jones & Syms, 1998). En este estudio se observó que la mortandad de la especie clave D. mexicanum, tuvo efectos inmediatos en la comunidad íctica de hábitos alimenticios similares al erizo, afectando de manera directa a Stegastes acapulcoensis y en menor grado a otros herbívoros como Scarus gobban y Prionurus punctatus; mientras que de manera indirecta, otros peces de hábitos carnívoros aumentaron sustancialmente, lo que sugiere que el impacto fue más profundo, alterando a toda la fauna arrecifal.

