











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkIn the world, approximately 151 million hectares are cultivated with rice (FAO Stat, 2008). In Argentina, irrigated rice production is carried out mainly in Corrientes and Entre Ríos Provinces because of the agroecological characteristics (water, climate and soil) of this region, and nowdays, an approximate of 164 300 hectares are planted with this crop in both Provinces (Asociación Correntina de Plantadores de Arroz y Bolsa de Cereales de Entre Ríos, 2013). The irrigated rice fields behave as temporary wetlands, with alternating periods of flooding in summer and drought in winter (Fasola & Ruiz, 1996). These rice fields act as artificial aquatic ecosystems which connect and share water with natural wetlands (rivers and water courses, groundwater or dams filled with rainwater) (Blanco & Balze, 2011). These man-made environments, are included in the definition of wetlands adopted by the Wetlands Convention (Secretaría de la Convención de Ramsar, 2010), and harbor a great variety of aquatic invertebrates (Fernando, 1993; Fernando, Furtado, & Lim, 1979), including insects (Bambaradeniya, 2000; Bambarade-niya et al., 2004). Most aquatic invertebrates, mainly microcrustaceans, insect larvae, gastropods, and oligochaetes, contribute to nutrient cycling in rice fields, and play important roles in the decomposition of the photosynthetic aquatic biomass that develops in rice field floodwater (Simpson & Roger, 1991).
In South America, studies about aquatic insects in rice fields are scarce. Some studies about the arthropod community have been carried out in Southern Brazil (Stenert, Bacca, Maltchik, & Rocha, 2009; Fritz et al., 2011); besides, a paper on rice fields biodiversity was conducted in South-Central Corrientes Province (Blanco & Balze, 2011). Although beetles are important components of the aquatic fauna, little is known about their ecology in these artificial ecosystems in this study area. A pioneering study by Yano, Chu, and Sato (1983) documented 117 species of water beetles belonging to 14 families in rice fields worldwide. Most of the studies on the biodiversity of aquatic insects in rice fields have been conducted in Asian countries (e.g. Rozilah & Ali, 1998; Bambaradeniya & Amerasinghe, 2003; Bahaar & Bhat, 2011), probably because Asia stands out as the largest producer and consumer of rice in the world.
Abundance and species richness of aquatic beetles have been reported to be strictly related to the physical variables of the habitat such as vegetation and the chemistry of water (e.g. Nilsson & Soderberg, 1996). Most studies in this type of environment have dealt mainly with agronomic aspects and rice pests; however, studies on biodiversity in these agroecosystems fauna remain scarce. The aims of this study were to determine the diversity and species richness of coleopterans in a typical rice field, and to characterize the community of water beetles through different abundance models.
Materials and Methods
Study site: This study was conducted in a rice field located 30 km Southwest of Corrientes city, Argentina, in the "El Sombrero" town (27°42'12" S - 58°46'2" W). The site has belonged to the National Institute of Agricultural Technology (INTA) since 1958. This field has an area of ~1 175 ha. Agricultural and livestock activities are carried out in the site.
Irrigated rice production: Rice sowing began in September and October. The field was then flooded with water drawn from the Paraná River, pumped from the main and secondary channels. The water is maintained on the plots by containment edges called "taipas" until April, and it is drained for later harvest.
In the initial phase of flooding, no vegetation was observed in the irrigation channels; later on, the predominant hydrophilic vegetation consisted of Sagittaria montevidensis Cham. and Schlecht, Hydrocotyle ranunculoides L. f., Limnobium sp. and Ludwigia sp.
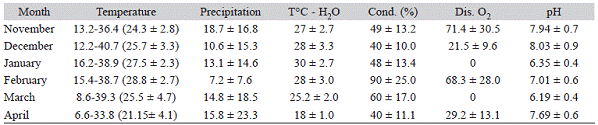
Sampling methods: Samples were carried out monthly during a rice cultivation cycle (November 2011-April 2012). Aquatic insects were collected with an aquatic hand net (mesh size 300 Lim, diameter 30 cm) both inside the rice field and in channels that drain the water from the river into the plantation, and connect to the different sowed areas. The sampling was started immediately after the field preparation work, which coincided with spring and sowing of the rice. The random sampling technique was used in different parts of the site, usually between 10 a.m. and 1 p.m. The samples were taken manually by one person who sampled for one hour, from the cultivated parcels and irrigation channels, in order to avoid underestimation of the biodiversity. During the sampling months, water depth ranged between 5 and 10 cm in the cultivated parcels, and between 10 and 50 cm in the irrigation channels. Table 1 shows data on monthly temperature, precipitation, water temperature, conductivity, dissolved oxygen and pH for this period.
The insect samples were fixed in situ in formaldehyde 5 %, transferred to the laboratory for identification, and then stored in 70 % etha-nol. Taxa were classified according to Lawrence and Newton (1995). Hydrophiloidea was classified following Hansen (1999) and Short and Fikácek (2011). Hydrophilidae was classified following Short and Fikácek (2013), whereas Dytiscidae was classified following Nilsson (2001). Species were identified following Tré-mouilles and Bachmann (1981), Trémouilles (1984, 1989, 1995), Trémouilles, Oliva and Bachmann (1995), Trémouilles, Michat and Torres (2005), Oliva, Fernández and Bachmann (2002) and Vondel and Spangler (2008). The material collected was deposited in the collection of the Centro de Ecología Aplicada del Literal (CECOAL-(CONICET-UNNE)), Corrientes Province.
Table 1:Temperature (°C): minimum-maximum (mean ± standard deviation); mean precipitation (mm) ± SD; water temperature (°C) ± SD, conductivity (%) ± SD, dissolved oxygen ± SD and pH ± SD during a rice cultivation cycle in a rice field of Corrientes province, Argentina
Only adult individuals were taken into consideration, for different analysis. Richness and diversity species were calculated on a monthly basis. The diversity was calculated by the Shannon diversity index (Shannon & Weaver, 1949). Rarefaction methods were first used to compare the average diversity and the average richness between the different months. Rarefaction uses a probability theory to derive expressions for the expectation and variance of diversity and richness of species for a sample of a given size. This method lowers the sample to a common abundance level and then compares species diversity and richness. Ecosim 7.0 software (Gotelli & Entsminger, 2004) was used for this calculation. The richness and diversity results were obtained for 21 classes of abundance (10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200 and 205 individuals). Subsequently, the diversity and species richness for each of the 21 classes of abundance was compared through the non-parametric Kruskal-Wallis test and then by the Dunn test with the Bonferroni correction. The statistical analyses were calculated with XlStat 7.5 software (Addinsoft, 2006).
The equitability for each month sampled was calculated by the formula of the Shannon equitability index (Magurran, 2004).
The community composition for each sampling date was characterized by means of rank-abundance models. Three of the most common series (logarithmic, geometric or "broken stick") presented a better adjustment to the data that was specified for each case. The different variables were compared by means of the Chi-square test. Every calculation was made with the Bio-Dap software (Thomas & Clay, 2000), which is based on the methods proposed by Magurran (1988).
Results
A total of 74 species of Coleoptera, belonging to 28 genera and seven families, were identified. Dytiscidae was the richest genera family, while Hydrophilidae was the richest species family (Dytiscidae: 13 genera; 22 species, Hydrophilidae, 7 genera; 36 species). The following richest families were Noteridae (3 genera; 9 species), Dryopidae (2 genera; 3 species), Hydrochidae (1 genus; 2 species), Haliplidae (1 genus; 1 species) and Gyrinidae (1 genus; 1 species) (Table 2).
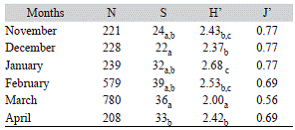
In order to compare richness and diversity of the aquatic Coleoptera community among the six months sampled, the parameters were rarefied to the smallest individual number (April, N = 205). Significant differences were observed in diversity and species abundance in the six months analyzed (Kruskal-Wallis Test; K = 66.94, d.f = 5, P < 0.05, and K = 19.37, d.f. = 5, P < 0.05, respectively). According to the Dunn Test with Bonferroni correction (= 0.0033), the diversity in January and February was significantly higher than in the other months, where diversity showed similar lower values. The species richness during December and March was significantly higher than in April, whereas in the other months was similar (Table 3).
Table 2: Aquatic Coleoptera species collected in a rice field located in Corrientes province (Argentina) from November 2011 to April 2012
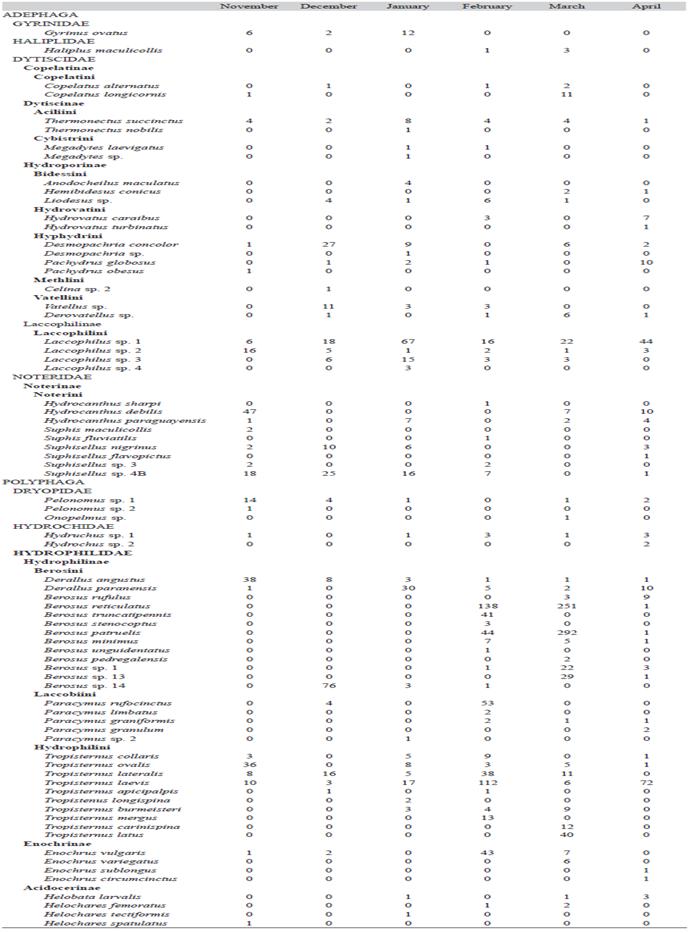
Table 3: Richness parameters summary (N): number of individuals; (S): species richness; (H): Shannon Index, and (J'): Equitability Index. Dunn test with Bonferroni correction for diversity results (calculated using the Shannon Index) and species richness in the rice field
*Same letters indicate no significant differences; different letters indicate significant differences between groups
The greatest number of species was recorded in February (S = 39); and the highest abundance was observed in March (N = 780) (Table 3). The greatest diversity was recorded in January (H' = 2.68) and the lowest in March (H' = 2.00). The Shannon equitability index indicated that evenness of most of the months sampled was high (J' < 0.69), except for March (J' = 0.56) (Table 3).
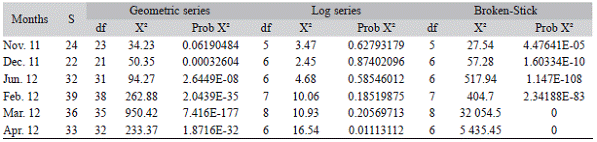
The distribution of species abundance in the different months was adjusted to a log-series model; nevertheless, the April distribution of species abundance did not fit any of the series models (Table 4). The test of goodness of fit (Chi-square test) indicated that no other model fitted well the data of species distribution in the others analyzed communities.
Discussion
Previous studies recognize the Coleoptera as the most diverse insect order living in rice fields in the province Corrientes (Blanco & Balze, 2011). In the present study, the aquatic coleopteran assemblage was characterized by the presence of Dytiscidae, Hydrophilidae, Noteridae, Dryopidae, Hydrochidae, Halipli-dae and Gyrinidae, also reported in several rice field studies (e.g. Leitäo, Pinto, Pereira, & Brito, 2007; Stenert et al., 2009; Bahaar & Bhat, 2011). However, the most representative families regarding their abundance were Hydrophilidae, Dytiscidae and Noteridae. Similar results were found in a study carried out in two permanent ponds in Corrientes province, where eight families were recorded, and where Hydrophilidae (48%), Dytiscidae (27%) and Noteridae (18%) showed the highest species percentage (Gómez Lutz, Fernández, & Kehr, 2012).
Hydrophilidae has also been reported as the family with significantly higher number of species among aquatic Coleoptera (Bahaar & Bhat, 2011; Gómez Lutz et al., 2012), and Berosus and Tropisternus have been the most important genera. Similar results have been obtained in permanent ponds from Corrientes Province, where both genera had more species number inside Hydrophilidae (Gomez Lutz et al., 2012). Berosus reticulatus, Berosus patrue-llis and Tropisternus laevis were the most abundant during February and March in this study. It is possible that rice fields in this area provide similar natural environmental conditions to the aquatic Coleoptera community.
Different studies on the fauna have also showed that these artificial environments may harbor a fauna similar to undisturbed wetlands (Bambaradeniya et al., 2004; Duré, Kehr, Schaefer, & Marangoni, 2008; Maltchik, Rolon, Stenert, Machado, & Rocha, 2011). These rice fields are a refuge to a great variety of organisms, all well adapted to this temporarily and highly manipulated ecosystem that is periodically disrupted by various agricultural practices (Fernando, 1995). The community of water beetles was rich and abundant during the study, and similar to one reported for a national park (128 species included in 44 genera and seven families) and natural environments of this Province (127 species included in 40 genera and eight families) (Torres et al., 2012; Gómez Lutz et al., 2012). Most of the months sampled showed high diversity; the highest diversity index was observed in January. March was the month with the highest species richness but also the month with the lowest value of diversity. In contrast, January showed an intermediate number of species but the highest diversity index of all months. All these species were similarly represented. The equitability test was generally high, which showed a heterogeneous representation in the number of individuals within the community. However, it is important to note that this lack of homogeneity could be given by an explosion in the abundance of two Berosus species in that month. January and February are the most favorable months for studies related with the coleopteran communities in the rice field, since during these months the highest richness and diversity were registered.
The rank-abundance models aided the interpretation of the data. The most commonly used models are the geometric, the logarithmic and the "broken stick"; all of them were required to fit data from the different months of rice cultivation. The geometric series describes communities of highly uneven species-abundance distribution and low diversity characterized by a few dominant species; the "broken stick" model characterized by reflecting a more equitable condition among species and for having a more uniform distribution (Magurran, 1988); and the logarithmic series fitted better to communities characterized as being influenced by only one or a few determining factors affecting ecological interactions, with a high proportion of species found to be very uncommon (Magurran, 1988). The analyzed communities in most of the months (except April) fitted only the rank-abundance models of the logarithmic series. This is where the species with intermediate abundance were those which dominated the different analyzed communities.
In a study of artificial environments in Argentina, Schnack, Francesco, Colado, Novoa, and Schnack (2000) suggested that these environments would contribute preserve the diversity. Our results on the diversity and richness of water beetles species in rice fields, showed that these can function as important systems that may contribute to biodiversity conservation, especially in those widespread areas where these environments have become part of the landscape.
The results suggest that the rice irrigation fields in Corrientes Province can host a wide richness of aquatic Coleoptera. This feature would be justified because the community structure was characterized by being conformed by both species, common and uncommon, (logarithmic model features) with similar abundances and slightly fluctuating. Therefore, these environments could be an alternative for biodiversity conservation in the region where the rice fields expand rapidly, due to the optimal production characteristics of soil and climate in the province. Understanding the factors regulating coleopteran abundance and diversity in agricultural areas will help to elucidate possible effects of human activities and the perturbations on natural populations.

