











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.63 no.2 San José abr./jun. 2015
The stands of Copernicia tectorum (Arecaceae) in the Caribbean lowlands of Colombia: a managed pioneer palm facing river dynamics
Poblaciones de Copernicia tectorum (Arecaceae) en las tierras bajas del Caribe de Colombia: una palma pionera expuesta a la dinámica de un río
Poblaciones de Copernicia tectorum (Arecaceae) en las tierras bajas del Caribe de Colombia: una palma pionera expuesta a la dinámica de un río
Abstract
Copernicia tectorum is a palm that grows in large populations on seasonally flooded savannas in the Caribbean region of Colombia, where its stems and expanded leaves are used in construction, and its unexpanded leaves are used to make handicrafts. We studied abundance and population structure in 34 plots of 20×10m (0.68 ha) of three localities (Plato, Córdoba and Magangué) at the Mompox Depression, an inner delta formed by the confluence of four large rivers. We recorded growth and mortality of 164 palms of different size classes over 13 months, and additionally we estimated seedling and juvenile mortality in two 10x10m plots within the same sites. Data analysis using Kolgomorov-Smirnov (KS), Kruskal-Wallis and Mann-Whitney tests were made on Statgraphics Plus and SPSS. We related the structure and dynamics of the palm stands to flood intensity. Copernicia tectorum has the fastest leaf production rate recorded for any palm (19-23 leaves/year in subadults and adults), and a short life span of ca. 46 years. The abundance, density and high leaf production rate of this palm offer a great potential for the sustainable use of its unexpanded leaves (especially at Plato, where there are ca. 480ha of palm stands with 300-1 000 individuals/ha), as leaf harvest from subadult and adult individuals does not appear to affect population structure. The palm is a pioneer of the flood plain, and the palm stands at particular sites appear to wax and wane following sedimentation dynamics, in which the species plays in important role. Current population structure and the scarce recruitment of juveniles at the study site suggest that the population is waning in that site. Management actions should be taken to reestablish natural river dynamics at the lagoon complex where the palm grows. Appropriate management of palm stands may contribute to minimize any negative effects of river dynamics.
Key words: leaf harvest impact, leaf production, Mompox Depression, NTFP, pioneer plants, population structure, seasonally flooded savanna.
Resumen
La palma Copernicia tectorum forma grandes poblaciones en sabanas estacionalmente inundables de la región Caribe de Colombia, donde sus tallos y hojas se utilizan en construcción y sus cogollos para la elaboración de artesanías. Estudiamos la abundancia y la estructura poblacional en 34 parcelas de 20×10m (0.68 ha), en tres localidades (Plato, Córdoba y Magangué) en la Depresión Momposina, un delta interior formado por la confluencia de cuatro grandes ríos. Registramos el crecimiento y la mortalidad de 164 palmas de diferentes clases de tamaño durante 13 meses y estimamos además la mortalidad en plántulas y juveniles en dos parcelas de 10x10m en los mismos sitios. Los análisis se realizaron con las pruebas de Kolgomorov-Smirnov (KS), Kruskal-Wallis y Mann-Whitney en Statgraphics Plus and SPSS. Relacionamos la estructura y dinámica de los palmares con la intensidad de la inundación. Copernicia tectorum tiene la tasa de producción de hojas más rápida registrada hasta ahora para cualquier palma (19-23 hojas por año en subadultos y adultos) y una vida corta de ca. 46 años. La abundancia, densidad y alta tasa de producción de hojas de la palma ofrecen un gran potencial para el uso sostenible de sus cogollos (especialmente en Plato, donde hay unas 480 ha de palmares con 300-1 000 individuos/ha), ya que la cosecha de hojas de palmas adultas y subadultas, parece no afectar la estructura de la población. La palma es una pionera de las llanuras inundables, y los palmares en un sitio particular crecen y se reducen siguiendo la dinámica de sedimentación, en la cual la especie juega un papel importante. La estructura actual de las poblaciones y el escaso reclutamiento en las clases juveniles en el sitio de estudio sugieren que allí la población se está reduciendo. Se deben emprender acciones de manejo que apunten a restablecer la dinámica fluvial natural en los complejos cenagosos en los que crece la palma. A su vez, el manejo apropiado de los palmares puede ayudar a minimizar los efectos negativos de la dinámica fluvial.
Palabras clave: Depresión Momposina, estructura poblacional, impacto de la cosecha, plantas pioneras, PFNM, producción de hojas, sabana estacionalmente inundable.
Fibers obtained from wild palms play a significant role in Colombian handicraft production (Linares, Galeano, García, & Figueroa, 2008). One of the most important fiber-producing palms in the Caribbean lowlands is the sará palm, Copernicia tectorum (Kunth) Mart., which grows only in dry savannas in Northern Colombia and Venezuela, where it forms extensive stands in areas that are subject to severe drought during the dry season, and to permanent flooding during the rainy season (Troth, 1987; Galeano, & Bernal, 2010). In some lagoon complexes at the Mompox Depression, in Northern Colombia, this species is the dominant element in the vegetation. In these areas the palm has a major ecological role, due both to its abundance and to the numerous interactions with other organisms-it harbors termites, wasps, ants, birds, and epiphytes, and its fruits are food for fish, birds, and mammals (Troth, 1987; Torres, 2013; pers. obs.).
The sará palm also has a great importance to local communities, as its leaves and stems are used in construction, and fiber obtained from its unexpanded leaves is the base of a flourishing handicraft activity of local importance. In the municipalities of Magangué (department of Bolívar) and Plato (department of Magdalena), along the Magdalena river, at least 200 families are involved in this handicraft production (Linares et al., 2008). However, the ongoing process of deforestation and lagoon desiccation for establishing crops and for cattle ranching has reduced the palm populations, causing a decrease of fiber availability and a greater pressure on the surviving palm stands (Linares et al., 2008; Torres, 2013). Sustainable management of this species requires an understanding of the stands, their role in these savannas and lagoon complexes, and the possible impact of leaf harvest on populations.
Extraction of non-timber forest products (NTFPs) has been the subject of a long debate as to the scope and extent of the research that is required to guarantee sustainability (Peters, 1996; Wong, Thornber, & Baker, 2001; Ticktin, 2004). There seems to be a consensus, however, about the need of basic biological and ecological studies, as a starting point to plan sustainable management, even for those species that have been traditionally used. The required basic information includes abundance, density, population structure, production rate of the plant organ used, and the plant’s overall growth rate (Peters, 1996), as well as an assessment of any other ecosystem services involving the used species, and its role in the ecosystem. A first sign of unsustainable harvest is evidenced in size class distribution (Peters, 1996; Bernal, & Galeano, 2013). Although for many plants the ideal size class distribution has an inverted-J shape (many seedlings and a decreasing number of juveniles and adults), other size class distribution curves are found among species growing in open areas or among those subject to extreme environmental changes (Peters, 1996). Because of this, an assessment of harvest impact must be based on the study of both harvested and unharvested populations (Bernal, & Galeano, 2013).
In this paper we used data on abundance, density, and size class distribution and plant growth, to assess the state of populations of Copernicia tectorum under various harvest regimes at three localities in the Mompox Depression, in the Caribbean region of Colombia. We also present data on the impact of river floods on the populations, and describe a plausible scenario of changes in the palm stands in the long run.
Materials and methods
Study site: We studied populations of C. tectorum around Ciénaga Zárate, and Ciénaga Cascaloa, a complex of lagoons in the floodlands of the lower Magdalena river, near the towns of Plato and Magangué, in the departments of Magdalena and Bolívar, in Northern Colombia. This area is part of the Mompox Depression (Depresión Momposina), Colombia’s largest wetland area, considered as an inner delta formed by the confluence of four large rivers -Magdalena, Cauca, Cesar, and San Jorge (Plazas, Falchetti, Van der Hammen, & Botero, 1988). This area is classified as tropical dry forest in Holdridge’s life zone system (Instituto Geográfico Agustín Codazzi, 1977), with an average temperature of 28°C, and 1 250-1 500mm annual rainfall. Intense rains fall during March-May and August-December, and there are severe droughts during June-July and December-February. Flood dynamics throughout the seasons determines a water flow from the Magdalena river to the lagoons and vice versa. Floods follow the rainfall regime of the river basin’s midlands and highlands, which is mostly bimodal, with a peak from mid-April to early June, and a second, higher one, from mid-August to December. As a result, there is a first flood between May and July and a second one between October and December. However, flood frequency, duration and level have changed in recent years, as a result of road construction, lagoon dessication, and the illegal use of lagoons and shores (Ministerio del Medio Ambiente [MMA], 2002). This is evidenced by the fact that the hydrological station at Plato recorded for the Magdalena river in February 2010 a lower level of 4.2m, close to the lowest value ever recorded (4.0m in 1977), and well below the average for that month (6.5m); similarly, in December 2010 there was a maximun level of 12.8m, well over the average for that month (9m) (Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia, 2011).
Study species: Copernicia tectorum is a solitary palm with an erect stem 5-10m tall, 20-30cm in diameter, which remains covered for some time with persistent leaf bases (Galeano, & Bernal, 2010). It has a mass of adventitious roots at the base, which have been considered as an adaptation to whithstand the long floods (Throth 1987). The crown has 15-25 palmate, circular leaves composed of numerous rigid segments up to 1 m long, with up to 1.6 m long petioles that bear hooked spines along their margins. Inflorescences are interfoliar and project beyond the leaves. They have numerous branches and bear small, hermaphrodite flowers. Fruits are ovoid to obovoid, ca. 3cm long, bright black when ripe, and have one seed. In Colombia this species grows only in the Caribbean lowlands (Galeano, & Bernal, 2010). It is unusual in its adaptation to extreme conditions of flooding and drought and its resistance to fire (Troth, 1987) (Fig. 1).
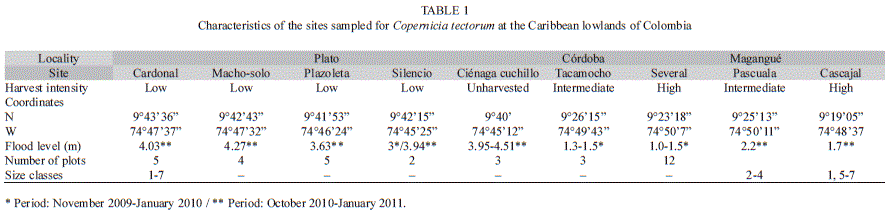
Sampling: Data were taken at three localities: Plato (Magdalena) and Magangué and Córdoba (Bolívar). These are the areas where harvest is most intensive and handicraft production takes place. The three sampling localities differed in the nature of the palm stands, in the level of flooding, and in harvest intensity (Table 1). We chose sites with a history of use of at least 20 years, in order to be able to detect the effect of harvest on the palm populations. Sites were identified with the help of land owners and harvesters. Five sites were chosen at Plato, where the largest palm populations occur (area >1ha), mostly on common lands. Four of these sites are harvested (Cardonal, Machosolo, Plazoleta, and Silencio); at the fifth site (Ciénaga Cuchillo) there was no records of spear leaf harvest, although expanded leaves were occasionally harvested for thatching. At Magangué and Córdoba, palm stands are small (<1ha), scattered, and they are all found on private property. Flood level at all sites was measured on basis of the marks left by water on the palm stems or on neighboring trees.
Field work was carried out in February-March 2010 and April 2011. We established 34 plots of 20×10m, 19 of them in Plato, 12 in Magangué and three in Córdoba. Plot number was proportional to the area of the palm stands. At each plot we counted and tagged all individuals, and recorded the following variables for each individual of C. tectorum: stem height (up to the petiole base of the oldest live leaf); diameter at breast height (1.3m) of naked stems; number of leaves; and number and developmental stage of reproductive structures. Additionally, for juveniles lacking an aerial stem we recorded the length and number of segments of the youngest expanded leaf; for seedlings we recorded lamina length and number of primary veins of the youngest expanded leaf (Galeano et al., 2010). We studied leaf production by marking with colored wire the youngest leaf of 164 palms of different size classes at three sites in Plato and Magangué (Table 1). Thirteen months later we counted new leaves produced, and the increase in number of primary veins or segments from one leaf to the next one in 87 palms that survived. We used the mortality rate of these marked palms to complement data obtained from two 10x10 plots, one at Plato and one at Magangué.
Abundance was estimated using Google Earth images and aerial photographs of the Instituto Geográfico Agustín Codazzi (flight IGAC 2719 of 2004), in which palm crowns of adults and subadults can be recognized. The area of the palm stands at Plato was calculated on the images, and abundance was estimated through a direct count of crowns on the images, combined with plot data. This estimate did not include isolated palms or areas smaller than 1ha, nor did it include the palm stands of Magangué and Córdoba, where the smaller size of the stands and of the palms did not make them clearly recognizable on the images.
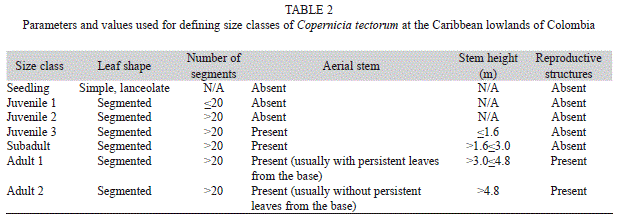
For data analysis, we divided the population into seven size classes, based on developmental traits, like leaf division, occurrence of an aerial stem, and reproduction (Table 2) (Galeano et al., 2010). When plotting segment number vs. segment length, we found a change in the slope of the curve at 20 segments (data not shown); because of this, we took >20 segments as the lower limit of the juvenile 2 class. The presence of an aerial stem marks the division of juveniles 2 and juveniles 3, but some stemmed palms have leaves similar to those of juvenile 2; because of this, we chose as the upper limit of the juvenile 2 size class the average number between the highest number of segments of a stemless palm and the lowest number of segments of a stemmed palm. We then used that figure to classify any particular individual as either juvenile 2 or juvenile 3. We chose a height of 1.6m as the upper limit of the juvenile 3 category, as this was the lowest height recorded for a reproducing individual. As palms age, they shed the persistent leaves from the base. Based on this, we defined the limit of adults 1 and adults 2 at 4.8m, the average height of the stem’s naked portion.
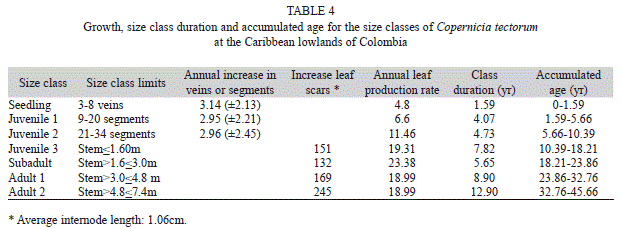
Duration of the seedling and juvenile1 and juvenile 2 age classes was estimated by dividing the range in number of veins (seedlings) or of leaf segments (juveniles) by the annual rate of increase in veins or segments, respectively. For juveniles 3, subadults, and adults, we combined stem length, internode length, and leaf production rate. Leaf scars are close to each other (Fig. 2), and most stems have damages at several places, thus measuring the length of individual internodes throughout the stem is not feasible on all individuals; therefore, we obtained average internode length by dividing the length of a stem portion 0.5-1m long by the number of scars in that portion in stems where scars could be clearly identified. Measurements were taken in a sample of ten individuals of various sizes and at different sites. The total age of a stemmed individual was calculated as the total number of internodes estimated for its size, divided by the leaf production rate for the corresponding size, and adding the estimated age for the stemless phase (Corner, 1966; Pinard, & Putz, 1992; Galeano et al., 2010). As leaf production rate did not differ between Magangué and Plato, data were therefore pooled. Similarly, leaf production rates of adults 1 and adults 2 were also pooled, as the sample of adults 2 (four palms) was too small.
We compared population structure among the three localities Magangué, Córdoba and Plato and among sites at Plato using Kolgomorov-Smirnov (KS) test (Sokal, & Rohlf, 1995). Palm density and leaf production rate were analyzed with the non-parametric Kruskal-Wallis test. When significant differences were found, we used the Mann-Whitney test to explore which groups were different. All analyses were made in Statgraphics Plus and SPSS.
Results
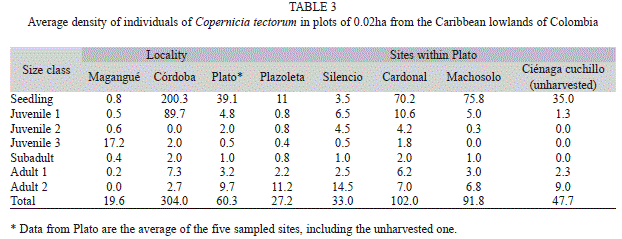
The area of palm stands at Plato was estimated at 480 ha, located between the Magdalena river and the Zárate lagoon. The number of adults and subadults was estimated as ca. 273 000. Overall stand density ranged between 9 and 391 individuals per plot, i.e., 450-19 550 individuals per ha. Adult density was 300-1 000 individuals per ha at Plato, 0-100 individuals per ha at Magangué, and 250-950 individuals per ha at Córdoba. In general, density was significantly different at the three localities (KV=15.3748, p=0.0005), but not within the sites at Plato, including the unharvested site (KV=5.4476, p=0.2444) (Table 3).
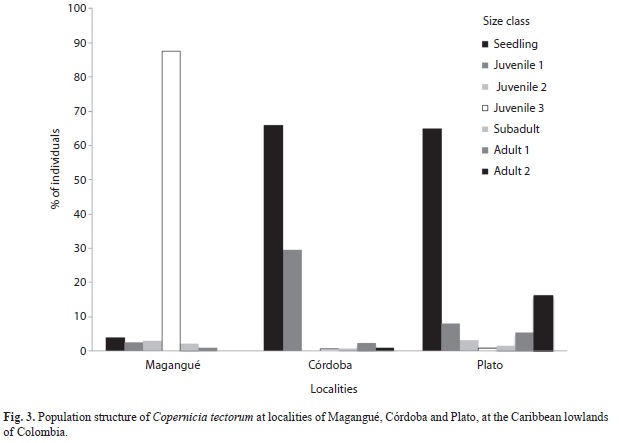
The population at Magangué differed from those at Plato and Córdoba in seedlings, juveniles 3, and adults 1; it also differed from populations at Plato in terms of adults 2. Plato and Córdoba differed only in adults 2 (Fig. 3). At Magangué most palms (88%) were juveniles 3, whereas reproductive individuals (1%) and seedlings (4%) were scarce (Fig. 4A). At Plato and Córdoba there were high proportions of seedlings (65% and 66%, respectively), a decreasing amount of juveniles 1 (8% and 29%, respectively), but few juveniles 2, juveniles 3, and subadults (5.7% and 1.3%, respectively, when pooled). Plato, in turn, had a remarkable proportion of adults 2 (16%) (Fig. 4B), while this class was scarcely represented in Córdoba (1%) and did not occur in Magangué.
Sites within Plato, including the unharvested one, did not differ among them, except for a difference in adults 2 between Silencio y Machosolo. Only the sites Plazoleta, Cardonal and Silencio had individuals in all size classes. The unharvested population at Ciénaga Cuchillo lacked juveniles 2, juveniles 3, and subadults, and it had scarce juveniles 1 (3%) (Fig. 5).
There was a steady increase in leaf production rate with age, reaching a maximum of 23.4 leaves per year in subadults, and then decreasing to 19 leaves per year in adults (Table 4). It took adult palms about one year to replace their whole crown. There was no strong correlation between initial leaf number and leaf production rate either at Plato or Magangué (r=0.459 at Plato and 0.099 at Magangué). Palms start to reproduce when they are ca. 23.9 years old; the age of the tallest palm measured (7.4m) was estimated as 46 years. Mortality at Plato was highest for seedlings (57%), and decreased steadily for juveniles (juveniles 1: 43%; juveniles 2: 40%; juveniles 3: 35%). Mortality in Magangué was 2.3% for juveniles 2 and 3.35% for juveniles 3.
Discussion
The similar demographic structure of C. tectorum at Plato and Córdoba, both at harvested and unharvested sites (biased towards old individuals, with abundant seedlings and scarce recruitment into the juvenile 1 and juvenile 2 classes), suggests that spear leaf harvest is not affecting the populations. This is probably due to the fact that at this site leaves are harvested only from size class subadults and adults, with a frequency that allows the palms to recover (Torres, 2013; Torres et al., in prep.). At Magangué, on the other hand, juveniles 3 predominate and there are scarce seedlings and adults. This appears to be a result of leaf harvest, as the scarcity of adults forces harvesters to cut spear leaves from individuals as young as juveniles 3, which delays their growth and renders them stunted.
Leaf production rate (19-23 leaves/yr in subadults and adults) appears to be the fastest one recorded so far for any palm (cf. Henderson, 2002; Bernal & Galeano, 2013), comparable only to that of Hyphaene petersiana (Fanshawe, 1967), another coryphoid palm that grows in similar environments in Africa. Such a fast leaf production rate accounts for the fast growth and the short life span. Leaf production does not seem to be affected by flood level, as there were no differences between Plato and Magangué, although they had different flood levels (4m and 1.7-2.2m respectively).
An apparent cause of death in juveniles was the accumulation on their crown of plants brought by the flood, which got trapped by the petiole thorns, sometimes completely covering the crown; additionally, hemiepiphytes often grew among the leaf bases, probably from seeds brought also by the flood. This situation did not appear to affect plants over 3m tall, whose crown emerged above the flood level, or older palms with naked stems.
Fires taking place during the dry season did not appear to be a relevant cause of death; after the long drought of early 2010, at all of our study sites we found palms with all their leaves burned but still producing a new spear leaf.
Population structure at Plato and Córdoba resembles Peters’s (1996) type III size class distribution, with most individuals of the same age or size, which is common in heliophitic and pioneer plants. Several traits in the life history of C. tectorum indicate that it is indeed a pioneer of riverine floodlands, as suggested by Troth (1987) for the populations of the Venezuelan savannas. Not only does it thrive in open areas, where it forms dense, monodominant stands, as other riverine pioneers do (cf. Parolin, Oliveira, Piedade, Wittmann, & Junk 2002), but it also has a short establishment phase of only 10.4 years, and its life span (46 years) is one of the shortest recorded for any iteroparous palm (Henderson, 2002).
As with other pioneer riverine plants, flood dynamics appears to be the most important factor influencing the populations of C. tectorum. First, flood water is known to be the primary dispersal agent of C. tectorum (Troth, 1987). Additionally, flooding probably increases seedling recruitment, as water soaking favors seed germination, as shown by Kitzke (1958). But, on the other hand, longer and higher floods increase mortality of seedlings (Troth, 1987) and juveniles, as found at our study sites, and it can also cause severe damage to adults through the accumulation of debris and vegetation that get trapped by the recurved petiole spines, and the germination of epiphytes and hemiepiphytes among the leaf bases.
The palm stands probably play a significant role in regulating river channel dynamics in this inner delta area, where total sedimentation has averaged 3-4 mm/yr over the past 7 500 years (van der Hammen, 1986; Plazas et al., 1988; Kettner, Restrepo, & Syvitski, 2010). This rate is probably higher at the palm stands, as thicker plant stems and larger leaves are known to be effective at capturing particles by reducing water velocity and by providing a surface for sediment adhesion (Brueske, & Barret, 1994; Pasternak, & Brush, 1998). As a matter of fact, at one of our study sites, sediment deposition at the base of some juvenile Copernicia palms reached 2cm during the period 2010-2011.
As sediments accumulate over decades, inundation in the palm stand will decrease, reducing the favorable conditions described above for palm population growth. Eventually, the palm grove will become non-flooded and seedling recruitment will decrease accordingly, and population dynamics will reverse, leading to the eventual disappearance of the palm stand within several decades, given the palm’s short life span. As sedimentation stops, erosion begins, whereas the described accretion process takes place in neighboring areas, where the palm eventually starts to colonize. As accretion progresses in those areas, the former palm grove on this side would be progressively invaded by water. This kind of sedimentary dynamics, well documented for deltaic areas (Serodes, & Troude, 1984; Defries, 1986), is probably altered when the palm stands are destroyed for clearing the land or for using the stems, or when their structure is altered through overharvest, and herbaceous and shrubby vegetation with thinner stems and smaller leaves, less suited for reducing water velocity and trapping sediments, replaces land cover.
If Copernicia tectorum is indeed a pioneer species, with discontinuous and occasional recruitment as a result of river dynamics, the expected population would consist mostly of individuals belonging to just one or two size classes, while at other sites other size classes would be represented. This is exactly what Troth (1967) found in the Venezuelan savannas, where no population had all size classes. Troth pointed the same pattern of stands with individuals of the same size and irregular representation of size classes in other palm species of harsh environments. These include Washingtonia filifera from oases in California (Vogl, & Mchargue, 1966); Copernicia prunifera from semiarid areas in Northeastern Brazil (Taube, 1952); and Copernicia alba from hyperseasonal savannas in the semiarid Chaco (Puechagut, Politi, Bellis, & Rivera, 2012).
Population structure at all sites in Plato shows that only well established stands, mostly with adult palms, occur at this site. In fact, at the youngest stand found (Ciénaga Cuchillo, in Plato) average height of individuals was 4.2m, which suggests that the last successful recruitment took place ca. 30 years ago. Considering the palm’s short life span, this stand will probably disappear within the next two or three decades.
At the palm stands of Plato and Córdoba, abundance, density and leaf production rate offer favorable conditions for leaf harvest for handicraft manufacture. At Magangué, on the contrary, population structure indicates a limited possibility for use, and thus leaf harvest in this area should be regulated, in order to favor recovery of the palm stands. Comparison of harvested and unharvested stands at Plato shows that harvest does not affect population structure if only subadult and adult palms are involved.
Considering the role of river dynamics on the population fate of Copernicia tectorum, management actions must focus on reestablishing, as much as possible, water flow between the river and the lagoon complexes where the palm grows. This includes cleaning up the natural channels and adjusting some infrastructure works in order to improve water circulation, as well as reverting some agricultural areas around the wetlands to their former protective function. These actions have already been identified by the environmental authority (MMA, 2002) as necessary for the recovery of the wetlands in the Mompox Depression.
The described actions should be combined with the implementation of any necessary practices involving leaf harvest itself. Under this scenario, the appropriate management of the C. tectorum stands not only offers a good opportunity for integrating use and conservation of this particular species, but may also have a tremendous impact on the decades-long deltaic cycles of floodable vs. terra firme areas. A better understanding of the palm’s role in the sedimentation cycles might probably help to minimize any negative effects of river dynamics.
Acknowledgments
We thank Universidad Nacional de Colombia, and the Instituto de Ciencias Naturales for offering the facilities for conducting this research. We also thank all the harvesters and artisans of Plato and Magangué, and Nagyla Garrido and Leonardo Rivera for their support during field work. Financial support came from the PALMS FP-7 Project of the European Union (No. 212631). Support from Patrimonio Natural and USAID (Formulación de planes de manejo y uso sostenible de palmas promisorias de la región Caribe que aporte a una estrategia de uso de recursos naturales del bosque seco tropical) was vital for writing the manuscript.
References
Bernal, R., & Galeano, G. (Eds). (2013). Cosechar sin destruir: Aprovechamiento sostenible de palmas colombianas. Bogotá, Colombia: Universidad Nacional de Colombia. [ Links ]
Brueske, C. C., & Barrett, G. W. (1994). Effects of vegetation and hydrologic load on sedimentation patterns in experimental wetland ecosystems. Ecological Engineering, 3, 429-447. [ Links ]
Corner, E. J. H. (1966). The natural history of palms. London: Weidenfeld and Nicolson. [ Links ]
Defries, R. S. (1986). Effects of land-use history on sedimentation in the Potomac estuary Maryland: A water-quality study of the tidal Potomac River and estuary. Washington: United States Geological Survey, Water-Supply Paper 2234-K, United States Government Printing Office. [ Links ]
Fanshawe, D. (1967). The vegetable ivory palm Hyphaene ventricosa Kirk its ecology, silviculture and utilization. Kirkia, 6,105-117. [ Links ]
Galeano, G., & Bernal, R. (2010). Palmas de Colombia: Guía de Campo. Bogotá, Colombia: Editorial Universidad Nacional de Colombia. [ Links ]
Galeano, G., Bernal, R., Isaza, C., Navarro, J., García, N., Vallejo, M. I., & Torres, C. (2010). Evaluación de la sostenibilidad del manejo de palmas. Ecología en Bolivia, 45(3), 85-101. [ Links ]
Henderson, A. (2002). Evolution and ecology of palms. New York, NY: The New York Botanical Garden Press. [ Links ]
Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia. (2011). Informes hidrológicos diarios. Available at http://www.pronosticosyalertas.gov.co/jsp/loader.jsf?lServicio=Publicaciones & lTipo=publicaciones & lFuncion=loadContenidoPublicacion & id=751. [ Links ]
Instituto Geográfico Agustín Codazzi. (1977). Zonas de vida o formaciones vegetales de Colombia. Bogotá, Colombia. [ Links ]
Kettner, A. J., Restrepo, J. D., & Syvitski, J. P. M. (2010). Spatial simulation experiment to replicate fluvial sediment fluxes within the Magdalena River basin, Colombia. Journal of Geology, 118(4), 363-379. [ Links ]
Kitzke, E. D. (1958). A method for germinating Copernicia palm seeds. Principes, 2, 5-8. [ Links ]
Linares, E., Galeano, G., García, N., & Figueroa, Y. (2008). Fibras vegetales empleadas en artesanías en Colombia. Bogotá, Colombia: Artesanías de Colombia S.A., Universidad Nacional de Colombia. [ Links ]
Ministerio del Medio Ambiente. (2002). Plan de Manejo Integral de los Humedales, Subregión de la Depresión Momposina y Cuenca del Rio Sinú. Magangué, Colombia. [ Links ]
Parolin, P., Oliveira, A. C., Piedade, M. T. F., Wittmann, F., & Junk, W. J. (2002). Pioneer trees in Amazonian floodplains: Three key species form monospecific stands indifferent habitats. Folia Geobotanica, 37, 225-238. [ Links ]
Pasternak, G. B., & Brush, G. S. (1998). Sedimentation cycles in a river-mouth tidal freshwater marsh. Estuaries, 21, 407-415. [ Links ]
Peters, C. (1996). The ecology and management of non-timber forest resources. Washington, D.C.: The World Bank. [ Links ]
Pinard, M. A., & Putz, F. E. (1992). Population matrix models and palm resource management. Bulletin de I’Institut Franais d’Etudes Andines, 21, 637-649. [ Links ]
Plazas, C., Falchetti, A. M., van der Hammen, T., & Botero, P. (1988). Cambios ambientales y desarrollo cultural en el bajo río San Jorge. Boletín del Museo del Oro, 20, 58-59. [ Links ]
Puechagut, P., Politi, N., Bellis, L., & Rivera, L. (2012). A disappearing oasis in the semi-arid chaco: deficient palm regeneration and establishment. Journal for Nature Conservation, 21(1), 31-36. [ Links ]
Serodes, J. B., & Troude, J. P. (1984). Sedimentation cycle of a freshwater tidal flat in the St. Lawrence estuary. Estuaries, 7, 119-127. [ Links ]
Sokal, R. & Rohlf, F. (1995). Biometry: the principles and practice of statistics in biological research. 3rd edition. New York, NY: W. H. Freeman and Co. [ Links ]
Taube, E. (1952). Carnauba wax - product of a Braziliam palm. Economic Botany, 6, 379-401. [ Links ]
Ticktin, T. (2004). Review: the ecological implications of harvesting non-timber forest products. Journal of Applied Ecology, 41, 11-21. [ Links ]
Torres, M. C. (2013). Sará (Copernicia tectorum). In R. Bernal, & G. Galeano (Eds.), Cosechar sin destruir: Aprovechamiento sostenible de palmas colombianas (pp. 190-199). Bogotá, Colombia: Editorial Universidad Nacional de Colombia. [ Links ]
Troth, R. (1987). Ecology of woody plant communities in flooded savannas (Llanos) of central Venezuela, and the role of Copernicia tectorum (Palmae). (Ph. D. Dissertation). University of Michigan. [ Links ]
van der Hammen, T. (1986). Fluctuaciones holocénicas del nivel de inundaciones en la cuenca del Bajo Magdalena-Cauca-San Jorge (Colombia). Geología Norandina, 10, 11-18. [ Links ]
Vogl, R. J., & Mchargue, L. T. (1966). Vegetation of California Fan Palm Oases on the San Andreas Fault. Ecology, 47(4), 532-540. [ Links ]
Wong, J., Thornber, K., & Baker, N. (2001). Evaluación de los recursos de productos forestales no madereros: Experiencia y principios biométricos. (Productos forestales no madereros 13). Roma, Italia: Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO). [ Links ]
Brueske, C. C., & Barrett, G. W. (1994). Effects of vegetation and hydrologic load on sedimentation patterns in experimental wetland ecosystems. Ecological Engineering, 3, 429-447. [ Links ]
Corner, E. J. H. (1966). The natural history of palms. London: Weidenfeld and Nicolson. [ Links ]
Defries, R. S. (1986). Effects of land-use history on sedimentation in the Potomac estuary Maryland: A water-quality study of the tidal Potomac River and estuary. Washington: United States Geological Survey, Water-Supply Paper 2234-K, United States Government Printing Office. [ Links ]
Fanshawe, D. (1967). The vegetable ivory palm Hyphaene ventricosa Kirk its ecology, silviculture and utilization. Kirkia, 6,105-117. [ Links ]
Galeano, G., & Bernal, R. (2010). Palmas de Colombia: Guía de Campo. Bogotá, Colombia: Editorial Universidad Nacional de Colombia. [ Links ]
Galeano, G., Bernal, R., Isaza, C., Navarro, J., García, N., Vallejo, M. I., & Torres, C. (2010). Evaluación de la sostenibilidad del manejo de palmas. Ecología en Bolivia, 45(3), 85-101. [ Links ]
Henderson, A. (2002). Evolution and ecology of palms. New York, NY: The New York Botanical Garden Press. [ Links ]
Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia. (2011). Informes hidrológicos diarios. Available at http://www.pronosticosyalertas.gov.co/jsp/loader.jsf?lServicio=Publicaciones & lTipo=publicaciones & lFuncion=loadContenidoPublicacion & id=751. [ Links ]
Instituto Geográfico Agustín Codazzi. (1977). Zonas de vida o formaciones vegetales de Colombia. Bogotá, Colombia. [ Links ]
Kettner, A. J., Restrepo, J. D., & Syvitski, J. P. M. (2010). Spatial simulation experiment to replicate fluvial sediment fluxes within the Magdalena River basin, Colombia. Journal of Geology, 118(4), 363-379. [ Links ]
Kitzke, E. D. (1958). A method for germinating Copernicia palm seeds. Principes, 2, 5-8. [ Links ]
Linares, E., Galeano, G., García, N., & Figueroa, Y. (2008). Fibras vegetales empleadas en artesanías en Colombia. Bogotá, Colombia: Artesanías de Colombia S.A., Universidad Nacional de Colombia. [ Links ]
Ministerio del Medio Ambiente. (2002). Plan de Manejo Integral de los Humedales, Subregión de la Depresión Momposina y Cuenca del Rio Sinú. Magangué, Colombia. [ Links ]
Parolin, P., Oliveira, A. C., Piedade, M. T. F., Wittmann, F., & Junk, W. J. (2002). Pioneer trees in Amazonian floodplains: Three key species form monospecific stands indifferent habitats. Folia Geobotanica, 37, 225-238. [ Links ]
Pasternak, G. B., & Brush, G. S. (1998). Sedimentation cycles in a river-mouth tidal freshwater marsh. Estuaries, 21, 407-415. [ Links ]
Peters, C. (1996). The ecology and management of non-timber forest resources. Washington, D.C.: The World Bank. [ Links ]
Pinard, M. A., & Putz, F. E. (1992). Population matrix models and palm resource management. Bulletin de I’Institut Franais d’Etudes Andines, 21, 637-649. [ Links ]
Plazas, C., Falchetti, A. M., van der Hammen, T., & Botero, P. (1988). Cambios ambientales y desarrollo cultural en el bajo río San Jorge. Boletín del Museo del Oro, 20, 58-59. [ Links ]
Puechagut, P., Politi, N., Bellis, L., & Rivera, L. (2012). A disappearing oasis in the semi-arid chaco: deficient palm regeneration and establishment. Journal for Nature Conservation, 21(1), 31-36. [ Links ]
Serodes, J. B., & Troude, J. P. (1984). Sedimentation cycle of a freshwater tidal flat in the St. Lawrence estuary. Estuaries, 7, 119-127. [ Links ]
Sokal, R. & Rohlf, F. (1995). Biometry: the principles and practice of statistics in biological research. 3rd edition. New York, NY: W. H. Freeman and Co. [ Links ]
Taube, E. (1952). Carnauba wax - product of a Braziliam palm. Economic Botany, 6, 379-401. [ Links ]
Ticktin, T. (2004). Review: the ecological implications of harvesting non-timber forest products. Journal of Applied Ecology, 41, 11-21. [ Links ]
Torres, M. C. (2013). Sará (Copernicia tectorum). In R. Bernal, & G. Galeano (Eds.), Cosechar sin destruir: Aprovechamiento sostenible de palmas colombianas (pp. 190-199). Bogotá, Colombia: Editorial Universidad Nacional de Colombia. [ Links ]
Troth, R. (1987). Ecology of woody plant communities in flooded savannas (Llanos) of central Venezuela, and the role of Copernicia tectorum (Palmae). (Ph. D. Dissertation). University of Michigan. [ Links ]
van der Hammen, T. (1986). Fluctuaciones holocénicas del nivel de inundaciones en la cuenca del Bajo Magdalena-Cauca-San Jorge (Colombia). Geología Norandina, 10, 11-18. [ Links ]
Vogl, R. J., & Mchargue, L. T. (1966). Vegetation of California Fan Palm Oases on the San Andreas Fault. Ecology, 47(4), 532-540. [ Links ]
Wong, J., Thornber, K., & Baker, N. (2001). Evaluación de los recursos de productos forestales no madereros: Experiencia y principios biométricos. (Productos forestales no madereros 13). Roma, Italia: Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO). [ Links ]
1. Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Apartado 7495, Bogotá, Colombia; mctorresrom@unal.edu.co, gagaleanog@unal.edu.co, rgbernalg@gmail.com
Received 12-VI-2014. Corrected 10-XII-2014. Accepted 19-I-2015.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}