











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkLa costa del Pacífico costarricense corresponde con una porción importante del Pacífico Tropical Oriental (PTO), uno de los ecosistemas de aguas abiertas más productivos y con mayor biodiversidad en megafauna (Ballance, Pittman & Fiedler, 2006). May-Collado, Gerrodette, Calambokidis, Rasmussen y Sereg (2005) hacen una descripción detallada del patrón de avistamiento general para cetáceos en aguas territoriales del Pacífico, en específico, en términos de riqueza de especie y distribución potencial. Sin embargo, dicha contribución se sustenta en su mayoría, en datos de censos focalizados al ambiente oceánico a escala regional del PTO. En consecuencia, el patrón de diversidad en cetofauna, presenta vacíos a nivel de meso-escala y de patrones locales.
Golfo Dulce es una de las áreas de estudio en Costa Rica con mayor productividad académica en trabajos pertinentes a ecología de cetáceos (Acevedo & Smultea, 1995; Acevedo-Gutiérrez, 1996; Acevedo-Gutiérrez & Burkhart, 1998; Cubero-Pardo, 1998; AcevedoGutiérrez & Matthews, 2005; Cubero-Pardo, 2007a; Cubero-Pardo, 2007b; Oviedo, 2007; Pacheco-Polanco & Oviedo, 2007; Oviedo, 2008; Oviedo, Pacheco & Herra-Miranda, 2009; Pacheco-Polanco, Oviedo, Herra-Miranda & Silva, 2011; Márquez-Artavia et al., 2012; Oviedo et al., 2012; Bessesen et al., 2014), esto debido a los atributos fisiográficos y estructurales de este importante hábitat marino-costero, que lo hacen un excelente laboratorio natural para el estudio de cetáceos. Los retos metodológicos y logísticos del trabajo con mamíferos marinos, en áreas donde los limites son más fluidos que fijos, son facilitados notablemente en Golfo Dulce, lo que incide en la efectividad de la colecta de observaciones con mayor resolución, así como en la potencialidad para estudios a largo plazo.
En este estudio presentamos un análisis descriptivo espacial de la diversidad de cetáceos en Golfo Dulce, a nivel de estructura de comunidades. Dentro de dicha estructura comunitaria se detalla la riqueza de especies incluyendo la posible presencia de ecotipos en las formas específicas, ya que las razas ecológicas se asocian a la estructura de hábitats. El objetivo es proveer una visión integrada de la diversidad estructural del hábitat y la composición faunística de nuestro grupo indicador Cetacea, a lo largo de nueve años de muestreo con presencia sistemática en campo. Este estudio sigue las premisas establecidas por Pielou (1975) para el estudio de comunidades ecológicas, que enfatizan la importancia de definir y especificar claramente: a) los limites espaciales del área en evaluación y los métodos de muestreos, b) el tiempo límite de las observaciones y finalmente, c) el set de especies perteneciente a un mismo taxón.
A pesar del carácter único de Golfo Dulce como un hábitat marino costero: un estuario tropical con características similares a fiordos y una biodiversidad marina particularmente rica, aun este no posee una categoría de manejo que garantice no solo la protección integral de sus recursos, sino la utilización sustentable de los mismos. Este trabajo al describir y documentar espacialmente la distribución de las especies que constituyen las comunidades de cetáceos en Golfo Dulce, representa entonces, un aporte a la comprensión en la diversidad de cetáceos en aguas del Pacífico Sur de Costa Rica, información que se integraría a la base de conocimiento científico sobre el ecosistema marino y su dinámica, que debe regir estrategias de gestión y manejo, en particular aquellas dirigidas hacia un Manejo de Base Ecosistémica (Crowder & Norse, 2008).
Materiales y métodos
Área de Estudio
Golfo Dulce tiene una extensión aproximada de 50km de largo y 10 a 15km de ancho, situado en la región del Pacífico Sur de Costa Rica, este estuario tropical tiene una fisiografía y patrón de circulación similar a un fiordo (Wolff, Hartman & Koch, 1996; Quirós, 2003; Oviedo, 2007). El clima predominante en la Península de Osa es tropical húmedo, con una estación lluviosa (finales de mayo, principios de noviembre) que genera una precipitación promedio mensual dentro del rango 100-700mm (Quesada-Alpízar & Morales-Ramírez, 2004). Los ríos Coto Colorado, Tigre, Esquinas y Rincón tienen influencia directa en el patrón de circulación y las corrientes superficiales. Golfo Dulce se sub-divide en tres unidades de paisaje de acuerdo a lo planteado en Oviedo et al. (2009); una cuenca interna profunda (Z máx=215m) de circulación superficial restringida, que se comunica al exterior del PTO por una zona externa poco profunda (Z máx=60m), con un umbral a 20km de la boca del Golfo, finalmente, una zona transición-oceánica que accede al Océano Pacífico y donde a una distancia relativa de menos de seis kilómetros se alcanzan profundidades cercana a los 1 000m. La hidrología compleja de este sistema es analizada en detalle en Quirós (2003) y en Svendsen et al. (2006).
Colecta y análisis de datos
El área de estudio se muestreó periódicamente en una embarcación de siete metros de eslora, con un motor cuatro tiempos de 115HP. Los muestreos que conforman este análisis abarcaron dos temporadas: seca (noviembre-mayo) y la de lluvia (junio-octubre), por un periodo aproximado de nueve años, desde marzo del 2005 hasta diciembre del 2014, los esfuerzos de búsqueda se iniciaron en periodos matutinos (07:00-08:00h) y culminaron a mitad de la tarde (14:00-16:00h). La unidad de muestreo; los avistamientos, incluye la definición de grupo tal como se plantea en Karczmarski, Wursig, Gailey, Larson y Vanderlip (2005). Cada avistamiento registrado se identificó al nivel de especie e incluso a nivel de subespecies, incluyendo las razas ecológicas que podrían ser formas potencialmente divergentes taxonómicamente, en base a la descripción detallada de las características diagnósticas en la literatura de referencia para el PTO (Baird, 2008; Jefferson, Webber & Pitman, 2008; Kato & Perrin, 2008; Macalpine, 2008; Perrin, 2008a b c; Wells & Scott, 2008), cuando no fue posible identificar el nivel taxonómico especifico, el avistamiento se clasificó a nivel de género y familia. De acuerdo a Wells y Scott (2008) y Pacheco-Polanco et al. (2011), para el Pacífico Tropical Oriental, las diferencias fenotípicas que permiten un diagnóstico de los ecotipos de Tursiops truncatus, en base a la morfología son el tamaño (forma costera de mayor tamaño y complexión que la forma oceánica) y la coloración (forma oceánica mucho más oscura y uniforme, forma costera con coloración clara que permite diferenciar tres capas). En el caso de Stenella attenuata graffmani (Perrin, 1975; 2001; 2008b), esta raza ecológica se distingue de S. a. attenuata por su mayor tamaño y complexión, así como un por un patrón de coloración con abundantes manchas, en contraste a la forma más estilizada y con menos manchas (incluso ausentes) de la forma oceánica. Cada 30min se realizó una lectura de posicionamiento por GPS junto con algunas variables ambientales, como la condición del mar (escala Beaufort), temperatura, ciclo de mareas y la presencia o no de cetáceos (Gowans & Whitehead, 1995). Lo anteriormente descrito permitió un registro del esfuerzo y cobertura en campo (Fig. 1), así como la generación de una matriz de presencia-ausencia de cetáceos.
Para cada encuentro el bote se situó a una distancia aproximada de 100m del grupo focal del avistamiento, posteriormente, datos como: la hora inicial y final del encuentro, tipo de especie, tamaño y composición de grupo, así como la conducta (inicial al encuentro y a los 10min del encuentro), fueron registrados junto con la posición espacial del bote, por lecturas de GPS (Garmin e-trex H, cinco metros de error aproximado), las cuales fueron equiparada como la posición relativa del grupo focal del avistamiento. El análisis de datos comprendió la evaluación de los avistamientos por medio de un sistema de información geográfica (Kiszka, Ersts & Ridoux, 2007; 2010), utilizando el programa ArcGIS 9.3 y la estimación de los índices de abundancia relativa (pi=Ni/N), así como el de diversidad de Simpson, de acuerdo a lo planteado en Flores-Ramírez, Urban, Villareal-Chávez & Valles-Jiménez (1996) y Carlo, Peter y Soetaert (1998). Se favoreció el índice de Simpson, dado su fácil interpretación y versatilidad, lo que permitió la posibilidad de establecer una probabilidad basada en el dominio de los tipos ponderados en el set de datos. Para determinar las diferencias a nivel de estructura de comunidades, entre pares de unidades de paisaje marino de Golfo Dulce (detallados en la descripción del área de estudio), se estimó un índice de complementariedad (CAB=UAB/SAB) entre la cuenca interna y el área del umbral, así como entre el área del umbral y la zona de transición oceánica, tal como está planteado en el análisis propuesto por Arellano y Halffter (2003). Para detalles sobre el cálculo de dichos índices, se sugiere revisar las referencias citadas anteriormente. Es importante mencionar que los ecotipos observados, se consideran en este análisis, como un componente aparte de la diversidad de cetáceos, sin que esto implique, bajo el nivel actual de conocimiento la diversificación taxonómica de estos morfotipos ecológicos, en base a lo anterior, las formas costeras y oceánicas del delfín nariz de botella, así como del delfín manchado pantropical se diversifican como parte de la riqueza de especies.
Resultados
Se obtuvieron 943 registros de avistamientos de cetáceos (2005-2014) constituidos por 12 especies, entre las que se incluyen ocho de la familia Delphinidae, incluyendo un morfotipo a nivel de sub-especies, que sería único para el PTO: S. longirostris centroamericana (anteriormente conocido como delfín tornillo de Costa Rica), también una especie de la familia Kogiidae (Kogia sima), un representante de la familia Ziphidae (Ziphius cavirostris) y dos representante de la familia Balaenopteridae: la ballena jorobada y el rorcual tropical (Cuadro 1).
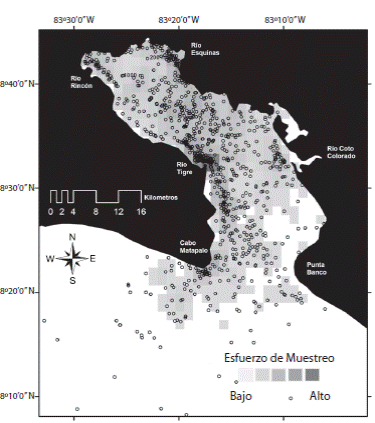
Fig. 1 Field effort distribution in Golfo Dulce: The study área is divided into cells of 1.6x1.6km (n=1 293). Spatial coverage is illustrate by means of a qualitative gradient scale, where the maximum effort invested implied 85h and 267km covered in each cell of 2.56km2. Sightings analyzed in this study are represented by unfilled circle, in contrast with the field effort coverage.
El delfín nariz de botella costero, se observó principalmente en la cuenca interna y en la zona del umbral (Figs. 2 y 3), mientras que en la zona de transición-oceánica, la forma oceánica de T. truncatus fue la especie de delfín más común (Fig. 4). El delfín manchado pantropical presente en la cuenca interna de Golfo Dulce, correspondería con la forma costera (S. a. graffmani), en base a su distribución regional en el PTO, sin embargo, encuentros en la zona de transición-oceánica, muestran diferencias externas en morfología y patrón de manchas entre los ecotipos observados dentro y fuera del Golfo Dulce (Fig. 5), los delfines manchados pantropicales en la cuenca interna del golfo (individuos adultos), serian intermedios en dimensiones corporales y patrón de manchas entre los ecotipos S. a graffmani y S. a attenuata.
El cuadro 1 muestra los resultados en la riqueza de especies, la abundancia relativa en porcentaje, el esfuerzo de búsqueda y el índice de diversidad de Simpson del grupo indicador, de manera diversificada en unidades estructurales del paisaje marino en Golfo Dulce. El índice de diversidad de Simpson refleja un gradiente desde la cuenca interna hasta la zona de transición oceánica, mientras que el índice de complementariedad sustenta dicho gradiente, al ilustrar diferencias medias en composición faunística entre la cuenca interna y la zona del umbral (CAB=0.50) y notablemente abruptas entre el umbral y la zona de transición oceánica (CAB=0.93).
Cuadro 1 Resultados pertinentes a la estructura del paisaje, la riqueza de especies por unidad de paisaje, los índices de abundancia relativa (porcentaje), esfuerzo de búsqueda (horas) y diversidad de Simpson
| Unidad de Paisaje | Riqueza de Especies (n) | Esfuerzo de Busqueda (horas) | Abundancia Relativa (%) | Índice de Simpson |
|---|---|---|---|---|
| Cuenca Interna | Tt (283) | 1 919 | 50 | 0.60 |
| - | Sa (204) | - | 36.04 | - |
| - | Mn (75) | - | 13.25 | - |
| - | Pc (4) | - | 0.71 | - |
| Área del Umbral | Tt (124) | 830 | 41.61 | 0.67 |
| - | Sa (77) | - | 25.84 | - |
| - | Mn (92) | - | 30.87 | - |
| - | Pc (4) | - | 1.34 | - |
| - | Sb (1) | - | 0.34 | - |
| Zona de Transición Oceánica | Sag (13) | 741 | 17.11 | 0.70 |
| - | TtO (31) | - | 40.79 | - |
| - | Sl (7) | - | 9.21 | - |
| - | Sb (14) | - | 18.42 | - |
| - | Lh (1) | - | 1.32 | - |
| - | Dd(1) | - | 1.32 | - |
| - | Gm (3) | - | 3.95 | - |
| - | Ks (2) | - | 2.63 | - |
| - | Zc (2) | - | 2.63 | - |
| - | Oo(1) | - | 1.32 | - |
| - | Ba(1) | - | 1.32 | - |
Tt=forma costera delfín nariz de botella T. truncatus, Sa=delfín manchado pantropical S. attenuata (ecotipo potencial), Mn=ballena jorobada M. novaeangliae, Pc=falsa orca P. crassidens, Sb=delfín dientes rugosos S. bredanensis, Sl=delfín tornillo Centro-Americano S. longirostris centroamericana (sub-especie Centroamericana), Lh=delfín de Fraser Lagenodelphis hosei, TtO=forma oceánica delfín nariz de botella, Sag=delfín manchado S. attenuata graffmani, Dd=delfín común D. delphis, Gm=ballena piloto G. macrorhynchus, Ks=cachalote enano K. sima. Zc=ballena de Cuvier Z. cavirostris, Oo=Orca O. orca y Ba=rorcual Balaenoptera sp.
Tt=in shore bottlenose dolphin T. truncatus, Sa=pantropical spotted dolphin S. attenuata (potential ecotype), Mn=humpback whale M. novaeangliae, Pc=false killer whales P. crassidens, Sb=rough toothed dolphin S. bredanensis, Sl=spinner dolphin S. longirostris centroamericana (Central American sub-especie), Lh=Fraser´s dolphin Lagenodelphis hosei, TtO=off-shore bottlenose dolphin, Sag=coastal pantropical spotted dolphin S. attenuata graffmani, Dd=common dolphin D. delphis, Gm=pilot whale G. macrorhynchus, Ks=dwarf sperm whale K. sima. Zc=Cuvier´s beaked whale Z. cavirostris, Oo=Orca O. orca y Ba=rorcual whale Balaenoptera sp.
La diversidad de la cuenca interna se restringe a cuatro especies, dos de las cuales son residentes: delfín manchado y nariz de botella, así como una especie residente temporal, Megaptera novaeangliae, adicional a la incursión ocasional de la falsa orca. Las mismas cuatro especies observadas en la cuenca interna se adicionan a un delfínido pelágico; el delfín de dientes rugosos (Steno bredanensis) para constituir la riqueza de especies presentes en la zona correspondiente al umbral (Fig. 3). Un cambio más abrupto en composición faunística se produce en la zona de transición-oceánica, donde las formas oceánicas prevalecen, incluyendo especies predominantemente teutófagas: zifio de Cuvier (Z. cavirotris), ballenas pilotos (Globicephala macrorhynchus) y cachalote enano (K. sima), en contraposición con el dominio de especies de delfinidos de naturaleza más piscívoras (Fig. 4) y la presencia de depredadores de nivel tope como la ballena orca (Orcinus orca).
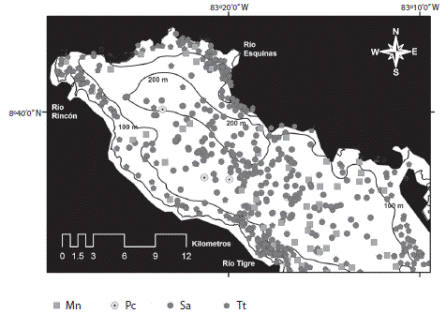
Fig. 2 Cetacean community structure at the inner basin of Golfo Dulce (2005-2014): Tt=in-shore bottlenose dolphin, Sa=pantropical spotted dolphins (potential ecotype), Mn=humpback whale, Pc=false killer whale.
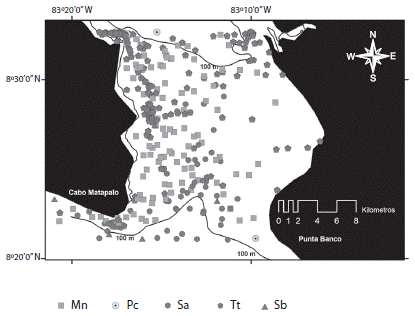
Fig. 3 Estructura de la comunidad de cetáceos en el área del umbral de Golfo Dulce (2005-2014): Tt=delfín nariz de botella, Sa=delfín manchado pantropical (ecotipo potencial), Mn=ballena jorobada, Pc=falsa orca, Sb=delfín dientes rugosos. Fig. 3. Cetacean community structure at the sill area of Golfo Dulce (2005-2014): Tt= in-shore bottlenose dolphin, Sa=pantropical spotted dolphin (potential ecotype), Mn=humpback whale, Pc=false killer whale, Sb=rough toothed dolphin.
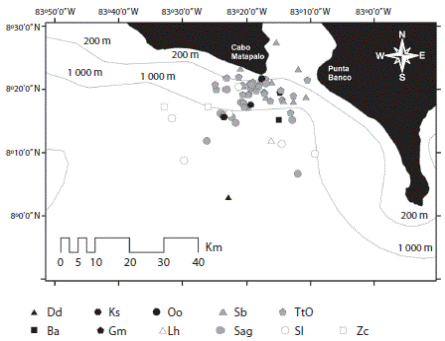
Fig. 4 Cetacean community structure at the transitional-oceanic area of Golfo Dulce (2005-2013): TtO=off-shore bottlenose dolphin, Sag=pantropical spotted dolphin (subespecie: S. attenuata graffmani), Sb=rough toothed dolphin, Sl=spinner dolphin (Central American sub-especie), Lh=Fraser´s dolphin, Dd common dolphin, Gm=short finned pilot whale, Ks=dwarf sperm whale, Zc=ballena de Cuvier, Oo=Orca y Ba=ballena rorcual.
Discusión
Este estudio documenta la notable diferencia entre la composición de especies de cetáceos entre la cuenca interna, el umbral y la zona de transición oceánica de Golfo Dulce. Dichas diferencias en el patrón de diversidad de cetáceos en Golfo Dulce, estarían en líneas generales determinadas por: (a) limitantes en la posibilidades de dispersión; (b) limitantes eco-geográficos; y finalmente (c) la dinámica interna de la comunidad de cetáceos, la cual estaría dada por atributos colectivos como la riqueza de especies y la biomasa, así como atributos de tipo emergentes como la coexistencia de especies y la estabilidad de la trama trófica ante posibles alteraciones (Begon, Townsend & Harper, 2006).
La riqueza de especies cetáceos en Golfo Dulce y el nivel de complementariedad entre las tres sub-áreas permiten describir tres comunidades:
Cuenca interna
El dominio de esta comunidad de cetáceos lo ejercen las dos especies residentes; el delfín manchado y la forma costera de delfín nariz de botella. La estructura interna y las características fisiográficas de esta área facilitarían la diferenciación de nichos que se refleja en la distribución de ambas especies (Acevedo-Gutiérrez & Burkhart, 1998; Cubero-Pardo, 1998; 2007ab; Oviedo, 2007; Oviedo, 2008). Elementos como la pendiente y la profundidad, son aspectos que permiten interpretar los procesos a lo que depredadores como delfines estarían respondiendo; por ejemplo, a la abundancia de presas y la repartición de nichos, lo que representaría una estrategia de coexistencia en Golfo Dulce (Oviedo, 2007), un fundamento clave en la estructuración de la comunidad. Dos aspectos importantes complementan la estructura de esta comunidad: 1) la incursión de un depredador tope como la falsa orca (Pseudorca crassidens), potencialmente siguiendo la migración de sus presas y 2) el uso de hábitat de una especie migratoria como la ballena jorobada (M. novaeangliae).

El área del umbral
De acuerdo a la riqueza de especie y a la complementariedad de la misma con la cuenca interna, en conjunto con las características un poco más homogénea de esta zona, el área del umbral se definiría como una zona clave de transito de especies que incursionan a la cuenca interna regularmente (delfines manchado y nariz de botella, así como la ballena jorobada), incluso aquellas especies que ingresan ocasionalmente siguiendo presas potenciales como en el caso de P. crassidens. El avistamiento reciente de delfines dientes rugosos (19/03/2013) en la zona de umbral confirma la incursión y uso por parte de especies comunes a la zona de transición-oceánica en aguas someras dentro del Golfo Dulce.
La comunidad de cetáceos de la zona del umbral estaría dominada por la presencia de especies residentes, en particular T. truncatus con la extensión de sus áreas de alimentación por la influencia de ríos y quebradas en la zonas aledañas entre Puntarenitas y Cabo Matapalo. La otra especie clave seria la ballena jorobada, con un uso intensivo de dos subpoblaciones migrantes: hemisferio sur, durante los meses de invierno austral (julio-octubre: M. n. australis), junto con las ballenas que proviene del hemisferio norte (M. n. kuzira) durante finales de Noviembre a mediados de Marzo (Márquez-Artavia et al., 2012; Jackson et al., 2014).
La zona de transición oceánica
Esta área representa el contraste más abrupto en la estructura de la comunidad de cetáceos, enfatizado por la poca complementariedad con la zona del umbral y cuenca interna. El dominio en cetofauna lo ejercen delfínidos como las forma oceánica de T. truncatus y la subespecie del delfín manchado S. a. graffmani, complementados por especies relevantes como S. l. centroamericana, Delphinus delphis y S. bredanensis. La presencia de cetáceos de apnea profunda como Z. cavirostris, G. macrorhynchus y K. sima, evidencia la oferta de nichos para especies teutófagas (Pauly, Trites, Capuli & Christensen, 1998). Esta área es igualmente utilizada por depredadores tope de carácter transitorio, tal como se evidencia con registros documentados y publicados sobre la presencia de Orcinus orca, (Ferlt, Acevedo-Gutiérrez & Darby, 1996) y los actuales de esta especie en la base de datos de este estudio.
La estructura del hábitat pareciese ser mucho más uniforme, en consecuencia, se esperaría niveles importantes de exclusión competitivas que incidirían en la conformación de la comunidad de cetáceos de esta área, particularmente entre las especies que dominan, que son predominantemente piscívoras, sin embargo, la conducta de agregación típica de delfínidos oceánicos (Gowans, Wursig & Karczmarski, 2008), junto con la distribución heterogénea del recurso presa, harían posible la coexistencia sin una diferenciación de nichos aparente (Wertheim, Sevenster, Eijs & van Alphen, 2000).
La complementariedad entre sub-áreas sustenta la diversificación de las comunidades descritas anteriormente, particularmente entre cetáceos en aguas internas del Golfo Dulce y el Océano Pacífico, donde el cambio estructural en hábitat y en riqueza de especie es más notable. Un aspecto a resaltar es el uso de hábitat de depredadores transitorios como la falsa orca y la orca en el marco de esta complementariedad, con un rango hogar extenso que incluso abarca ámbitos fuera de las aguas de la Península de Osa.
Formas geográficas y ecotipos
Es importante enfatizar en la divergencia potencial en formas geográficas o eco-tipos de especies representativas a nivel del paisaje de forma integral. Pacheco-Polanco et al. (2011) documento la presencia de las formas costeras y oceánicas del delfín nariz de botella, en base a diferencias morfológicas en tamaño y coloración previamente documentadas para otras áreas dentro del PTO (Wells & Scott, 2008), esta contribución ofrece evidencias claras de cómo cada una de estas formas, ejercen un dominio espacial en las comunidades y hábitats donde se presentan: el ecotipo costero del delfín nariz de botella en la cuenca interna y el área del umbral de Golfo Dulce y el ecotipo oceánico de la misma especie en la zona de transición oceánica fuera del Golfo. Las evidencias fotográficas que se aportan en este estudio, indicarían también posibles diferencias fenotípicas en el delfín manchado pantropical, siendo la forma presente en las afueras de la cuenca interna y el área umbral consecuente con la descripción de S. a. graffmani (Perrin, 2001; 2008b), en relación al tamaño y al patrón considerable de manchas, mientras que la forma en la cuenca interna difiere en tamaño, siendo estos delfines más pequeños y estilizados con manchas menos resaltantes.
Proponemos la hipótesis sobre la posible presencia de un morfotipo de S. attenuata para Golfo Dulce, en base a diferencias fenotípicas consistentes y al hecho de que tales diferencias parecen corresponder, no solo con lo propuesto por Escorza-Treviño, Archer, Rosales, Lang y Dizon (2005), en términos de sub-poblaciones y unidades de manejo (incluyendo una para Costa Rica), sino con lo sugerido por el mapa de distribución de la especie en aguas de la Península de Osa y Golfo Dulce por May-
Collado et al. (2005), con la notación particular de sub-especie desconocida. Los esfuerzos actuales en determinar estructura poblacional de S. attenuata en Golfo Dulce, progresarían a la determinación de un perfil genético comparativo para esta especie, para elucidar la presencia o no de un ecotipo de delfín manchado pantropical para Golfo Dulce.
Finalmente, la proximidad de diferentes tipos de hábitat en aguas dentro y fuera del Golfo Dulce determina este sistema, como un área importante de abundancia de cetáceos, especialmente para una notable comunidad diversificada de odontocetos. Estudios más precisos en uso de hábitat, tasa de encuentros, abundancia absoluta y diversificación de nichos, están actualmente en progreso con el fin de informar y promover una zonificación efectiva que permita mitigar conflictos potenciales por uso de hábitat entre mamíferos marinos y comunidades costeras humanas. Esta información es vital en la fase de planeamiento y análisis de las estrategias de gestión para Golfo Dulce, de acuerdo a St Martin & Hall-Arber (2008), es la etapa donde la georeferencia detallada del paisaje marino y sus elementos asociados, entre estos la diversidad (Crowder & Norse, 2008; Foley et al., 2010), permite ampliar el entendimiento de la heterogeneidad de las comunidades biológicas y sus componentes claves.

