










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.63 n.2 San José Apr./Jun. 2015
Population structure of the red mangrove crab, Goniopsis cruentata (Decapoda: Grapsidae) under different fishery impacts: Implications for resource management
Estructura de la población del cangrejo rojo de manglar, Goniopsis cruentata (Decapoda: Grapsidae) bajo diferentes impactos de pesca: Implicaciones para la gestión de los recursos
Estructura de la población del cangrejo rojo de manglar, Goniopsis cruentata (Decapoda: Grapsidae) bajo diferentes impactos de pesca: Implicaciones para la gestión de los recursos
Gustavo L. Hirose1*,2*, Laize S. Souza1, Sonja L.R. Silva1, Douglas F.R. Alves1,2 & Maria Lucia Negreiros-Fransozo2,3*
Abstract
The red mangrove crab, Goniopsis cruentata, influences the recruitment and composition of plant species in the mangrove ecosystem and it is an important fishery resource. Nevertheless, no current management and conservation plans are available for this species for the Brazilian coast. This investigation evaluated the population structure and reproductive biology in populations of G. cruentata under contrasting fishery pressures. The sampling program was carried out in two mangroves, Vaza-Barris and Sergipe River, from January through December 2011. Crabs from both mangroves were randomly collected by a professional fisherman during daytime low tide periods, using a fishing rod baited with pieces of a locally abundant gastropod, Pugilina morio, during 20min/area (catch per unit effort). Monthly measurements of air, sediment surface layer and water temperatures were obtained with a digital thermometer and salinity with an optical refractometer. Both crab populations were compared concerning their abundance, body size, sex ratio, size at onset of sexual maturity and fecundity (FI). Abiotic factors (air, water and mud temperature; and salinity) showed no significant differences between sampling localities. A total of 4 370 crabs were sampled, 2 829 from the Sergipe River and 1 541 from the Vaza-Barris River. The abundance and body size of crabs were compared between mangroves, and statistically significant differences were found. The sex ratio for both populations differed from the expected 1:1 ratio, and a significant deviation in favor of juvenile males was obtained, while adults showed a bias toward females. The estimated size at onset of sexual maturity for both sexes was similar in both populations. However, the populations differed significantly in the number and volume of eggs: a higher FI was obtained in females from the Sergipe River, while a higher egg volume was observed in females from the Vaza-Barris River mangrove. These results indicated a tendency to decrease the body size, the abundance of crabs and the reproductive potential of the species, with higher fishing intensities, reinforcing the need to develop a management plan as well as to establish conservation units for G. cruentata in the Northeastern Brazilian coast. Habitat loss in conjunction with long-term overfishing can have irreversible consequences, which can impact not only the populations of commercially exploited crabs, but the dynamics of virtually the entire mangrove ecosystem.
Key words: body size, fecundity, Grapsidae, management plan, overfishing.
Resumen
El cangrejo rojo Goniopsis cruentata influye en el reclutamiento y la composición de especies vegetales del ecosistema manglar y es un importante recurso pesquero. Sin embargo hasta la fecha, no se han desarrollado planes de manejo y conservación para esta especie en Brasil. La presente investigación evaluó la estructura poblacional y la biología reproductiva de las poblaciones de G. cruentata expuestas a diferentes presiones de pesca. El muestreo se realizó en dos manglares, Río Sergipe y Río Vaza-Barris entre enero y diciembre 2011. En ambos manglares, los animales fueron recolectados de día durante la marea baja por un pescador profesional, utilizando una caña de pescar con carnada del gasterópodo Pugilina morio, durante 20min/área (captura por unidad de esfuerzo, CPUE). Mensualmente con un termómetro digital se registró la temperatura del aire, agua y capa superficial del sedimento, mientras que la salinidad fue cuantificada utilizando un refractómetro óptico. Las poblaciones fueron comparadas en su abundancia, tamaño corporal, proporción de sexos, talla de primera madurez sexual y fecundidad. Los factores abióticos(temperatura del aire, agua y sedimento y la salinidad) no mostraron diferencias significativas entre ambos sitios de muestreo. Se recolectaron un total de 4 370 cangrejos; 2 829 en el río Sergipe y 1 541 en Vaza-Barris. Se registraron diferencias significativas entre los manglares con respecto a la abundancia y el tamaño corporal de los animales. La proporción de sexos de ambas poblaciones difirió de la relación 1:1 esperada y se obtuvo una desviación significativa en favor de machos jóvenes, mientras que los adultos mostraron una tendencia hacia las hembras. A su vez, las poblaciones difirieron significativamente en el número de huevos, con un mayor IF para las hembras del manglar del río Sergipe. El volumen de los huevos también mostró diferencias significativas entre ambas poblaciones. Estos resultados indican que la alta intensidad de pesca podría estar afectando dichos parámetros, lo que refuerza la necesidad de desarrollar un plan de manejo y la creación de unidades de conservación para G. cruentata en la costa noreste de Brasil. La pérdida del hábitat junto con la sobrepesca a largo plazo, pueden tener consecuencias irreversibles, que afectan no sólo a las poblaciones de cangrejos de importancia comercial, sino también la dinámica del ecosistema de manglar.
Palabras clave: fecundidad, Grapsidae, plan de manejo, sobrepesca, tamaño corporal.
Brazil is the largest South American country, with 7 408km of coastline of which 6 786km contains mangrove forests with an estimated area of 25 000km2 (Saenger, Hegerl, & Davie, 1983; Schaeffer-Novelli, Cintrón-Molero, Soares, & De-Rosa, 2000). The mangrove vegetation contributes to habitat complexity and the diversity of the mangrove ecosystem (Macintosh, Ashton, & Havanon, 2002; Colpo, Chacur, Guimarães, & Negreiros-Fransozo, 2011), and is considered the habitat with the richest diversity of land-dwelling crabs (Hartnoll, & Gould, 1988).
Crabs of the families Ocypodidae, Ucididae, Sesarmidae and Grapsidae are among the most abundant and ecologically significant animals found in mangrove ecosystems, where they play a key role in food webs and energy flow (Macintosh, 1988; Schubart, Cuesta, & Felder, 2002; Kristensen, 2008). Ocypodid and Ucidid crabs cause bioturbation of the sediment and have the potential to influence and regulate mangrove production (Nordhaus, Wolff, & Diele, 2006; Kristensen, 2008; Smith, Wilcox, & Lessmann, 2009).
Grapsid crabs consume and process large quantities of mangrove leaf litter (Lee, 1998), often acting as the first consumers processing the mangrove carbon, even before microbial colonization (Nerot, Meziane, Provost-Govrich, & Rybarczyk, 2009). By facilitating microbial colonization and degradation of organic matter, grapsid crabs can also provide food for other organisms in mangrove areas (Lee, 1997; Nerot et al., 2009). Goniopsis cruentata is one of the most abundant grapsid crabs of mangrove ecosystems in the Brazilian South Atlantic, and the trophic role of this crab in the ecosystem is wider than that observed for sesarmid and ocypodid mangrove crabs (Lima-Gomes, Cobo, & Fransozo, 2011). Recently, a study by Ferreira, Ganade, Freire and Attayde (2013) showed that G. cruentata plays an important role in propagule predation, and may affect the mangrove recruitment and community composition.
Goniopsis cruentata (Latreille, 1803) is restricted to the Western Atlantic, and in Brazil occurs from Pará to Santa Catarina (Melo, 1996). During low tide this species can be found walking on the substrate, hiding in burrows made by other species and crevices, or climbing the trunks and roots of trees, occupying virtually all mangrove microhabitats (Cobo, & Fransozo, 2003). The species is consumed and exploited by artisanal fisherman, mainly in Northeastern Brazil (Moura, & Coelho, 2004; Maciel, & Alves, 2009).
In recent years, fishing pressure on G. cruentata has been increasing, since the stocks of Ucides cordatus (Linnaeus, 1763) have been significantly reduced, mainly due to habitat destruction, Lethargic Crab Disease (LCD) and overfishing (Botelho, Andrade, & Santos, 2004; Diele, Koch, & Saint-Paul, 2005; Guerra et al., 2013). Some researchers have also reported an apparent decline in stocks of G. cruentata under different impacts, such as, cutting trees and discharge of solid waste (Menezes, Araújo, & Calado, 2012), as well as decreased body size of these crabs (Maciel, & Alves, 2009). This is a matter for concern, as no management plan has yet been developed to ensure the conservation of natural stocks of this species in Brazil. The majority of studies have focused on the exploitation of resources in aquatic ecosystems (Pauly et al., 2002; Costa, Fransozo, Freire, & Castilho, 2007; Fisher, Frank, & Leggett, 2010; Almeida, Baeza, Fransozo, Castilho, & Fransozo, 2012; Baeza et al., 2013), compared with the exploitation of semi-terrestrial crabs living in the inter- and supratidal zones (Diele, Koch, & Saint-Paul, 2005).
In the last decade, fishery monitoring data for G. cruentata, provided by the Brazilian Institute of Environment and Renewable Natural Resources (IBAMA), point to an estimated catch for the State of Sergipe (Brazil) of about 0.5tons/year. However, recent fishery statistics for the Sergipe coast showed different results, estimating the harvest for the year 2010 alone at 115 tons (Souza, Dantas-Junior, Silva, Félix, & Santos, 2012). This estimate exceeds the average annual production for all of Northeastern Brazil, estimated by IBAMA at about 71.87±38.25tons/year. This is alarming because underestimation of fishing of G. cruentata on the Northeast coast of Brazil may be hiding the actual situation of overfishing of stocks of this species.
Taking into account the ecological and socioeconomic importance of G. cruentata on the Brazilian coast and the lack of a plan for management and conservation by government agencies, the present investigation evaluated the population structure and reproductive biology in two populations of G. cruentata under different fishery pressures. The data were compared with results obtained by different researchers in recent years, in order to assess possible spatial and temporal changes in population features.
Materials and Methods
Study site: The samples of G. cruentata were obtained monthly from January through December 2011. The sampling program was conducted in two mangroves, Sergipe (10º48’52” S - 37º06’10.4” W) and Vaza-Barris (11º06’26.6” S - 37º13’53.7” W), the two major mangrove ecosystems in the state of Sergipe, with estimated areas of 54.96km2 and 59.96km2, respectively, on the Vaza-Barris and Sergipe rivers (Carvalho, & Fontes, 2007). The mangroves in this part of the Brazilian coast develop in protected estuaries because of the high wave energy on the open coast. The mangroves are composed mainly of members of the genera Rhizophora, Laguncularia and Avicennia, with stands reaching more than 10m in height (Schaeffer-Novelli, Cintrón-Molero, & Adaime, 1990). In this study, Rhizophora mangle L. dominated the two study areas, and the substrate was typically muddy (personal observation).
The catch of G. cruentata in the Sergipe River mangrove during 2010 was estimated at 187.12kg/year, and for the Vaza-Barris River mangrove at 17 140.27kg/year (Souza et al., 2012). Unfortunately, long-term monitoring data for the species in these areas are nonexistent. The distance between the sampling localities is approximately 25km; the great difference in the crab catch between these mangroves is mainly related to the fishing intensity from riverside communities, which have distinctive cultural traits. The main target of artisanal fishing on the Sergipe River is U. cordatus, while along the Vaza-Barris River the fishery is focused on bothG. cruentata and U. cordatus.
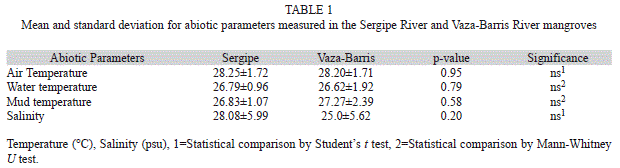
Abiotic factors: The similarity of the localities was analyzed in relation to environmental variables. Monthly measurements of air, sediment surface layer and water temperatures were obtained with a digital thermometer and salinity with an optical refractometer. Unfortunately, data concerning the sediment grain size and organic matter content were not taken. Data on air temperature and salinity were not transformed. Data on water and sediment temperature were log-transformed, but the homogeneity of variances remained heterogeneous. The similarity in air temperature and salinity between the localities was assessed by Student’s t-test (α=0.05). The water and mud temperatures were compared using a non-parametric Mann-Whitney U test (α=0.05).
Sampling methodology: The crabs were collected in the intertidal zone, during daytime low-tide periods (spring tides: new or full moon) in Sergipe and Vaza-Barris mangroves, which are areas typically exploited by the riverine communities. In each mangrove, six equidistant areas of 38.5m2 each were delimited, 20m apart and approximately 10m from the ebb-tide limit. The crabs were randomly captured by a professional fisherman using a fishing rod baited with pieces of a locally abundant gastropod, Pugilina morio, during 20min/area (catch per unit effort). To estimate the size of each area, the maximum range of the line attached to the fishing rod (3.5m) was used as the radius of the area. The total number of crabs caught each month in the six areas was considered a sample unit. This sampling procedure was employed because the use of quadrats or transects for measuring crab abundance is not appropriate in this species. The agility of crabs in moving in this heterogeneous habitat prevents sampling by other methods. This procedure is commonly used by artisanal fishermen to catch G. cruentata in Northeastern Brazil; for details, see Maciel and Alves (2009).
After each collection, the crabs were stored in plastic bags and transported to the laboratory, where they were sexed by observation of the abdomen (narrow for males and wide for females) and number of pleopods (2 pairs for males and 4 pairs for females). Ovigerous females were placed in individual plastic bags in the field, to prevent egg loss. Subsequently, crabs were measured for the following dimensions, with a digital caliper (0.05mm): Carapace Width (CW) and Abdomen Width (AW) for males and females; and Gonopod Length (GL) only for males.
Abundance and body size: The data on crab abundance were normal and homoscedastic. The abundance of G. cruentata was compared among months and localities, as was the interaction between these two factors, using a factorial analysis of variance (two-way ANOVA, months/localities as fixed factors) (Zar, 2010). After differences among months and between localities were found, a post hoc Tukey test (α=0.05) was carried out.
Data for body size were log-transformed, but the homogeneity of variances remained heterogeneous. Even though this violates the assumptions of a parametric test, a two-way ANOVA (α=0.05) was performed to compare the mean size of individuals and sexes. This procedure was adopted because the analysis of variance is robust, operating well even with considerable heterogeneity of variances (Zar, 2010).
Sex ratio: Crabs were separated into juvenile or adult demographic categories by adopting the highest value obtained by the analyses of morphological and gonadal maturity, for each sampling location. The sex ratio was estimated as the quotient between the number of males and the total number of individuals in the sample; sex ratios higher or lower than 0.5 indicated a population skewed toward males or females, respectively. For each locality, deviations from a balanced sex ratio were evaluated for juveniles and adults separately, using a binomial test (Wilson & Hardy, 2002).
Size at Onset Maturity (SOM): In order to estimate the SOM, males were first grouped into age categories according to the relationship between CW and GL. Similarly, females were first grouped into age categories according to the relationship between CW and AW. Morphometric variables used to determine SOM were based on previous reports, to allow comparisons. Next, a K-means non-hierarchical clustering analysis was performed; this clustering analysis distributes the data in groups of numbers previously established by an iterative process that minimizes the variance within groups and maximizes it among them. The result of the classification (K-means) was refined using discriminant analysis. This statistical procedure was based on Sampedro, González-Gurriarán, Freire & Muiño (1999) and Corgos and Freire (2006).
If the CW overlapped between the juvenile and adult age categories, the SOM was determined as the CW at which 50% of the specimens showed morphometric relationships characteristic of the adult condition (W 50%). This value was obtained by logistic function interpolation AP=a/(1+b*exp(-cx)) adjusted to the data for adult proportions (AP) vs. size classes (1.0mm) (CW), where the value of a is first estimated to be the maximal value of y. The values of b and c are then estimated using a straight-line fit to a linearized model.
Gonadal sexual maturity (SOG): Data for the carapace width (CW) were related to the stages of gonadal development in order to determine the SOG of G. cruentata. These data were analyzed separately for males and females by the macroscopic characterization, based on color, shape and volume of the gonads and hepatopancreas in relation to the thoracic cavity. Gonads were classified into four stages of development for both sexes, according to the model proposed by Cobo and Fransozo (2005) and Souza and Silva (2009) for G. cruentata.
Fecundity and egg size: The determination of fecundity was based on the number of eggs spawned per female. Thirty females from each mangrove, in the same breeding season, were selected for fecundity analysis. The females analyzed had eggs in the early stages of embryonic development (Costa, & Negreiros-Fransozo, 1996), to avoid any bias in the analysis caused by the loss of eggs during the incubation period. The egg mass of each ovigerous female was separated from the pleopods under a stereomicroscope. A subsample of 1 000 eggs (from different pleopods) was removed from each female. After the subsample was separated and counted, the eggs (total+subsample) were dried in an oven at a constant temperature of 60°C, and weighed on an analytical balance (0.0001g) every 3h until the weight stabilized.
After drying, the number of eggs of each female was extrapolated from the dry weight of the subsample in relation to the total dry weight of the sample (Pinheiro, & Terceiro, 2000). For comparative purposes, the fecundity index was calculated for each ovigerous female by the formula: FI=n/CW, where FI=Fecundity index, n=number of eggs.
The size of eggs (n=30) was obtained by measuring their diameter, using a stereomicroscope equipped with an imaging and measuring system, and the volume calculated by the ellipsoid formula according to Jones and Simons (1983): v=1/6πI3 where, v=volume of egg and I=diameter of egg. The number and volume of eggs from the two populations were compared using Student’s t test (Zar, 2010).
Results
Abiotic factors: No significant annual differences in abiotic factors between sampling localities were observed. The mean values for each abiotic factor are shown in Table 1.
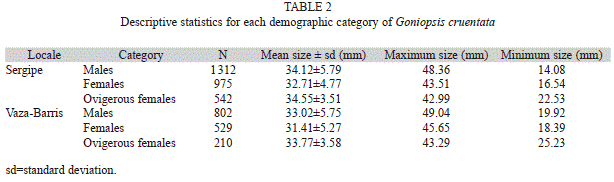
Abundance and body size: During the collection period, a total of 4 370 crabs were sampled, 2 829 from the Sergipe River and 1 541 from the Vaza-Barris River. The mean size of individuals from the Sergipe River was 33.72±5.13mm CL, and from the Vaza-Barris River 32.57±5.41mm CW. The descriptive statistics for each demographic category of the crabs in the two mangroves are shown in Table 2.
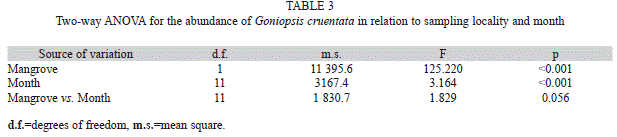
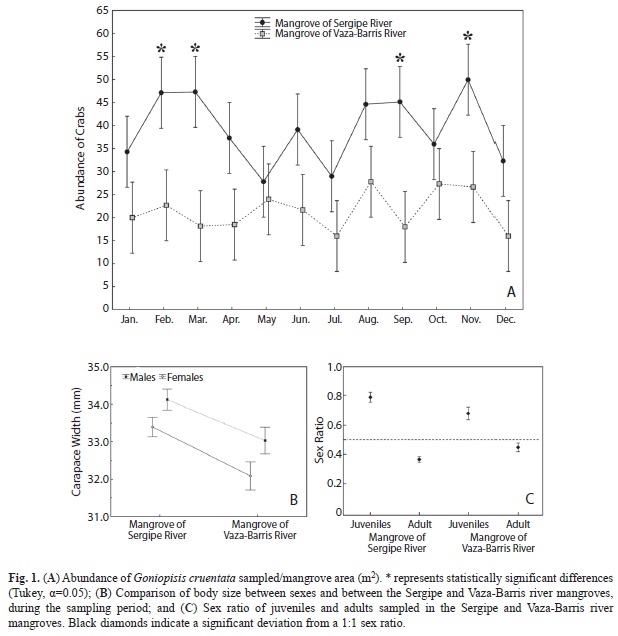
The abundance of crabs was compared between mangroves, and statistically significant differences were found in relation to the mangrove and months (Table 3). Although the number of crabs sampled from the Sergipe River mangrove was higher than from the Vaza-Barris River mangrove, statistically significant differences in the abundance of crabs between the mangroves were found only for February, March, September and November (Fig. 1A).
Regarding the size of the crabs, significant differences were also found between the mangroves and sexes (Table 4). The mean sizes of both sexes were larger in the Sergipe River than in the Vaza-Barris River mangrove. Males were larger than females in both mangroves (Fig. 1B).
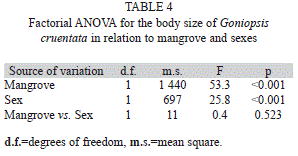
Sex ratio: The sex ratio for both populations differed from the expected 1:1 ratio (male:female) (p<0.01), for both juveniles and adults. A significant deviation in favor of juvenile males was obtained, while adults showed a bias toward females (Fig. 1C).
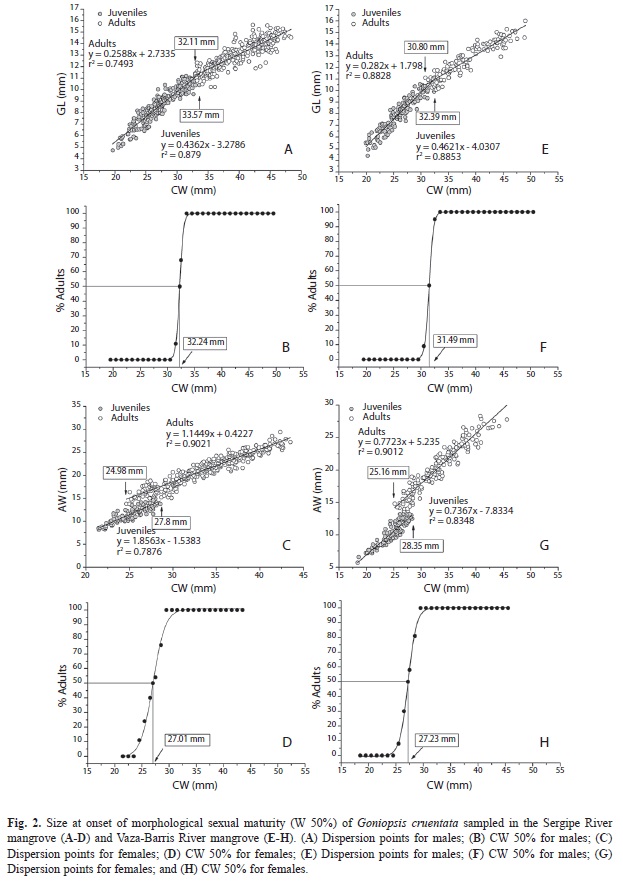
Morphological sexual maturity: The estimated value for morphological sexual maturity was higher for males than for females, for the population of the Sergipe River mangrove. The estimated size where 50% of individuals were already in adult form (W 50%) for males was 32.24mm CW (Fig. 2A, Fig. 2B); the largest juvenile male measured was 33.57mm CW, and the smallest adult male 32.11mm CW. For females, the estimated value was 27.01mm CW (Fig. 2C, Fig. 2D); the largest juvenile female measured was 27.8mm CW and the smallest adult female 24.98mm CW.
The estimated size at onset of sexual maturity for both sexes was similar in the two populations. The estimated size where 50% of individuals were already in adult form (W 50%) in males was 31.49mm CW (Fig. 2E, Fig. 2F); the largest juvenile male measured 32.39mm CW and the smallest adult 30.80mm CW. For females the estimated value was 27.23mm CW (Fig. 2G, Fig. 2H); the largest juvenile female measured 28.35mm CW and the smallest adult 25.16mm CW.
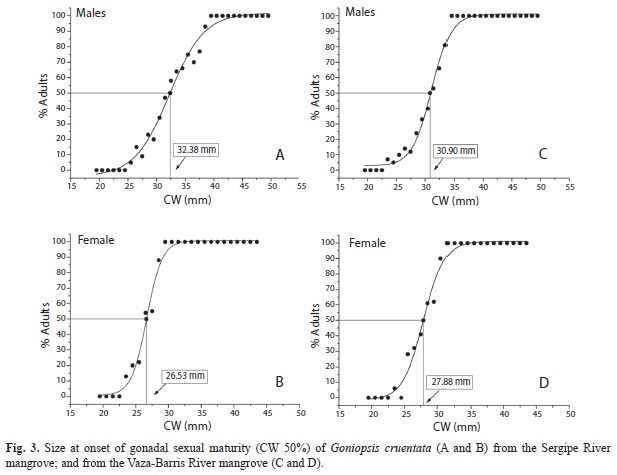
Gonadal sexual maturity: The estimated sizes for gonadal sexual maturity were very close to those estimated from the morphological sexual maturity, 32.38 and 26.53mm CW for males and females, respectively, from the Sergipe River mangrove (Fig. 3A, Fig. 3B) and 30.90 and 27.88mm CW for males and females, respectively, from the Vaza-Barris River mangrove (Fig. 3C, Fig. 3D).
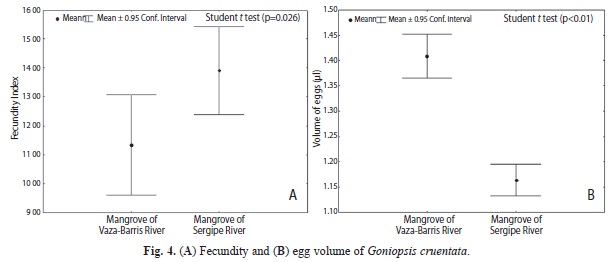
Fecundity and egg size: The populations differed significantly in the number of eggs, with a higher IF for females from the Sergipe River mangrove (t=2.28, p=0.026) (Fig. 4A). The mean number of eggs carried by females from the Sergipe River mangrove was estimated at 50 460±19 828 eggs, while females from the Vaza-Barris River carried 40 005±18 508 eggs. The mean CW for ovigerous females was 35.23±4.6mm for the Sergipe River mangrove; and the largest and smallest ovigerous females measured 42.04 and 25.71mm CW, respectively. The mean CW for ovigerous females in the Vaza-Barris River mangrove was 34.51mm; and the largest and smallest ovigerous females measured 42.08 and 25.62mm CW, respectively.
The egg volume also showed statistically significant differences between populations (t=8.98, p<0.001). In contrast to the trend for fecundity, the population from the Sergipe River carried smaller eggs than the population of the Vaza-Barris River mangrove (Fig. 4B).
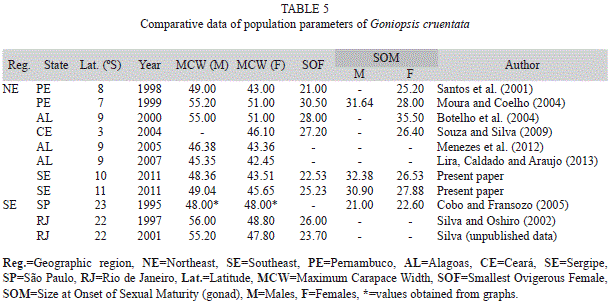
Table 5 compares the data obtained in the Sergipe State with other previously published data.
Discussion
The lack of studies and standardized procedures used in sampling G. cruentata prevents comparisons of its abundance with populations from other mangroves. Nevertheless, considering that this study used the same field procedure for both mangroves, we can infer that the mean crab abundance found in the Vaza-Barris River mangrove is lower than in the Sergipe River mangrove. These differences can be attributed to environmental factors that are intrinsic to a particular mangrove forest or crab population, such as primary productivity, pneumatophore density, or patterns of larval recruitment (Piou, Berger, & Feller, 2009; Sandrini-Neto, & Lana, 2012), but might also be a consequence of the stronger fishing intensity (Tripple, 1995; Carver, Wolcott, Wolcott, & Hines, 2005) in the Vaza-Barris River mangrove.
In Northeastern Brazil, G. cruentata is usually not sold as whole individuals but as processed meat, popularly known as “catado”. Goniopsis cruentata (mean size 33.72±5.13mm CW in individuals from Sergipe State) is not large compared to other mangrove crab species, such as U. cordatus (mean size about 45.00mm CW, on the Northeastern Brazilian coast) (Castiglioni, & Coelho, 2011). Therefore a large number of individuals of G. cruentata are usually required to produce a small amount of meat; according to Carriço et al. (unpublished data), 120-150 adult crabs are needed to produce one kg of “catado”. This makes fishing for G. cruentata a highly impactful activity for the natural populations.
The two populations differed significantly in body size. Body size variations are common, and may reflect the phenotypic plasticity of the organisms or the influence of environmental factors such as photoperiod, temperature, and food availability (Campbell, & Eagles, 1983; Negreiros-Fransozo, Fransozo, González-Gordillo, & Bertini, 2002). One of the most obvious of these changes is related to the latitudinal cline. Intraspecific studies on decapod crustaceans tend to show that larger-bodied crabs occur at higher latitudes (Jones, & Simons, 1983; Hines, 1989; Lardies, & Castilla, 2001; Castilho et al., 2007; Hirose, Fransozo, Tropea, Lopez-Greco, & Negreiros-Fransozo, 2012); this phenomenon is known as Bergmann´s rule (Blackburn, Gaston, & Loder, 1999).
Comparison of the body size of crabs in this and previous studies on the Northeastern Brazilian coast, in relation to space and time, does not show evidence of a latitudinal cline or a spatial dependency in the maximum body size, probably due to intrinsic differences in environmental factors of each mangrove forest. In contrast, the relationship of the body size vs. year shows an apparently temporal pattern, with a tendency to diminution of the maximum body size in relation to time. Body size is probably the most important quantitative trait of an individual. It deeply affects virtually all physiological (e.g., metabolic rate) and fitness traits (e.g., fecundity or mating success), producing strong but not necessarily well-understood allometric relationships within and among organisms (McNab, 1971; Blanckenhorn, & Demont, 2004).
The artisanal fishery for the red mangrove crab targets the larger individuals (personal observation), probably due to the small amounts of meat in juvenile crabs. The result is, in part, that juvenile individuals (less than the estimated W50%) are maintained in the population. Moreover, selection of the largest individuals by fishermen would tend to capture larger numbers of males due to their larger body size. This could explain the deviation in sex ratio recorded for the adults in this study. Furthermore, the collection of larger individuals could favor genotypes with slower growth and a younger age at onset of maturation, among other changes, that reduce the reproductive success of a population (Conover, & Munch, 2002; Carver et al., 2005).
The size at onset of sexual maturity (W 50%) of G. cruentata from Sergipe State was estimated utilizing different procedures (morphological and gonadal maturation). The results were similar, in agreement with Souza and Silva (2009). However, when compared among localities or on a time scale, no apparent pattern was found. This is probably because the SOM is very plastic under different environmental conditions, i.e., different levels of productivity and stress, or the presence, mainly in mangrove areas, of large amounts of growth-inhibiting substances (e.g., polyphenols derived from tannins of halophyte plants) (Power, & Bliss, 1983; Rodríguez, 1987; Díaz, & Conde, 1988 and 1989; Conde, & Díaz, 1989; Negreiros-Fransozo et al., 2002; Hirose et al., 2012).
The existing management plans for commercially exploited crustacean species in Brazil are based on a set of measures designed to maintain a breeding stock large enough for adequate recruitment, and to prevent the capture of individuals in critical phases of their life cycles (MMA/IBAMA, 2011). The size at onset of sexual maturity (W50%) is regularly used to define a minimum size of capture (estimated for Sergipe State in 32.4 and 27.9mm CW for males and females, respectively). In view of the high variability found here (W50%) compared to other locations, an ecosystem-based management plan should be adopted to increase the efficiency of these measures as alternatives for protection of natural populations (Ma, Townsend, Zhang, Sigrist, & Christensen, 2010).
The effect of fishing on the reproductive potential of G. cruentata should be taken into account. Our results show that both sexes are taken by fishermen, and approximately 33% of the females sampled were bearing eggs. According to our observation, ovigerous females are typically used in the production of “catado” in Sergipe, which could heavily impact the reproductive potential of the stock. The estimated mean fecundity of G. cruentata from Sergipe (42 923±19 171 eggs) is lower than that described by Moura and Coelho (2003) from Ceará in 1999 (87 000 eggs). This may be a reflection of the smaller body size in Northeastern populations, which would also tend to reduce the reproductive potential of the species.
Fecundity and egg size differed significantly between the two mangrove areas under different fishing pressure in Sergipe state. These results suggest an apparent trade-off between egg size and egg number. Trade-offs between egg number and size are characteristic of most animals (Timi, Lanfranchi, & Poulin, 2005). According to Fox and Czesak (2000), when environmental conditions vary, the relationship between progeny size and progeny fitness is likely to vary too, resulting in a different optimal progeny size in different environments. Factors that may affect the trade-off between egg size and egg number include climate conditions, predation and population density (Fox, & Czesak, 2000). In the present study, the significant differences in population density between the mangrove localities may affect competition for food and consequently select for egg size (Parker, & Begon, 1986).
Habitat loss in conjunction with long-term overfishing can have irreversible consequences, which can impact not only the populations of commercially exploited crabs, but the dynamics of virtually the entire mangrove ecosystem. Our results indicated a tendency towards a decrease in the body size, abundance and reproductive potential of G. cruentata, probably in response to pressure from artisanal fishing. Our overall evaluation indicates that Northeastern populations of G. cruentata can be currently overfished, reinforcing the need to develop a management plan for the species, as well as for the implementation of conservation units in the mangroves on the Northeastern Brazilian coast.
Acknowledgments
To the CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico) for financial support (CNPq-Universal #472386/2010-7 to the first author; and CNPq – Research Fellowship to MLNF). All sampling in Brazil was carried out according to state and federal laws regulating wildlife collection (SISBIO #24097-1).
References
Almeida, A. C., Baeza, J. A., Fransozo, V., Castilho, A. L., & Fransozo, A. (2012). Reproductive biology and recruitment of Xiphopenaeus kroyeri in a marine protected area in the Western Atlantic: implications for resource management. Aquatic Biology, 17(1), 57-69. [ Links ]
Baeza, J. A., Furlan, M., Almeida, A. C., Barros-Alves, S. P., Alves, D. F. R., & Fransozo, V. (2013). Population dynamics and reproductive traits of the ornamental crab Porcellana sayana: implications for fishery management and aquaculture. Sexuality and Early Development in Aquatic Organisms, 1, 1-12. [ Links ]
Blackburn, T. M., Gaston, K. J., & Loder, N. (1999). Geographic gradients in body size: a clarification of Bergmann’s rule. Diversity and Distributions, 5(4), 165-174. [ Links ]
Blanckenhorn, W. U., & Demont, M. (2004). Bergmann and converse Bergmann latitudinal clines in arthropods: Two ends of a continuum? Integrative and Comparative Biology, 44(6), 413-424. [ Links ]
Botelho, E. R. O., Andrade, C. E. R., & Santos, M. C. F. (2004). Estudo da população de aratu-do-mangue, Goniopsis cruentata (Latreille, 1803) (Crustacea, Decapoda, Grapsidae) no estuário do rio Camaragibe (Alagoas – Brasil). Boletim Técnico Científico do CEPENE, 12, 91-98. [ Links ]
Campbell, A., & Eagles, M. D. (1983). Size at maturity and fecundity of rock crabs, Cancer irroratus, from the Bay of Fundy and southwestern Nova Scotia. Fishery Bulletin, 81(2), 357-362. [ Links ]
Carvalho, M. E., & Fontes, A. L. (2007). A carcinicultura no espaço litorâneo Sergipano. Revista da Fapese, 3(1), 87-112. [ Links ]
Carver, A. M., Wolcott, T. G., Wolcott, D. L., & Hines, A. H. (2005). Unnatural selection: effects of a male-focused size-selective fishery on reproductive potential of a blue crab population. Journal of Experimental Marine Biology and Ecology, 319(1), 29-41. [ Links ]
Castiglioni, D. S., & Coelho, P. A. (2011). Determinação da maturidade sexual de Ucides cordatus (Crustacea, Brachyura, Ucididae) em duas áreas de manguezal do litoral sul de Pernambuco, Brasil. Iheringia Série Zoologia, 101(1-2), 138-144. [ Links ]
Castilho, A. L., Gavio, M. A., Costa, R. C., Boschi, E. E., Bauer, R. T., & Fransozo, A. (2007). Latitudinal variation in population structure and reproductive pattern of the endemic South American shrimp Artemesia longinaris (Decapoda: Penaeoidea). Journal of Crustacean Biology, 27(4), 548-552. [ Links ]
Cobo, V. J., & Fransozo, A. (2003). External factors determining breeding season in the red mangrove crab Goniopsis cruentata (Latreille) (Crustacea, Brachyura, Grapsidae) on the São Paulo State northern coast, Brazil. Revista Brasileira de Zoologia, 20(2), 213-217. [ Links ]
Cobo, V. J., & Fransozo, A. (2005). Physiological maturity and relationship of growth and reproduction in the red mangrove crab Goniopsis cruentata (Latreille) (Brachyura, Grapsidae) on the coast of São Paulo, Brazil. Revista Brasileira de Zoologia, 22(1), 219-223. [ Links ]
Colpo, K. D., Chacur, M. M., Guimarães, F. J., & Negreiros-Fransozo, M. L. (2011). Subtropical Brazilian mangroves as a refuge of crab (Decapoda: Brachyura) diversity. Biodiversity and Conservation, 20(13), 3239-3250. [ Links ]
Conde, J. E., & Díaz, H. (1989). The mangrove tree crab Aratus pisonii in a tropical estuarine coastal lagoon. Estuarine, Coastal and Shelf Science, 28(6), 639-650. [ Links ]
Conover, D. O., & Munch, S. B. (2002). Sustaining fisheries yields over evolutionary time scales. Science, 297(5578), 94-96. [ Links ]
Corgos, A., & Freire, J. (2006). Morphometric and gonad maturity in the spider crab Maja brachydactyla: a comparison of methods for estimating size at maturity in species with determinate growth. Journal of Marine Science, 63(5), 851-859. [ Links ]
Costa, R. C., Fransozo, A., Freire, F. A. M., & Castilho, A. L. (2007). Abundance and ecological distribution of the “sete-barbas” shrimp Xiphopenaeus kroyeri (Heller, 1862) (Decapoda: Penaeoidea) in three bays of the Ubatuba region, southeastern Brazil. Gulf and Caribbean Research, 19, 33-41. [ Links ]
Costa, T. M., & Negreiros-Fransozo, M. L. (1996). Fecundidade de Callinectes danae Smith, 1869 (Crustacea, Decapoda, Portunidae) na região de Ubatuba (SP), Brasil. Arquivos de Biologia e Tecnologia, 39, 393-400. [ Links ]
Díaz, H., & Conde, J. E. (1988). On the food sources for the mangrove tree crab Aratus pisonii (Brachyura: Grapsidae). Biotropica, 20, 348-350. [ Links ]
Díaz, H., & Conde, J. E. (1989). Population dynamics and life history of the mangrove crab Aratus pisonii (Brachyura, Grapsidae) in a marine environment. Bulletin of Marine Science, 45(1), 148-163. [ Links ]
Diele, K., Koch, V., & Saint-Paul, U. 2005. Population structure, catch composition and CPUE of the artisanally harvested mangrove crab Ucides cordatus (Ocypodidae) in the Caeté estuary, North Brazil: indications for overfishing? Aquatic Living Resources, 18(2), 169-178. [ Links ]
Ferreira, A. C., Ganade, G., Freire, F. A. M., & Attayde, J. L. (2013). Propagule predation in a Neotropical mangrove: the role of the Grapsid crab Goniopsis cruentata. Hydrobiologia, 707(1), 135-146. [ Links ]
Fisher, J. A. D, Frank, K. T., & Leggett, W. C. (2010). Global variation in marine fish body size and its role in biodiversity–ecosystem functioning. Marine Ecology Progress Series, 405, 1-13. [ Links ]
Fox, C. W., & Czesak, M. L. 2000. Evolutionary ecology of progeny size in Arthropods. Annual Review of Entomology, 45(1), 341-369. [ Links ]
Guerra, R. S., Nascimento, M. M. F., Miesch, S., Najafzadeh, M. J., Ribeiro, R. O., Ostrensky, A., Hoog, G. S., Vicente, V. A., & Boeger, W. A. (2013). Black yeast biota in the mangrove, in search of the origin of the lethargic crab disease (LCD). Mycopathologia, 175(5-6), 421-430. [ Links ]
Hartnoll, R. G., & Gould, P. (1988). Brachyuran life history strategies and the optimization of egg production. Symposia of the Zoological Society of London, 59, 1-9. [ Links ]
Hines, A. H. (1989). Geographic variation in size at maturity in Brachyuran crabs. Bulletin of Marine Science, 45(2), 356-368. [ Links ]
Hirose, G. L., Fransozo, V., Tropea, C., Lopez-Greco, L. S., & Negreiros-Fransozo, M. L. (2012). Comparison of body size, relative growth and size at onset sexual maturity of Uca uruguayensis (Crustacea: Decapoda: Ocypodidae) from different latitudes in the southwestern Atlantic. Journal of the Marine Biological Association of the United Kingdom, 93(3), 781-788. [ Links ]
Jones, M. B., & Simons, M. J. (1983). Latitudinal variation in reproductive characteristics of a mud crab, Helice crassa (Grapsidae). Bulletin of Marine Science, 33(3), 656-670. [ Links ]
Kristensen, E. (2008). Mangrove crabs as ecosystem engineers; with emphasis on sediment processes. Journal of Sea Research, 59(1), 30-43. [ Links ]
Lardies, M. A., & Castilla, J. C. 2001. Latitudinal variation in the reproductive biology of the commensal crab Pinnaxodes chilenis (Decapoda: Pinnotheridae) along the Chilean coast. Marine Biology, 139(6), 1125-1133. [ Links ]
Lee, S. Y. (1997). Potential trophic importance of the faecal material of the mangrove sesarmine crab Sesarma messa. Marine Ecology Progress Series, 159, 275-284. [ Links ]
Lee, S. Y. (1998). Ecological role of grapsid crabs in mangrove ecosystems: a review. Marine and Freshwater Research, 49, 335-343. [ Links ]
Lima-Gomes, R. C., Cobo, V. J., & Fransozo, A. (2011). Feeding behaviour and ecosystem role of the red mangrove crab Goniopsis cruentata (Latreille, 1803) (Decapoda, Grapsoidea) in a subtropical estuary on the Brazilian coast. Crustaceana, 84, 735-747. [ Links ]
Lira, J. J. P. R., Calado, T. C. S., & Araújo, M. S. L. C. (2013). Breeding period in the mangrove crab Goniopsis cruentata (Decapoda: Grapsidae) in Northeast Brazil. Revista Biología Tropical, 61, 29-38. [ Links ]
Ma, H., Townsend, H., Zhang, X., Sigrist, M., & Christensen, V. (2010). Using a fisheries ecosystem model with a water quality model to explore trophic and habitat impacts on a fisheries stock: A case study of the blue crab population in the Chesapeake Bay. Ecological Modelling, 221, 997-1004. [ Links ]
Maciel, D. C., & Alves, A. G. C. (2009). Conhecimentos e práticas locais relacionadas ao aratu Goniopsis cruentata (Latreille, 1803) em Barra de Sirinhaém, litoral sul de Pernambuco, Brasil. Biota Neotropica, 9, 29-36. [ Links ]
Macintosh, D. J. (1988). The ecology and physiology of decapods of mangrove swamps. Symposia of the Zoological Society of London, 59, 315-341. [ Links ]
Macintosh, D. J., Ashton, E. C., & Havanon, S. (2002). Mangrove rehabilitation and intertidal biodiversity: a study in the Ranong Mangrove Ecosystem, Thailand. Estuarine, Coastal and Shelf Science, 55, 331-345. [ Links ]
McNab, B. K. (1971). On the ecological significance of Bergmann’s rule. Ecology, 52, 845-854. [ Links ]
Melo, G. A. S. (1996). Manual de identificação dos Brachyura (caranguejos e siris) do litoral brasileiro. São Paulo: Plêiade. [ Links ]
Menezes, A. P. D., Araújo, M. S. L. C., & Calado, T. C. S. (2012). Bioecologia de Goniopsis cruentata (Latreille, 1803) (Decapoda, Grapsidae) do complexo estuarino-lagunar Mundaú/Manguaba, Alagoas, Brasil. Natural Resources, 2(2), 37-49. [ Links ]
Moura, N. F. O., & Coelho, P. A. (2003). Fecundidade de Goniopsis cruentata (Latreille, 1803) (Crustacea: Brachyura: Grapsidae) no manguezal do rio Paripe – Pernambuco – Brasil. Tropical Oceanography, 31(2), 127-133. [ Links ]
Moura, N. O. F., & Coelho, P. A. (2004). Maturidade sexual fisiológica em Goniopsis cruentata (Latreille) (Crustácea, Brachyura, Grapsidae) no Estuário de Paripe, Pernambuco, Brasil. Revista Brasileira de Zoologia, 21(4), 1011-1015. [ Links ]
Negreiros-Fransozo, M. L., Fransozo, A., González-Gordillo, J. I., & Bertini, G. (2002). First appraisal on releasing and reinvasion of decapod larvae in a subtropical estuary from Brazil. Acta Limnologica Brasiliensis, 14(2), 87-94. [ Links ]
Nerot, C., Meziane, T., Provost-Govrich, A., Rybarczyk, H., & Lee, S. Y. (2009). Role of grapsid crabs, Parasesarma erythrodactyla, in entry of mangrove leaves into an estuarine food web: a mesocosm study. Marine Biology, 156(11), 2343-2352. [ Links ]
Nordhaus, I., Wolff, M., & Diele, K. (2006). Litter processing and population food intake of the mangrove crab Ucides cordatus in a high intertidal forest in northern Brazil. Estuarine, Coastal and Shelf Science, 67(1), 239-250. [ Links ]
Parker, G. A., & Begon, M. B. (1986). Optimal egg size and clutch size: effects of environment and maternal phenotype. The American Naturalist, 128, 573-592. [ Links ]
Pauly, D., Christensen, V., Guénette, S., Pitcher, T. J., Sumaila, U. R., Walters, C. J., Watson, R., & Zeller, D. (2002). Towards sustainability in world fisheries. Nature, 418(6898), 689-695. [ Links ]
Pinheiro, M. A. A., & Terceiro, O. S. L. (2000). Fecundity and reproductive output of the speckled swimming crab Arenaeus cribrarius (Lamarck, 1818) (Brachyura, Portunidae). Crustaceana, 73(9), 1121-1137. [ Links ]
Piou, C., Berger, U., & Feller, I. C. (2009). Spatial structure of a leaf-removing crab population in a mangrove of North-Brazil. Wetlands Ecology and Management, 17(2), 93-106. [ Links ]
Powers, L. W., & Bliss, D. E. 1983. Terrestrial adaptations. In D. E. Bliss (Ed.), The Biology of Crustacea (pp. 271-333). New York: Academic Press. [ Links ]
Rodríguez, G. (1987). Structure and production in Neotropical mangroves. Tree, 2(9), 264-267. [ Links ]
Saenger, P., Hegerl, E. J., & Davie, J. D. (1983). Global status of mangrove ecosystems. Environmentalist, 3(3), 1-88. [ Links ]
Sampedro, M., González-Gurriarán, E., Freire, J., & Muiño, R. (1999). Morphometry and sexual maturity in the spider crab Maja squinado (Decapoda: Majidae) in Galicia, Spain. Journal of Crustacean Biology, 19, 578-592. [ Links ]
Sandrini-Neto, L., & Lana, P. C. (2012). Distribution patterns of the crab Ucides cordatus (Brachyura, Ucididae) at different spatial scales in subtropical mangroves of Paranaguá Bay (southern Brazil). Helgoland Marine Research, 66(2), 167-174. [ Links ]
Santos, M. C. F., Botelho, E. R. R. O., & Ivo, C. T. C. (2001). Biologia populacional e manejo da pesca de aratu, Goniopsis cruentata (Latreille, 1803) (Crustacea: Decapoda: Grapsidae) no litoral sul de Pernambuco–Brasil. Boletim Técnico-Científico do CEPENE, 9, 87-123. [ Links ]
Schaeffer-Novelli,Y., Cintrón-Molero, G., & Adaime, R. R. (1990). Variability of mangrove ecosystems along the Brazilian Coast. Estuaries, 13(2), 204-218. [ Links ]
Schaeffer-Novelli, Y., Cintrón-Molero, G., Soares, M. L. G., & De-Rosa, T. (2000). Brazilian mangroves. Aquatic Ecosystem Health and Management, 3(4), 561-570. [ Links ]
Schubart, C. D., Cuesta, J. A., & Felder, D. L. (2002). Glyptograpsidae, a new brachyuran family from Central America: larval and adult morphology, and a molecular phylogeny of the Grapsoidea. Journal of Crustacean Biology, 22(1), 28-44. [ Links ]
Silva, Z. S., & Oshiro, L. M. Y. (2002). Aspectos reprodutivos de Goniopsis cruentata (Latreille) (Crustacea, Brachyura, Grapsidae) na Baía de Sepetiba, Rio de Janeiro, Brasil. Revista Brasileira de Zoologia, 19(3), 907-914. [ Links ]
Smith, N. F., Wilcox, C., & Lessmann, J. M. (2009). Fiddler crab burrowing affects growth and production on the white mangrove (Laguncularia racemosa) in a restored Florida coastal marsh. Marine Biology, 156(11), 2255-2266. [ Links ]
Souza, L. P., & Silva, J. R. F. (2009). Morphology of the female reproductive system of the red-clawed mangrove tree crab (Goniopsis cruentata Latreille, 1803). Scientia Marina, 73(3), 527-539. [ Links ]
Souza, M. J. F. T., Dantas-Junior, J. F., Silva, F. C. B., Félix, D. C. F., & Santos, J. C. (2012). Estatística pesqueira da costa do Estado de Sergipe e Extremo norte da Bahia 2010. São Cristóvão: Editora UFS. [ Links ]
Timi, J. T., Lanfranchi, A. L., & Poulin, R. (2005). Is there a trade-off between fecundity and egg volume in the parasitic copepod Lernanthropus cynoscicola? Parasitology Research, 95(1), 1-4. [ Links ]
Wilson, K., & Hardy, I. C. W. (2002). Statistical analysis of sex ratios: an introduction. In I. C. W. Hardy (Ed.), Sex ratios: concepts and research methods (pp. 48-92). Cambridge: Cambridge University Press. [ Links ]
Zar, J. H. (2010). Biostatistical analysis. Upper Saddle River: Prentice-Hall/Pearson. [ Links ]
Baeza, J. A., Furlan, M., Almeida, A. C., Barros-Alves, S. P., Alves, D. F. R., & Fransozo, V. (2013). Population dynamics and reproductive traits of the ornamental crab Porcellana sayana: implications for fishery management and aquaculture. Sexuality and Early Development in Aquatic Organisms, 1, 1-12. [ Links ]
Blackburn, T. M., Gaston, K. J., & Loder, N. (1999). Geographic gradients in body size: a clarification of Bergmann’s rule. Diversity and Distributions, 5(4), 165-174. [ Links ]
Blanckenhorn, W. U., & Demont, M. (2004). Bergmann and converse Bergmann latitudinal clines in arthropods: Two ends of a continuum? Integrative and Comparative Biology, 44(6), 413-424. [ Links ]
Botelho, E. R. O., Andrade, C. E. R., & Santos, M. C. F. (2004). Estudo da população de aratu-do-mangue, Goniopsis cruentata (Latreille, 1803) (Crustacea, Decapoda, Grapsidae) no estuário do rio Camaragibe (Alagoas – Brasil). Boletim Técnico Científico do CEPENE, 12, 91-98. [ Links ]
Campbell, A., & Eagles, M. D. (1983). Size at maturity and fecundity of rock crabs, Cancer irroratus, from the Bay of Fundy and southwestern Nova Scotia. Fishery Bulletin, 81(2), 357-362. [ Links ]
Carvalho, M. E., & Fontes, A. L. (2007). A carcinicultura no espaço litorâneo Sergipano. Revista da Fapese, 3(1), 87-112. [ Links ]
Carver, A. M., Wolcott, T. G., Wolcott, D. L., & Hines, A. H. (2005). Unnatural selection: effects of a male-focused size-selective fishery on reproductive potential of a blue crab population. Journal of Experimental Marine Biology and Ecology, 319(1), 29-41. [ Links ]
Castiglioni, D. S., & Coelho, P. A. (2011). Determinação da maturidade sexual de Ucides cordatus (Crustacea, Brachyura, Ucididae) em duas áreas de manguezal do litoral sul de Pernambuco, Brasil. Iheringia Série Zoologia, 101(1-2), 138-144. [ Links ]
Castilho, A. L., Gavio, M. A., Costa, R. C., Boschi, E. E., Bauer, R. T., & Fransozo, A. (2007). Latitudinal variation in population structure and reproductive pattern of the endemic South American shrimp Artemesia longinaris (Decapoda: Penaeoidea). Journal of Crustacean Biology, 27(4), 548-552. [ Links ]
Cobo, V. J., & Fransozo, A. (2003). External factors determining breeding season in the red mangrove crab Goniopsis cruentata (Latreille) (Crustacea, Brachyura, Grapsidae) on the São Paulo State northern coast, Brazil. Revista Brasileira de Zoologia, 20(2), 213-217. [ Links ]
Cobo, V. J., & Fransozo, A. (2005). Physiological maturity and relationship of growth and reproduction in the red mangrove crab Goniopsis cruentata (Latreille) (Brachyura, Grapsidae) on the coast of São Paulo, Brazil. Revista Brasileira de Zoologia, 22(1), 219-223. [ Links ]
Colpo, K. D., Chacur, M. M., Guimarães, F. J., & Negreiros-Fransozo, M. L. (2011). Subtropical Brazilian mangroves as a refuge of crab (Decapoda: Brachyura) diversity. Biodiversity and Conservation, 20(13), 3239-3250. [ Links ]
Conde, J. E., & Díaz, H. (1989). The mangrove tree crab Aratus pisonii in a tropical estuarine coastal lagoon. Estuarine, Coastal and Shelf Science, 28(6), 639-650. [ Links ]
Conover, D. O., & Munch, S. B. (2002). Sustaining fisheries yields over evolutionary time scales. Science, 297(5578), 94-96. [ Links ]
Corgos, A., & Freire, J. (2006). Morphometric and gonad maturity in the spider crab Maja brachydactyla: a comparison of methods for estimating size at maturity in species with determinate growth. Journal of Marine Science, 63(5), 851-859. [ Links ]
Costa, R. C., Fransozo, A., Freire, F. A. M., & Castilho, A. L. (2007). Abundance and ecological distribution of the “sete-barbas” shrimp Xiphopenaeus kroyeri (Heller, 1862) (Decapoda: Penaeoidea) in three bays of the Ubatuba region, southeastern Brazil. Gulf and Caribbean Research, 19, 33-41. [ Links ]
Costa, T. M., & Negreiros-Fransozo, M. L. (1996). Fecundidade de Callinectes danae Smith, 1869 (Crustacea, Decapoda, Portunidae) na região de Ubatuba (SP), Brasil. Arquivos de Biologia e Tecnologia, 39, 393-400. [ Links ]
Díaz, H., & Conde, J. E. (1988). On the food sources for the mangrove tree crab Aratus pisonii (Brachyura: Grapsidae). Biotropica, 20, 348-350. [ Links ]
Díaz, H., & Conde, J. E. (1989). Population dynamics and life history of the mangrove crab Aratus pisonii (Brachyura, Grapsidae) in a marine environment. Bulletin of Marine Science, 45(1), 148-163. [ Links ]
Diele, K., Koch, V., & Saint-Paul, U. 2005. Population structure, catch composition and CPUE of the artisanally harvested mangrove crab Ucides cordatus (Ocypodidae) in the Caeté estuary, North Brazil: indications for overfishing? Aquatic Living Resources, 18(2), 169-178. [ Links ]
Ferreira, A. C., Ganade, G., Freire, F. A. M., & Attayde, J. L. (2013). Propagule predation in a Neotropical mangrove: the role of the Grapsid crab Goniopsis cruentata. Hydrobiologia, 707(1), 135-146. [ Links ]
Fisher, J. A. D, Frank, K. T., & Leggett, W. C. (2010). Global variation in marine fish body size and its role in biodiversity–ecosystem functioning. Marine Ecology Progress Series, 405, 1-13. [ Links ]
Fox, C. W., & Czesak, M. L. 2000. Evolutionary ecology of progeny size in Arthropods. Annual Review of Entomology, 45(1), 341-369. [ Links ]
Guerra, R. S., Nascimento, M. M. F., Miesch, S., Najafzadeh, M. J., Ribeiro, R. O., Ostrensky, A., Hoog, G. S., Vicente, V. A., & Boeger, W. A. (2013). Black yeast biota in the mangrove, in search of the origin of the lethargic crab disease (LCD). Mycopathologia, 175(5-6), 421-430. [ Links ]
Hartnoll, R. G., & Gould, P. (1988). Brachyuran life history strategies and the optimization of egg production. Symposia of the Zoological Society of London, 59, 1-9. [ Links ]
Hines, A. H. (1989). Geographic variation in size at maturity in Brachyuran crabs. Bulletin of Marine Science, 45(2), 356-368. [ Links ]
Hirose, G. L., Fransozo, V., Tropea, C., Lopez-Greco, L. S., & Negreiros-Fransozo, M. L. (2012). Comparison of body size, relative growth and size at onset sexual maturity of Uca uruguayensis (Crustacea: Decapoda: Ocypodidae) from different latitudes in the southwestern Atlantic. Journal of the Marine Biological Association of the United Kingdom, 93(3), 781-788. [ Links ]
Jones, M. B., & Simons, M. J. (1983). Latitudinal variation in reproductive characteristics of a mud crab, Helice crassa (Grapsidae). Bulletin of Marine Science, 33(3), 656-670. [ Links ]
Kristensen, E. (2008). Mangrove crabs as ecosystem engineers; with emphasis on sediment processes. Journal of Sea Research, 59(1), 30-43. [ Links ]
Lardies, M. A., & Castilla, J. C. 2001. Latitudinal variation in the reproductive biology of the commensal crab Pinnaxodes chilenis (Decapoda: Pinnotheridae) along the Chilean coast. Marine Biology, 139(6), 1125-1133. [ Links ]
Lee, S. Y. (1997). Potential trophic importance of the faecal material of the mangrove sesarmine crab Sesarma messa. Marine Ecology Progress Series, 159, 275-284. [ Links ]
Lee, S. Y. (1998). Ecological role of grapsid crabs in mangrove ecosystems: a review. Marine and Freshwater Research, 49, 335-343. [ Links ]
Lima-Gomes, R. C., Cobo, V. J., & Fransozo, A. (2011). Feeding behaviour and ecosystem role of the red mangrove crab Goniopsis cruentata (Latreille, 1803) (Decapoda, Grapsoidea) in a subtropical estuary on the Brazilian coast. Crustaceana, 84, 735-747. [ Links ]
Lira, J. J. P. R., Calado, T. C. S., & Araújo, M. S. L. C. (2013). Breeding period in the mangrove crab Goniopsis cruentata (Decapoda: Grapsidae) in Northeast Brazil. Revista Biología Tropical, 61, 29-38. [ Links ]
Ma, H., Townsend, H., Zhang, X., Sigrist, M., & Christensen, V. (2010). Using a fisheries ecosystem model with a water quality model to explore trophic and habitat impacts on a fisheries stock: A case study of the blue crab population in the Chesapeake Bay. Ecological Modelling, 221, 997-1004. [ Links ]
Maciel, D. C., & Alves, A. G. C. (2009). Conhecimentos e práticas locais relacionadas ao aratu Goniopsis cruentata (Latreille, 1803) em Barra de Sirinhaém, litoral sul de Pernambuco, Brasil. Biota Neotropica, 9, 29-36. [ Links ]
Macintosh, D. J. (1988). The ecology and physiology of decapods of mangrove swamps. Symposia of the Zoological Society of London, 59, 315-341. [ Links ]
Macintosh, D. J., Ashton, E. C., & Havanon, S. (2002). Mangrove rehabilitation and intertidal biodiversity: a study in the Ranong Mangrove Ecosystem, Thailand. Estuarine, Coastal and Shelf Science, 55, 331-345. [ Links ]
McNab, B. K. (1971). On the ecological significance of Bergmann’s rule. Ecology, 52, 845-854. [ Links ]
Melo, G. A. S. (1996). Manual de identificação dos Brachyura (caranguejos e siris) do litoral brasileiro. São Paulo: Plêiade. [ Links ]
Menezes, A. P. D., Araújo, M. S. L. C., & Calado, T. C. S. (2012). Bioecologia de Goniopsis cruentata (Latreille, 1803) (Decapoda, Grapsidae) do complexo estuarino-lagunar Mundaú/Manguaba, Alagoas, Brasil. Natural Resources, 2(2), 37-49. [ Links ]
Moura, N. F. O., & Coelho, P. A. (2003). Fecundidade de Goniopsis cruentata (Latreille, 1803) (Crustacea: Brachyura: Grapsidae) no manguezal do rio Paripe – Pernambuco – Brasil. Tropical Oceanography, 31(2), 127-133. [ Links ]
Moura, N. O. F., & Coelho, P. A. (2004). Maturidade sexual fisiológica em Goniopsis cruentata (Latreille) (Crustácea, Brachyura, Grapsidae) no Estuário de Paripe, Pernambuco, Brasil. Revista Brasileira de Zoologia, 21(4), 1011-1015. [ Links ]
Negreiros-Fransozo, M. L., Fransozo, A., González-Gordillo, J. I., & Bertini, G. (2002). First appraisal on releasing and reinvasion of decapod larvae in a subtropical estuary from Brazil. Acta Limnologica Brasiliensis, 14(2), 87-94. [ Links ]
Nerot, C., Meziane, T., Provost-Govrich, A., Rybarczyk, H., & Lee, S. Y. (2009). Role of grapsid crabs, Parasesarma erythrodactyla, in entry of mangrove leaves into an estuarine food web: a mesocosm study. Marine Biology, 156(11), 2343-2352. [ Links ]
Nordhaus, I., Wolff, M., & Diele, K. (2006). Litter processing and population food intake of the mangrove crab Ucides cordatus in a high intertidal forest in northern Brazil. Estuarine, Coastal and Shelf Science, 67(1), 239-250. [ Links ]
Parker, G. A., & Begon, M. B. (1986). Optimal egg size and clutch size: effects of environment and maternal phenotype. The American Naturalist, 128, 573-592. [ Links ]
Pauly, D., Christensen, V., Guénette, S., Pitcher, T. J., Sumaila, U. R., Walters, C. J., Watson, R., & Zeller, D. (2002). Towards sustainability in world fisheries. Nature, 418(6898), 689-695. [ Links ]
Pinheiro, M. A. A., & Terceiro, O. S. L. (2000). Fecundity and reproductive output of the speckled swimming crab Arenaeus cribrarius (Lamarck, 1818) (Brachyura, Portunidae). Crustaceana, 73(9), 1121-1137. [ Links ]
Piou, C., Berger, U., & Feller, I. C. (2009). Spatial structure of a leaf-removing crab population in a mangrove of North-Brazil. Wetlands Ecology and Management, 17(2), 93-106. [ Links ]
Powers, L. W., & Bliss, D. E. 1983. Terrestrial adaptations. In D. E. Bliss (Ed.), The Biology of Crustacea (pp. 271-333). New York: Academic Press. [ Links ]
Rodríguez, G. (1987). Structure and production in Neotropical mangroves. Tree, 2(9), 264-267. [ Links ]
Saenger, P., Hegerl, E. J., & Davie, J. D. (1983). Global status of mangrove ecosystems. Environmentalist, 3(3), 1-88. [ Links ]
Sampedro, M., González-Gurriarán, E., Freire, J., & Muiño, R. (1999). Morphometry and sexual maturity in the spider crab Maja squinado (Decapoda: Majidae) in Galicia, Spain. Journal of Crustacean Biology, 19, 578-592. [ Links ]
Sandrini-Neto, L., & Lana, P. C. (2012). Distribution patterns of the crab Ucides cordatus (Brachyura, Ucididae) at different spatial scales in subtropical mangroves of Paranaguá Bay (southern Brazil). Helgoland Marine Research, 66(2), 167-174. [ Links ]
Santos, M. C. F., Botelho, E. R. R. O., & Ivo, C. T. C. (2001). Biologia populacional e manejo da pesca de aratu, Goniopsis cruentata (Latreille, 1803) (Crustacea: Decapoda: Grapsidae) no litoral sul de Pernambuco–Brasil. Boletim Técnico-Científico do CEPENE, 9, 87-123. [ Links ]
Schaeffer-Novelli,Y., Cintrón-Molero, G., & Adaime, R. R. (1990). Variability of mangrove ecosystems along the Brazilian Coast. Estuaries, 13(2), 204-218. [ Links ]
Schaeffer-Novelli, Y., Cintrón-Molero, G., Soares, M. L. G., & De-Rosa, T. (2000). Brazilian mangroves. Aquatic Ecosystem Health and Management, 3(4), 561-570. [ Links ]
Schubart, C. D., Cuesta, J. A., & Felder, D. L. (2002). Glyptograpsidae, a new brachyuran family from Central America: larval and adult morphology, and a molecular phylogeny of the Grapsoidea. Journal of Crustacean Biology, 22(1), 28-44. [ Links ]
Silva, Z. S., & Oshiro, L. M. Y. (2002). Aspectos reprodutivos de Goniopsis cruentata (Latreille) (Crustacea, Brachyura, Grapsidae) na Baía de Sepetiba, Rio de Janeiro, Brasil. Revista Brasileira de Zoologia, 19(3), 907-914. [ Links ]
Smith, N. F., Wilcox, C., & Lessmann, J. M. (2009). Fiddler crab burrowing affects growth and production on the white mangrove (Laguncularia racemosa) in a restored Florida coastal marsh. Marine Biology, 156(11), 2255-2266. [ Links ]
Souza, L. P., & Silva, J. R. F. (2009). Morphology of the female reproductive system of the red-clawed mangrove tree crab (Goniopsis cruentata Latreille, 1803). Scientia Marina, 73(3), 527-539. [ Links ]
Souza, M. J. F. T., Dantas-Junior, J. F., Silva, F. C. B., Félix, D. C. F., & Santos, J. C. (2012). Estatística pesqueira da costa do Estado de Sergipe e Extremo norte da Bahia 2010. São Cristóvão: Editora UFS. [ Links ]
Timi, J. T., Lanfranchi, A. L., & Poulin, R. (2005). Is there a trade-off between fecundity and egg volume in the parasitic copepod Lernanthropus cynoscicola? Parasitology Research, 95(1), 1-4. [ Links ]
Wilson, K., & Hardy, I. C. W. (2002). Statistical analysis of sex ratios: an introduction. In I. C. W. Hardy (Ed.), Sex ratios: concepts and research methods (pp. 48-92). Cambridge: Cambridge University Press. [ Links ]
Zar, J. H. (2010). Biostatistical analysis. Upper Saddle River: Prentice-Hall/Pearson. [ Links ]
1. Programa de Pós-graduação em Ecologia e Conservação, Universidade Federal Sergipe. Av. Marechal Rondon, s/n, CEP: 49100-000, São Cristóvão-SE, Brasil; gustavo_lh@hotmail.com, laizes.s@hotmail.com, luana_resende2008@hotmail.com, douglas_biologo@yahoo.com.br
2. NEBECC, Group of Studies on Crustacean Biology, Ecology and Culture, Universidade Estadual Paulista. Distrito de Rubião Júnior, s/n, CEP: 18618-970, Botucatu, SP, Brasil.
3. Departamento de Zoologia, Instituto de Biociências, Universidade Estadual Paulista. Distrito de Rubião Júnior, s/n, CEP: 18618-970, Botucatu, SP, Brasil; mlnf@ibb.unesp.br
Received 31-VII-2014. Corrected 10-XII-2014. Accepted 19-I-2015.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}