











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.63 no.2 San José abr./jun. 2015
Influence of microhabitats on the performance of herbaceous species in areas of mature and secondary forest in the semiarid region of Brazil
Influencia de microhábitats en el desempeño de especies herbáceas en áreas de bosque maduro y secundario en la región semiárida de Brasil
Influencia de microhábitats en el desempeño de especies herbáceas en áreas de bosque maduro y secundario en la región semiárida de Brasil
Juliana Ramos de Andrade1*, Kleber Andrade da Silva2*, Josiene Maria Fraga dos Santos1, Danielle Melo dos Santos1, Thiago Pereira Guerra3* & Elcida de Lima Araújo1
Abstract
The conditions for plant establishment in mature forest are different from those found in disturbed areas. In dry environments, the herbaceous cover is the most important in the recolonization of deforested areas. It can, therefore, act as an ideal biological group for assessing how changes in habitat heterogeneity affect the resilience of dry forests. The aim of this research was to evaluate whether natural regeneration of the herbaceous stratum differed between areas of mature and secondary forest of Caatinga and to describe this process. The study took place in the Brazilian semiarid region during the rainy season 2011 (January to August), where fifty 1m² plots were set up, 25 allocated to the microhabitat established as “between canopies” and 25 to the microhabitat “under the canopy”. The herbaceous species selected for the study were Delilia biflora (Asteraceae), Gomphrena vaga (Amaranthaceae) and Pseudabutilon spicatum (Malvaceae), abundant species occurring in both areas. All individuals from the selected populations were counted, marked with sequential numbers, and the height of the stem was measured. Differences between areas, and in size and survival between microhabitats, were found only for the first two species. Fruit production was higher in the mature forest for the three species. The study concluded that: 1. The effect of the microhabitats “between canopies” and “under the canopy” in mature and secondary forest areas depends on the species considered; 2. Populations sensitive to light intensity differ in number of individuals, height and fruit production; and 3. The resilience of anthropogenic areas in semiarid environments can be characterized by the presence of spatial heterogeneity with regard to the emergence and survival of herbaceous seedlings, suggesting that the regeneration of disturbed areas may occur in patches. Rev. Biol. Trop. 63 (2): 357-368. Epub 2015 June 01.
Key words: demography, antropogenic area, microsite, plant canopy, herb.
Resumen
Las condiciones para el establecimiento de las plantas (agua, luz, nutrientes, etc.) en los fragmentos preservados se diferencian de las encontradas en áreas perturbadas. En ambientes secos, la cubierta herbácea predomina en la recolonización de áreas despejadas y puede actuar como un buen grupo biológico para evaluar los impactos de los cambios de la heterogeneidad de hábitats en el proceso de resiliencia de los bosques secos. Ante de esto, el objetivo fue evaluar si la regeneración natural de la capa herbácea difería entre áreas preservadas y áreas de Caatinga perturbadas y describir cómo ocurre esto. El estudio se realizó en la región semiárida brasileña durante la temporada de lluvias, donde 50 parcelas de 1m² fueron establecidas, 25 de las cuales se ubicaron en el microhábitat llamado “entre dosel” y 25 en “bajo dosel”. Las especies herbáceas seleccionadas para el estudio fueron: Delilia biflora (Asteraceae), Gomphrena vaga (Amaranthaceae) y Pseudabutilon spicatum (Malvaceae), especies abundantes que se producen en ambas áreas. Todos los individuos de las poblaciones seleccionadas se contaron, marcando con números secuenciales, y se midió la altura del tallo. Las diferencias entre las zonas, y el tamaño y supervivencia entre microhábitats, se encontraron solo para las dos primeras especies. La producción de frutos fue mayor en el bosque maduro para las tres especies. El estudio llega a la conclusión de que: 1. el efecto de la existencia de microhabitates “entre dosel” y “bajo dosel” en áreas preservadas y antropogénicas depende de la especie considerada; 2. las poblaciones sensibles a la variación de la intensidad luminosa presentan diferencias en cuanto al número de individuos, la altura y la producción de frutos de las plantas; 3. la resiliencia de las zonas antropogénicas de ambientes semiáridos se puede caracterizar por la existencia de heterogeneidad espacial en cuanto a la emergencia y supervivencia de plántulas herbáceas, lo que sugiere que la regeneración de áreas perturbadas puede ocurrir en parches.
Palabras clave: demografía, área antropogénica, micrositio, dosel, hierbas.
Most terrestrial habitats have been altered and fragmented due to human activities and pressure for plant use (Castelletti, Santos, Tabarelli, & Silva, 2003). Many areas after use are abandoned and regenerate naturally (Sampaio, Araújo, Salcedo, & Tiessen, 1998; Andrade, Pereira, Leite & Barbosa, 2005; Andrade, Santos, Lima, Lopes, Silva, & Araújo, 2007). However, the conditions for plant establishment vary from the conditions of mature forest areas because the anthropogenic modifications provide microclimatic changes, which may trigger structural changes in the population (Tews, Esther, Milton, & Jeltsh, 2006).
Microclimatic conditions in secondary forest areas (such as light availability, water availability, temperature, among others) are different from those found in mature forest areas (Crozier, & Boerner, 1984; Ferguson, Vandermeer, Morales, & Griffith 2003; Tews et al., 2006). These differences occur because the removal of the vegetation cover alters the local microclimate, and the influence of some biotic and abiotic factors turns more intense when compared to preserved habitats (Andrade et al., 2007; Kostrakiewicz, 2009; Legras, Vander Wall, & Board, 2010). These alterations enable the formation of new microhabitats, such as open areas exposed to sunlight, soil with greater or lesser ability to retain water or differentiation in nutrient availability, according to current studies (Brooks, 1999; Titus, & Tsuyuzaki, 2003; Gómez-Aparicio, Gómez, & Zamora, 2005). The existence of several microhabitats in a particular area can, on the one hand, provide an increase in species richness but, on the other hand, can influence population dynamics, facilitating or limiting the establishment of other species and productivity of fruits and seeds (Brooks, 1999; Singleton, Gardescu, Marks, & Geber, 2001; Araújo, Silva, Ferraz, Sampaio, & Silva, 2005; Reis, Araújo, Ferraz, & Moura, 2006; Andrade et al., 2007; Yu, Bell, Sternberg, & Kutiel, 2008; García-Cháves, Montaña, Perroni, Sosa, & García-Licona, 2014).
Some studies suggest that semiarid environments considered mature are shaded by trees, which provide differentiated microhabitats that favor the productivity of the herbaceous understory, due to a reduction of temperature and evaporation. For example, Grouzis and Akpo (1997) in the semiarid region of the Sahel, Senegal, evaluated the effects of forest cover on underground and aerial phytomass of the herbaceous strata and found that the aerial phytomass was 1.5 to four times higher, and that the phytomass of roots was about twice as high under the canopy. This increase in productivity of herbaceous species was attributed to better climate and fertile soil under the canopy.
Due to the evidence provided, this study hypothesizes that microhabitats have significant influence on the dynamics of herbaceous populations, but this influence will depend on the conservation status of the respective area. Thus, this study proposes to test whether: 1 - population dynamics, considering density, number of individuals born, number of dead individuals, plant height and fruit production of herbaceous plants, differs between areas of mature and secondary forest; 2 - population dynamics in different herbaceous microhabitats (sections between canopies and under the canopy of woody plants) differs between areas of mature and secondary forest; and to describe how this process occurs.
Material and methods
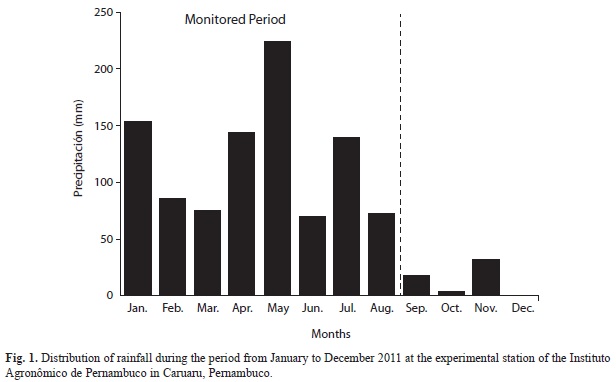
Study area: The study was conducted in the semiarid region of Brazil (Caatinga) in areas of mature and secondary forests, located in the ‘agreste’ of the state of Pernambuco, in the Pernambuco Agronomic Institute (IPA-Instituto Agronômico de Pernambuco-Estação Experimental José Nilson de Melo), municipality of Caruaru (8o14’18” S - 35o55’20” W, 537 masl). The climate is semiarid, Köppen type BSh (Köppen, 1948), with average annual rainfall of 710 mm, concentrated between March and August, and average temperature of 22.7oC. Despite the well-defined climatic seasonality, sporadic rainfall may occur during the dry season, and dry spells may occur during the rainy season. Moreover, the start of the season may be delayed or brought forward, as took place in the period of this study, when the rainy season of 2011 began early in January. Thus, the rainy season included in this study covered eight months.
The strong climatic seasonality determines the deciduousness of the woody flora during the dry season and the increased exposure of the herbaceous flora in the rainy season. The rainfall distribution during the monitored period, measured at the actual Experimental Station, can be found in figure 1.
The IPA was initially composed of a single area of natural Caatinga vegetation occupying an area of approximately 190 hectares but, due to research on agriculture and livestock, this has been reduced to a fragment 20 hectares. For about 50 years, this fragment (20 hectares) has been preserved from human disturbance. According to the floristic survey by Alcoforado-Filho, Sampaio and Rodal (2003) the woody flora of this area of mature forest displays high richness of Leguminosae and Euphorbiaceae. The herbaceous component is represented mainly by the families Poaceae, Asteraceae, Malvaceae, Convolvulaceae and Euphorbiaceae (Araújo et al., 2005; Reis et al., 2006).
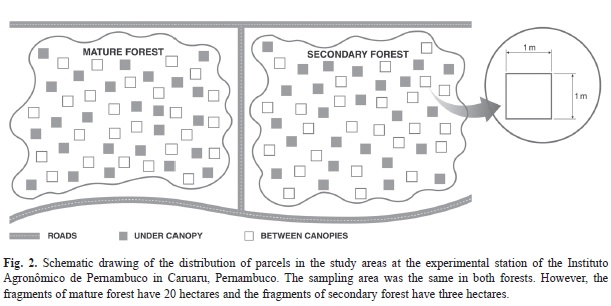
In 1994, a stretch of approximately 3 ha about 5 m away from the mature forest area has suffered clear-cutting for cultivation of the gigant palm (Opuntia ficus-indica Mill.) and neither fire nor pesticides were used for preparing the land (Fig. 2). About six months after this palm was planted, the area was abandoned and has since been regenerating, with no new records of human intervention.
Currently, vegetation has been established in the disturbed area, but it is in the early stages of succession and shows different microclimatic conditions from the area of mature forest. Its shrub-arboreal vegetation has a lower height, lower density and more open canopy when compared with the area of mature forest. Consequently, there is greater light penetration and higher temperatures for regeneration of herbaceous vegetation, whilst, in the area of mature forest, the soil is more shaded because the canopy is almost continuous (Lopes et al., 2012; Santos et al., 2013).
Selection of species and experimental design: The species selected for the study were the therophytes (species that spend an unfavorable season in seed form; Raunkiaer, 1934) with distribution in the American tropics, Delilia biflora (L.) Kuntze (Asteraceae), Gomphrena vaga Mart. (Amaranthaceae) and Pseudabutilon spicatum (Kunth) RE Fr (Malvaceae); these species populations were abundant and occurred in both areas of mature and secondary forests (Reis et al., 2006; Santos, Santos, Lopes, Silva, Sampaio, & Araújo, 2013). In general, herbaceous component performs an important role in the maintenance and restoring the biodiversity, by interfering with the recruitment of seedlings, being an additional source of food for fauna (providing pollen, nectar and resin), and assist in the retention of seeds in the topsoil through intertwining their roots (Araújo, & Ferraz, 2003; Lorenzon, Matrangolo, & Schoereder, 2003; Araújo, & Ferraz, 2003; Santos et al., 2013). Additionally, recent research has indicated the high importance of herbaceous species in the cure and treatment of diseases (Silva et al., 2013; Lozano, Araújo, Medeiros, & Albuquerque, 2014).
Monitoring was conducted during the rainy season of 2011 (January-August 2011), as it is in this season that woody plants have leaves, shade the soil and provide differentiated microhabitats that influence the dynamics of herbaceous populations. Fifty plots (total of 100 plots) were established in both areas of mature forest and secondary forests, equally divided between two types of microhabitats. In the microhabitat “under the canopy”, plots were allocated under tree canopies and in the microhabitat “between canopies”, plots were set up in the stretches between the trees, where there is direct incidence of sunlight (Fig. 2).
The 1x1m plots were set up using picket fences of 0.30m at a minimum distance of 1m from one another. Within the plots, all individuals from the selected populations were counted, marked with sequential numbers, and the height of the stem was measured. Measurements of height were taken with the aid of a ruler or tape measure. The marking of individuals was made with plastic labels that were attached to the base of the herbaceous plants with a wire.
Monthly visits were carried out for monitoring the growth of the individuals, recording new individuals (births=when seedling emergence was observed) and the number of deaths in the populations (the individuals that disappeared from the plots between sampling intervals or dried up and fell).
To quantify and evaluate whether there were differences in fruit production of individuals from the areas of both mature and secondary forests in the microhabitats, 25 individuals from population per microhabitat, established outside the plots, were randomly selected, totaling 50 individuals per area, which were monitored monthly until fruit production. As soon as fruits appeared, individuals were collected for counting the fruit.
Processing and data analysis: The survival of the population was described by the percentage of surviving individuals from the cohort at the start of the study. The normality of the mean data of density, number of individuals born, number of dead individuals and height was assessed using the Kolmogorov-Smirnov test, which clarified that the data were not normally distributed. Monthly differences in density, total birth, mortality and absolute growth (height) of the herbaceous plants between the areas (mature and secondary forests) and microhabitats (under the canopy and between canopies) were evaluated by the nonparametric Kruskal-Wallis test (Zar, 1996). Fruit production was quantified using its absolute value and the differences in production between areas and microhabitats were analyzed using the one-sample chi-square test (adhesion) with equal expected proportions (Ayres, Ayres Júnior, Ayres, & Santos, 2007). The analyses were performed using the program BioEstat 5.0.
Results
Density of herbaceous populations between areas and microhabitats: In the initial survey (January 2011), the populations found in the 50m² of the mature forest area were made up of 304 individuals of Delilia biflora, 331 individuals of Gomphrena vaga and 41 individuals of Peudabutilon spicatum. Regarding the density of individuals observed in each microhabitat, the total number of individuals of D. biflora recorded were 207 in the microhabitat “between canopies”, and 97 in the microhabitat “under the canopy”. For G. vaga, 199 individuals were found “between canopies” and 132 “under the canopy”; and for P. spicatum the microhabitat “between canopies” was represented by 37 individuals and the microhabitat “under the canopy” by four individuals.
In the disturbed area, 628 individuals of D. biflora, 202 individuals of G. vaga and 15 individuals of P. spicatum were found in the initial survey. For the population of D. biflora, 196 individuals were established in the microhabitat “between canopies” and 432 in the microhabitat “under the canopy”. For G. vaga, 57 individuals were found in the microhabitat “between canopies” and 145 in the microhabitat “under the canopy”. Six and nine individuals of P. spicatum were recorded in the microhabitats “between canopies” and “under the canopy”, respectively.
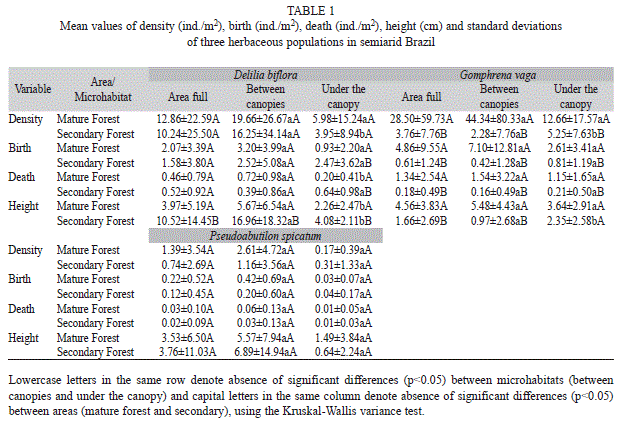
The population of P. spicatum did not show a significant difference in density between areas and microhabitats (Table 1). The population of D. biflora showed no significant difference between the areas and only a significant difference between the microhabitats of the area of secondary forest (H=6.26, p=0.01) (Table 1).
The mean density of G. vaga showed a significant difference between areas (H=18.44, p<0.01), being greater in the mature forest area. A significant difference was also verified between microhabitats (Table 1). For the mature forest area its mean density was found greater in the “between canopies” microhabitat. In the case of the secondary forest area, the mean density of G. vaga was found greater in the “under the canopy” microhabitat (Table 1).
Birth, death and survival of herbaceous populations between areas and microhabitats: The number of births varied between populations, between areas and between microhabitats. In the area of the mature forest, the number of births was higher for all populations in the “between canopies” microhabitat. For the area of secondary forest, only G. vaga had a greater number of births in the “under the canopy” microhabitat (Table 1 and Table 2).
In the population of Delilia biflora, the mean number of births showed no significant difference between the areas. Regarding the average number of dead individuals, D. biflora also showed no significant difference between the areas. In the secondary forest area, the mean number of births of D. biflora did not differ between microhabitats (“between canopies” x “under the canopy”) (Table 1).
The population of G. vaga showed significant difference in the mean number of births (H=19.50, p<0.01) and deaths (H=18.78, p<0.01) between areas, and no significant difference in the mean number of births and deaths between microhabitats in the same area. However, the influence of microhabitat (“between canopies” x “under the canopy”) on the mean number of births and deaths, differed between the mature and secondary forests areas (Table 1 and Table 2).
Significant differences regarding the mean number of births and deaths and between microhabitats between both areas were not recorded for P. spicatum (Table 1 and Table 2).
Regarding survival, in the mature forest area, 85.50% of the individuals of D. biflora survived until the end of the surveys in the “between canopies” microhabitat, and 95.87% in the “under the canopy” microhabitat. In the area of secondary forest, 88.77% of the individuals of D. biflora recruited in the “between canopies” microhabitat, and 74.53% of the individuals recruited “under the canopy” survived until the last month of the study.
For the population of G. vaga in the mature forest area, the percentage of survival of individuals recruited at the start of the surveys in the “between canopies” microhabitat was 73.36%, and “under the canopy” was 54.54%. In the area of secondary forest, G. vaga showed 59.64% survival in the “between canopies” microhabitat, and 72.41% in the “under the canopy” microhabitat.
When individuals of P. spicatum were evaluated in the area of mature forest, 89.18% were found in the “between canopies” microhabitat, and 50% “under the canopy”. In the area of secondary forest, 100% survived in the “between canopies” microhabitat, and 77.7% “under the canopy”.
Absolute growth and fruit production: The average height of individuals of Delilia biflora differed significantly between areas (H=5.08, p=0.02) (mature forest and secondary) and between the microhabitats in each area, the mean height being greater for the area of secondary forest (Table 1). The mean height of G. vaga differed significantly between areas (H=16.37). In the secondary forest area, a significant difference in mean height of G. vaga was recorded between microhabitats (H=4.43, p<0.01), which did not occur in the area of mature forest. The influence of the “between canopies” microhabitat was different between the areas of mature forest and secondary (H=14.24, p=0.02); the plants in this microhabitat condition grew to a greater mean height in the mature forest area. For P. spicatum, there was no significant difference in the mean height of individuals between areas and between microhabitats (Table 1).
Fruit production of D. biflora showed a significant difference between areas (p<0.01) (Table 3). In the mature forest area, there was a significant difference in production between microhabitats (p<0.01), ranging from 299 (“between canopies”) to 1878 (“under the canopy”) (Table 3). In the secondary forest area, production of D. biflora differed significantly (p<0.01) from 310 fruits in the microhabitat “under the canopy” to 736 fruits in the microhabitat “between canopies” (Table 3).
The number of fruits of G. vaga did not differ significantly between the study areas (p=0.91). The number of fruits collected in the mature forest (158 fruits) was similar to that recorded for the secondary forest area (160 fruits). However, in the mature forest area, G. vaga produced more fruit in the microhabitat “between canopies” (106 fruits), showing a significant difference between microhabitats (p<0.01) and, in the secondary forest, this species produced more fruit in the microhabitat “under the canopy” (87 fruits), although this difference was not significant according to the chi-square test (p=0.26) (Table 3).
The fruit production of P. spicatum was significantly different only between microhabitats of the disturbed area (p<0.01) (Table 3).
Considering both areas of mature and secondary forests, a greater fruit production was recorded under direct sunlight conditions, for which a double number of fruits were produced when compared to the diffused light area, for the three species (Table 3).
Discussion
The dynamics (density, birth, deaths and height) and survival of Delilia biflora and Gomphrena vaga differed between the areas and between the microhabitats for some of the variables analyzed. On the other hand, Pseudabutilon spicatum showed no significant difference in density, number of births, number of dead individuals and between areas or microhabitats, indicating that the different conditions of microhabitat (“under the canopy” and “between canopies”) did not influence the dynamics of the species. Some authors admit that other abiotic factors, such as local climatic characteristics and/or rainfall, may change the plasticity of the species over time (Belsky, 1990; Salo, 2004; Wang, 2005; Flores-Torres, & Montaña, 2012; García-Cháves et al., 2014). It is possible that this may have occurred to P. spicatum, but it is also possible that light, either between canopies or under the canopy, is not a critical factor in the dynamics of this population, and consequently, this species may not be considered a good model for assessing the impact of human actions on the recovery of abandoned areas.
Since G. vaga had a higher mean density in the mature forest area, it showed a tendency to develop best in undisturbed areas, specifically in microhabitats receiving a greater amount of light (between canopies). This result agrees with other studies in semiarid environments which record that there is a higher density of the herbaceous component in open segments, where sunlight directly reaches the herbaceous stratum, facilitating recruitment (Crozier, & Boerner, 1984; Kessler, 1992; Belsky, 1994; Aguilera, & Lauenroth, 1995; Mordelet, & Menaut, 1995; Silva et al., 2008; Flores-Torres, & Montaña, 2012).
The substantial number of individuals of D. biflora in the secondary forest, specifically in the microhabitat “under the canopy”, suggests that it is possibly a colonizing species of disturbed areas (which are in early stages of succession). Since the canopy in these areas is less dense than that of existing areas of mature forest, one can assume that the more shaded locations of areas of secondary forest offer a microclimatic condition similar to that found in open stretches of mature forest. Therefore, individuals that settle in secondary forest, experience microclimatic conditions (such as light availability, water availability, temperature, etc.) that are distinct from those found in mature forest, because the removal of the plant cover, alters the local microclimate and, additionally, the influence of biotic and abiotic factors becomes of higher intensity, when compared to the one that occurs in preserved habitats (Crozier, & Boerner, 1984; Ferguson et al., 2003; Tews et al., 2006; Andrade et al., 2007; Kostrakiewicz, 2009).
The occupation of areas with less dense woody vegetation by herbaceous species (similar to that observed in the area of secondary forest) was also found by Riginos, Grace, Augustine and Young (2009), when recording that shade provided by more isolated trees, or those that are spaced apart, may have a positive effect in the development of herbaceous species, whereas shade provided by a high density of woody plants that are close together, negatively affects herbaceous populations.
Differences found in this study regarding the density of herbaceous plants between the microhabitats (“between canopies” and “under the canopy”) showed that the effect of the canopy, that is, the existence of more or less illuminated microhabitats due to the presence of woody plants, is an important factor for the establishment of herbaceous seedlings, and the level of shading may be more relevant than the change in the area itself, which has also been recorded by Gómez-Aparicio et al. (2005) and Aguilera and Lauenroth (1995).
The number of dead individuals varied greatly between areas and microhabitats. The large number of dead individuals found in the microhabitats may be related to several factors. For example, individuals established in the “between canopies” microhabitat lose faster water to the environment, suffering water deficit more quickly and more intensely (characteristic of the caatinga), being unable to keep themselves alive in these microhabitats. In addition to water stress, other environmental factors can cause mortality of individuals in both microhabitats, such as the impact of rain on newly germinated seedlings or frail individuals who survived the previous drought (Andrade et al., 2007; Santos, Andrade, Lima, Silva, & Araújo, 2007); intra and interspecific competition for resources (Nordbakken, Rydgren, & Okland, 2004; Suzuki, Kudoh, & Kachi, 2004; Vilá, Bartolomeus, Gimeno, Traveset, & Moragues, 2006); attack by pathogens (Araújo, & Ferraz, 2003; Silveira, Araújo, Araújo, & Willadino, 2005); and herbivory (Belsky, 1990; Hanley, 1998; Leimu, & Lehtila, 2006).
The different environmental conditions also influenced the survival of the species, since they varied between areas and between microhabitats depending on the species considered. Although the species in this study showed a trend toward greater survival in open areas, studies such as that of Gómez-Aparicio et al. (2005) found different results, where the herbaceous species showed lower survival rates in open areas.
In the literature, the development of herbaceous species follows two types of models. The first model includes the herbaceous species that develop in more shaded locations, below the crown of woody plants; these are capable of inducing morphological changes in herbaceous species, so that they can increase the biomass produced (Kessler, 1992; Mordelet, & Menaut, 1995; Zhang, Wang, Liu, & Li, 2006; Legras et al., 2010). The second model encompasses the herbaceous species that thrive best in open areas (Crozier, & Boerner, 1984; Kessler, 1992; Belsky, 1994; Mordelet, & Menaut, 1995; Aguilera, & Lauenroth 1995; Silva et al., 2008). In this study, differences in the mean heights of plants between microhabitats confirmed the influence of establishment conditions on plant growth, showing that direct light is an important prerequisite for the development of the studied species. D. biflora and G. vaga had greater mean heights in the direct light and P. spicatum in the shaded locations.
Singleton et al. (2001) found differences in seed production between preserved and disturbed areas: the oldest forest showed higher seed productivity when compared to disturbed areas (secondary forest). The discovery that the conservation status of a forest influences the productivity of seeds was also confirmed in this study but, depending on the species, it is not necessarily the preserved habitat that exhibits greater production, since we observed that G. vaga and P. spicatum had higher fruit production in the disturbed area.
Considering that the mature forest area is located only 5 m away from the secondary forest area, the differences recorded showed that there are changes in the herbaceous populations dynamics during the natural regeneration of areas that suffer human intervention, which need to be characterized and quantified in order to understand the process of resilience in dry environments.
Our results shows that directly or partially illuminated microhabitats can influence the dynamics of herbaceous populations established in areas with different status conservation. However, due to the occurrence of variation in the precipitation interannual in areas of dry environments, it is necessary to monitor other rainy seasons to determine an average trend of the dynamic influence of the microhabitat annual populations. In addition, the influence of the “between canopies” or “under the canopy” microhabitats depends on the species considered. Populations sensitive to variation in light intensity vary in the number of individuals, height and fruit production.
The resilience of areas of secondary forest in semiarid environments can be characterized by the existence of spatial heterogeneity regarding the emergence and survival of herbaceous seedlings. Consequently, the role of these in maintaining the soil (avoiding erosion) and retaining seeds from other species (woody and herbaceous) is not uniform, indicating that regeneration of disturbed areas may occur in patches. Moreover, the fact that fruit production is higher in the mature forest for one of the species underscores the importance of habitat conservation for renewal of the seed stock in the soil and availability of propagules for the renewal of certain populations.
Acknowledgments
The authors thank CNPq for financial support to the project (process 478087/04-7 and 4772392009-9) and the MSc scholarship to the first author, the Estação Experimental José Nilson de Melo of the Instituto Agronômico de Pernambuco-IPA for allowing access to the study area and offering accommodation during the fieldwork, the Universidade Federal Rural de Pernambuco for logistical support, the Graduate Program in Botany of the UFRPE and institutional support to researchers from the Laboratório de Ecologia Vegetal dos Ecossistemas Naturais-LEVEN for help in fieldwork and data processing.
References
Aguilera, M. & Lauenroth, K. W. (1995). Influence of gap disturbances and type of microsites on seedling establishment in Bouteloua gracilis. Journal of Ecology, 83, 87-97. [ Links ]
Alcoforado-Filho, F. G., Sampaio, E. V. S. B., & Rodal, M. J. N. (2003). Florística e fitossociologia de um remanescente de vegetação caducifólia espinhosa arbórea em Caruaru. Acta Botânica Brasileira, 17, 287-303. [ Links ]
Andrade, J. R., Santos, J. M. F. F., Lima, E. M., Lopes, C. G. R., Silva, K. A., & Araújo, E. L. (2007). Estudo populacional de Panicum trichoides Swart. (Poaceae) em uma área de Caatinga em Caruaru, Pernambuco. Revista Brasileira de Biociências, 5, 858-860. [ Links ]
Andrade, L. A., Pereira, I. M., Leite, U. T., & Barbosa, M. R. V. (2005). Análise da cobertura de duas fitofisionomias de Caatinga, com diferentes históricos de uso, no Município de São João do Cariri, Estado da Paraíba. Cerne, 11, 253-262. [ Links ]
Araújo, E. L., Silva, K. A., Ferraz, E. M. N., Sampaio, E. V. S. B., & Silva, S. I. (2005). Diversidade de herbáceas em microhabitates rochoso, plano e ciliar em uma área de Caatinga, Caruaru- PE. Acta Botânica Brasílica, 19, 285-294. [ Links ]
Araújo, E. L., & Ferraz, E. M. N. (2003). Processos ecológicos mantenedores da diversidade vegetal na Caatinga: estado atual do conhecimento. In V. Claudino-Sales (Org.), Ecossistemas brasileiros: manejo e conservação. Fortaleza, Brazil: Expressão Gráfica. [ Links ]
Ayres, M., Ayres Júnior, M., Ayres, D. L., & Santos, A. A. (2007). Bioestat-Aplicações estatísticas nas áreas das ciências biomédicas. Belém: Ong Mamiraua. [ Links ]
Belsky, A. J. (1990). Tree/grass ratios in East African savannas: a comparison of existing models. Journal of Biogeography, 17, 483-489. [ Links ]
Belsky, A. J. (1994). Influences of trees on savanna productivity: Tests of shade, nutrients, and tree-grass competition. Journal of Ecology, 75, 922-932. [ Links ]
Brooks, M. L. (1999). Habitat invasibility and dominance by alien annual plants in the western Mojave Desert. Biological Invasions, 1, 325-337. [ Links ]
Castelletti, C. H. M., Santos, A. M. M., Tabarelli, M., & Silva, J. M. C. (2003). Quanto ainda resta da Caatinga? Uma estimativa preliminar. In I. R. Leal, M. Tabarelli, & J. M. C. Silva (Eds.), Ecologia e Conservação da Caatinga. Recife, Brazil: University Publishing UFPE. [ Links ]
Crozier, C. R., & Boerner, R. E. J. (1984). Correlations of Understory Herb Distribution Patterns with Microhabitats under Different Tree Species in a Mixed Mesophytic Forest. Oecologia, 62, 337-343. [ Links ]
Ferguson, B. G., Vandermeer, J., Morales, H., & Griffith, D. M. (2003). Post-agricultural succession in El Péten, Guatemala. Conservation Biology, 17, 818-828. [ Links ]
Flores-Torres, A., & Montaña, C. (2012). Recruiting mechanisms of Cylindropuntia leptocaulis (Cactaceae) in the Southern Chihuahuan Desert. Journal of Arid Environments, 84, 63-70. [ Links ]
García-Cháves, J. H., Montaña, C., Perroni, Y., Sosa, V. J., & García-Licona, J. B. (2014). The relative importance of solar radiation and soil origin in cactos seedling survivorship at two spatial scales: plant association and microhabitat. Journal of Vegetation Science, 25, 668-680. [ Links ]
Gómez-Aparicio, L., Gómez, J. M., & Zamora, R. (2005). Microhabitats shift rank in suitability for seedling establishment depending on habitat type and climate. Journal of Ecology, 93, 1194-1202. [ Links ]
Grouzis, M., & Akpo, L. E. (1997). Influence of tree cover on herbaceous layer above and below ground phytomass in Sahelian zone. Journal of Arid Environment, 35, 285-296. [ Links ]
Hanley, M. E. (1998). Seedling herbivory, community composition and plant life history traits. Perspectives in Plant Ecology, Evolution and Systematics, 1(2), 191-205. [ Links ]
Kessler, J. J. (1992). The influence of karité (Vitellaria paradoxa) and néré (Parkia biglobosa) trees on sorghum production in Burkina Faso. Agroforestry Systems, 17, 297-118. [ Links ]
Köeppen, W. (1948). Climatologia: con un estudio de los climas de la Tierra. México: Fondo de Cultura Económica. [ Links ]
Kostrakiewicz, K. (2009). The influence of shadow created by adjacent plants on phenotypic plasticity of endangered species Trollius europaeus L (Ranunculaceae). Polish Journal of Ecology, 4, 625-634. [ Links ]
Legras, E. C., Vander Wall, S. B., & Board, D. I. (2010). The role of germination microsite in the establishment of sugar pine and Jeffrey pine seedlings. Forest Ecology and Management, 260, 806-813. [ Links ]
Leimu, R. & Lehtila, K. (2006). Effects of two type of herbivores on the population dynamics of a perennial herb. Basic and Applied Ecology, 7, 224-235. [ Links ]
Lopes, C. G. R., Ferraz, E. M. N., Castro, C. C., Lima, E. N., Santos, J. M. F. F., Santos, D. M., & Araújo, E. L. (2012). Forest succession and distance from preserved patches in the Brazilian semiarid region. Forest Ecology Managenment, 271, 115-123. [ Links ]
Lorenzon, M. C. A., Matrangolo, C. A., & Schoereder, J. H. (2003). Flora visitada pelas abelhas eussociais (Hymenoptera, Apidae) na Serra da Capivara, em caatinga do sul do Piauí. Neotropical Entomology, 32, 27-36. [ Links ]
Lozano, A. B., Araújo, E. L., Medeiros, M. F. T., & Albuquerque, U. P. (2014). The apparency hypothesis applied to a local pharmacopoeia in the Brazilian northeast. Journal of Ehtnobiology and Ethnomedicine, 10, 1-17. [ Links ]
Mordelet, P. & Menaut, J. C. (1995). Influence of trees on above-ground production dynamics of grasses in a humid savanna. Journal Vegetable Science, 6, 223-228. [ Links ]
Nordbakken, J. F., Rydgren, K., & Okland, R. H. (2004). Demography and population dynamics of Drosera anglica and D. rotundifolia. Journal of Ecology, 92, 110-121. [ Links ]
Raunkiaer, C. (1934). Life forms of plants and statistical plant geography. Oxford, UK: Clarendon Press. [ Links ]
Reis, A. M. S., Araújo, E. L., Ferraz, E. M. N., & Moura, N. A. (2006). Variações interanuais na florística e fitossociologia do componente herbáceo de uma área de Caatinga, Pernambuco, Brazil. Acta Botânica Brasílica, 29, 497-508. [ Links ]
Riginos, C., Grace, J. B., Augustine, D. J., & Young, T. P. (2009). Local versus Landscape- Scale Effects of Savanna Trees on Grasses. Journal of Ecology, 97, 1337-1345. [ Links ]
Santos, J. M. F. F., Andrade, J. R., Lima, E. M., Silva, K. A., & Araújo, E. L. (2007). Dinâmica populacional de uma espécie herbácea em uma área de floresta tropical seca no Nordeste do Brasil. Revista Brasileira de Biociências, 5, 855-857. [ Links ]
Santos, J. M. F. F., Santos, D. M., Lopes, C. G. R., Silva, K. A., Sampaio, E. V. S. B., & Araújo, E. L. (2013). Natural regeneration of the herbaceous community in a semiarid region in Northeastern Brazil. Environmental Monitoring and Assessment, 185, 8287-8302. [ Links ]
Salo, L. F. (2004). Population dynamics of red brome (Bromus madritensis subsp. rubens): times for concern, opportunities for management. Journal of Arid Evironments, 57, 291-296. [ Links ]
Sampaio, E. V. S. B., Araújo, E. L., Salcedo, I. H., & Tiessen, H. (1998). Regeneração da vegetação após corte e queima, em Serra Talhada, PE. Revista Brasileira de Pesquisa Agropecuária, 33, 621-632. [ Links ]
Silva, A. C. O., Santana, E. F., Saraiva, A. M., Coutinho, F. N., Castro, H. A., Pisciottano, M. N. C., & Albuquerque, U. P. (2013). Which approach is more effective in the selection of plants with antimicrobial activity? Evidence-Based Complementary and Alternative Medicine, 1-9. [ Links ]
Silva, K. A., Lima, E. N., Santos, J. M. F. F., Andrade, J. R., Santos, D. M., Sampaio, E. V. S. B., & Araújo, E. L. (2008). Dinâmica de gramíneas em uma área de Caatinga de Pernambuco-Brasil. In U. P. Albuquerque, A. N. Moura, & E. L. Araújo (Eds.), Biodiversidade, potencial econômico e processos eco fisiológicos em ecossistemas nordestinos. Recife, Brazil: Comunigraf. [ Links ]
Silveira, A. P., Araújo, E. L., Araújo, F. S., & Willadino, L. G. (2005). Predação de frutos e germinação de sementes em Auxemma oncocalyx (Allemão) Baill. e Auxemma glazioviana Taub. In R. J. M. C. Nogueira, E. L. Araújo, L. G. Willadino, & U. M. T. Cavalcante (Eds.), Estresses ambientais: danos e benefícios em plantas. Recife, Brazil: MXM Gráfica e Editora. [ Links ]
Suzuki, Z. O., Kudoh, H., & Kachi, N. (2004). Spatial and temporal variations in mortality of the biennial plant, Lysimachia rubida: effects of intraspecific competition and environmental heterogeneity. Journal of Ecology, 91, 114-125. [ Links ]
Singleton, R., Gardescu, S., Marks, P. L., & Geber, M. A. (2001). Forest herb colonization of postagricultural forests in central New York State, USA. Journal of Ecology, 89, 325-338. [ Links ]
Tews, J., Esther, A., Milton, S. J., & Jeltsh, F. (2006). Linking a population model with an ecosystem model: Assessing the impact of land use and climate change on savanna shrub cover dynamics. Ecological Modelling, 195, 219-228. [ Links ]
Titus, J. H., & Tsuyuzaki, S. (2003). Distribution of plants in relation to microsites on recent volcanic substrates on Mount Koma, Hokkaido, Japan. Ecological Research, 18, 91-98. [ Links ]
Vilá, M., Bartolomeus, I., Gimeno, I., Traveset, A., & Moragues, E. (2006). Demography of the Invasive Geophyte Oxalis pes-caprae Across a Mediterranean Island. Annals of Botany, 97, 1055-1062. [ Links ]
Yu, S., Bell, D., Sternberg, M., & Kutiel, P. (2008). The effect of microhabitats on vegetation and its relationships with seedlings and soil seed bank in a Mediterranean coastal sand dune community. Journal of Arid Environments, 72, 2040-2053. [ Links ]
Wang, R. Z. (2005). Demographic variation and biomass allocation of Agrpyron cristatum grown on steppe and dune sites in the Hunshandake Desert, North China. Grass and forage Science, 60, 99-102. [ Links ]
Zar, J. H. (1996). Bioestatistical Analysis. New Jersey: Prentice Hall. [ Links ]
Zhang, J. J., Wang, L. S., Liu, Z. A., & Li, C. H. (2006). Recent advances in flower color research of tree peony. Acta Horticulturae Sintropical, 33, 1383-1389. [ Links ]
Alcoforado-Filho, F. G., Sampaio, E. V. S. B., & Rodal, M. J. N. (2003). Florística e fitossociologia de um remanescente de vegetação caducifólia espinhosa arbórea em Caruaru. Acta Botânica Brasileira, 17, 287-303. [ Links ]
Andrade, J. R., Santos, J. M. F. F., Lima, E. M., Lopes, C. G. R., Silva, K. A., & Araújo, E. L. (2007). Estudo populacional de Panicum trichoides Swart. (Poaceae) em uma área de Caatinga em Caruaru, Pernambuco. Revista Brasileira de Biociências, 5, 858-860. [ Links ]
Andrade, L. A., Pereira, I. M., Leite, U. T., & Barbosa, M. R. V. (2005). Análise da cobertura de duas fitofisionomias de Caatinga, com diferentes históricos de uso, no Município de São João do Cariri, Estado da Paraíba. Cerne, 11, 253-262. [ Links ]
Araújo, E. L., Silva, K. A., Ferraz, E. M. N., Sampaio, E. V. S. B., & Silva, S. I. (2005). Diversidade de herbáceas em microhabitates rochoso, plano e ciliar em uma área de Caatinga, Caruaru- PE. Acta Botânica Brasílica, 19, 285-294. [ Links ]
Araújo, E. L., & Ferraz, E. M. N. (2003). Processos ecológicos mantenedores da diversidade vegetal na Caatinga: estado atual do conhecimento. In V. Claudino-Sales (Org.), Ecossistemas brasileiros: manejo e conservação. Fortaleza, Brazil: Expressão Gráfica. [ Links ]
Ayres, M., Ayres Júnior, M., Ayres, D. L., & Santos, A. A. (2007). Bioestat-Aplicações estatísticas nas áreas das ciências biomédicas. Belém: Ong Mamiraua. [ Links ]
Belsky, A. J. (1990). Tree/grass ratios in East African savannas: a comparison of existing models. Journal of Biogeography, 17, 483-489. [ Links ]
Belsky, A. J. (1994). Influences of trees on savanna productivity: Tests of shade, nutrients, and tree-grass competition. Journal of Ecology, 75, 922-932. [ Links ]
Brooks, M. L. (1999). Habitat invasibility and dominance by alien annual plants in the western Mojave Desert. Biological Invasions, 1, 325-337. [ Links ]
Castelletti, C. H. M., Santos, A. M. M., Tabarelli, M., & Silva, J. M. C. (2003). Quanto ainda resta da Caatinga? Uma estimativa preliminar. In I. R. Leal, M. Tabarelli, & J. M. C. Silva (Eds.), Ecologia e Conservação da Caatinga. Recife, Brazil: University Publishing UFPE. [ Links ]
Crozier, C. R., & Boerner, R. E. J. (1984). Correlations of Understory Herb Distribution Patterns with Microhabitats under Different Tree Species in a Mixed Mesophytic Forest. Oecologia, 62, 337-343. [ Links ]
Ferguson, B. G., Vandermeer, J., Morales, H., & Griffith, D. M. (2003). Post-agricultural succession in El Péten, Guatemala. Conservation Biology, 17, 818-828. [ Links ]
Flores-Torres, A., & Montaña, C. (2012). Recruiting mechanisms of Cylindropuntia leptocaulis (Cactaceae) in the Southern Chihuahuan Desert. Journal of Arid Environments, 84, 63-70. [ Links ]
García-Cháves, J. H., Montaña, C., Perroni, Y., Sosa, V. J., & García-Licona, J. B. (2014). The relative importance of solar radiation and soil origin in cactos seedling survivorship at two spatial scales: plant association and microhabitat. Journal of Vegetation Science, 25, 668-680. [ Links ]
Gómez-Aparicio, L., Gómez, J. M., & Zamora, R. (2005). Microhabitats shift rank in suitability for seedling establishment depending on habitat type and climate. Journal of Ecology, 93, 1194-1202. [ Links ]
Grouzis, M., & Akpo, L. E. (1997). Influence of tree cover on herbaceous layer above and below ground phytomass in Sahelian zone. Journal of Arid Environment, 35, 285-296. [ Links ]
Hanley, M. E. (1998). Seedling herbivory, community composition and plant life history traits. Perspectives in Plant Ecology, Evolution and Systematics, 1(2), 191-205. [ Links ]
Kessler, J. J. (1992). The influence of karité (Vitellaria paradoxa) and néré (Parkia biglobosa) trees on sorghum production in Burkina Faso. Agroforestry Systems, 17, 297-118. [ Links ]
Köeppen, W. (1948). Climatologia: con un estudio de los climas de la Tierra. México: Fondo de Cultura Económica. [ Links ]
Kostrakiewicz, K. (2009). The influence of shadow created by adjacent plants on phenotypic plasticity of endangered species Trollius europaeus L (Ranunculaceae). Polish Journal of Ecology, 4, 625-634. [ Links ]
Legras, E. C., Vander Wall, S. B., & Board, D. I. (2010). The role of germination microsite in the establishment of sugar pine and Jeffrey pine seedlings. Forest Ecology and Management, 260, 806-813. [ Links ]
Leimu, R. & Lehtila, K. (2006). Effects of two type of herbivores on the population dynamics of a perennial herb. Basic and Applied Ecology, 7, 224-235. [ Links ]
Lopes, C. G. R., Ferraz, E. M. N., Castro, C. C., Lima, E. N., Santos, J. M. F. F., Santos, D. M., & Araújo, E. L. (2012). Forest succession and distance from preserved patches in the Brazilian semiarid region. Forest Ecology Managenment, 271, 115-123. [ Links ]
Lorenzon, M. C. A., Matrangolo, C. A., & Schoereder, J. H. (2003). Flora visitada pelas abelhas eussociais (Hymenoptera, Apidae) na Serra da Capivara, em caatinga do sul do Piauí. Neotropical Entomology, 32, 27-36. [ Links ]
Lozano, A. B., Araújo, E. L., Medeiros, M. F. T., & Albuquerque, U. P. (2014). The apparency hypothesis applied to a local pharmacopoeia in the Brazilian northeast. Journal of Ehtnobiology and Ethnomedicine, 10, 1-17. [ Links ]
Mordelet, P. & Menaut, J. C. (1995). Influence of trees on above-ground production dynamics of grasses in a humid savanna. Journal Vegetable Science, 6, 223-228. [ Links ]
Nordbakken, J. F., Rydgren, K., & Okland, R. H. (2004). Demography and population dynamics of Drosera anglica and D. rotundifolia. Journal of Ecology, 92, 110-121. [ Links ]
Raunkiaer, C. (1934). Life forms of plants and statistical plant geography. Oxford, UK: Clarendon Press. [ Links ]
Reis, A. M. S., Araújo, E. L., Ferraz, E. M. N., & Moura, N. A. (2006). Variações interanuais na florística e fitossociologia do componente herbáceo de uma área de Caatinga, Pernambuco, Brazil. Acta Botânica Brasílica, 29, 497-508. [ Links ]
Riginos, C., Grace, J. B., Augustine, D. J., & Young, T. P. (2009). Local versus Landscape- Scale Effects of Savanna Trees on Grasses. Journal of Ecology, 97, 1337-1345. [ Links ]
Santos, J. M. F. F., Andrade, J. R., Lima, E. M., Silva, K. A., & Araújo, E. L. (2007). Dinâmica populacional de uma espécie herbácea em uma área de floresta tropical seca no Nordeste do Brasil. Revista Brasileira de Biociências, 5, 855-857. [ Links ]
Santos, J. M. F. F., Santos, D. M., Lopes, C. G. R., Silva, K. A., Sampaio, E. V. S. B., & Araújo, E. L. (2013). Natural regeneration of the herbaceous community in a semiarid region in Northeastern Brazil. Environmental Monitoring and Assessment, 185, 8287-8302. [ Links ]
Salo, L. F. (2004). Population dynamics of red brome (Bromus madritensis subsp. rubens): times for concern, opportunities for management. Journal of Arid Evironments, 57, 291-296. [ Links ]
Sampaio, E. V. S. B., Araújo, E. L., Salcedo, I. H., & Tiessen, H. (1998). Regeneração da vegetação após corte e queima, em Serra Talhada, PE. Revista Brasileira de Pesquisa Agropecuária, 33, 621-632. [ Links ]
Silva, A. C. O., Santana, E. F., Saraiva, A. M., Coutinho, F. N., Castro, H. A., Pisciottano, M. N. C., & Albuquerque, U. P. (2013). Which approach is more effective in the selection of plants with antimicrobial activity? Evidence-Based Complementary and Alternative Medicine, 1-9. [ Links ]
Silva, K. A., Lima, E. N., Santos, J. M. F. F., Andrade, J. R., Santos, D. M., Sampaio, E. V. S. B., & Araújo, E. L. (2008). Dinâmica de gramíneas em uma área de Caatinga de Pernambuco-Brasil. In U. P. Albuquerque, A. N. Moura, & E. L. Araújo (Eds.), Biodiversidade, potencial econômico e processos eco fisiológicos em ecossistemas nordestinos. Recife, Brazil: Comunigraf. [ Links ]
Silveira, A. P., Araújo, E. L., Araújo, F. S., & Willadino, L. G. (2005). Predação de frutos e germinação de sementes em Auxemma oncocalyx (Allemão) Baill. e Auxemma glazioviana Taub. In R. J. M. C. Nogueira, E. L. Araújo, L. G. Willadino, & U. M. T. Cavalcante (Eds.), Estresses ambientais: danos e benefícios em plantas. Recife, Brazil: MXM Gráfica e Editora. [ Links ]
Suzuki, Z. O., Kudoh, H., & Kachi, N. (2004). Spatial and temporal variations in mortality of the biennial plant, Lysimachia rubida: effects of intraspecific competition and environmental heterogeneity. Journal of Ecology, 91, 114-125. [ Links ]
Singleton, R., Gardescu, S., Marks, P. L., & Geber, M. A. (2001). Forest herb colonization of postagricultural forests in central New York State, USA. Journal of Ecology, 89, 325-338. [ Links ]
Tews, J., Esther, A., Milton, S. J., & Jeltsh, F. (2006). Linking a population model with an ecosystem model: Assessing the impact of land use and climate change on savanna shrub cover dynamics. Ecological Modelling, 195, 219-228. [ Links ]
Titus, J. H., & Tsuyuzaki, S. (2003). Distribution of plants in relation to microsites on recent volcanic substrates on Mount Koma, Hokkaido, Japan. Ecological Research, 18, 91-98. [ Links ]
Vilá, M., Bartolomeus, I., Gimeno, I., Traveset, A., & Moragues, E. (2006). Demography of the Invasive Geophyte Oxalis pes-caprae Across a Mediterranean Island. Annals of Botany, 97, 1055-1062. [ Links ]
Yu, S., Bell, D., Sternberg, M., & Kutiel, P. (2008). The effect of microhabitats on vegetation and its relationships with seedlings and soil seed bank in a Mediterranean coastal sand dune community. Journal of Arid Environments, 72, 2040-2053. [ Links ]
Wang, R. Z. (2005). Demographic variation and biomass allocation of Agrpyron cristatum grown on steppe and dune sites in the Hunshandake Desert, North China. Grass and forage Science, 60, 99-102. [ Links ]
Zar, J. H. (1996). Bioestatistical Analysis. New Jersey: Prentice Hall. [ Links ]
Zhang, J. J., Wang, L. S., Liu, Z. A., & Li, C. H. (2006). Recent advances in flower color research of tree peony. Acta Horticulturae Sintropical, 33, 1383-1389. [ Links ]
1. Universidade Federal Rural de Pernambuco (UFRPE), Departamento de Biologia, Área Botânica, Laboratório de Ecologia Vegetal dos Ecossistemas Naturais, 52171-900, Dois Irmãos, Recife, Pernambuco, Brazil; julirandrade@yahoo.com.br, enefalcaoo@hotmail.com, danmelo_bio@hotmail.com, elcida@db.ufrpe.br
2. Universidade Federal de Pernambuco (UFPE), Centro Acadêmico de Vitória, Laboratório de Biodiversidade, 55.608-680, Alto do Reservatório, Vitória de Santo Antão, Pernambuco, Brazil; kleberandradedasilva@hotmail.com
3. Universidade Federal Rural de Pernambuco (UFRPE), Departamento de Biologia, Área Ecologia, Laboratório de Ecologia de Peixes, 52171-900, Dois Irmãos, Recife, Pernambuco, Brazil; thiagoguerra@yahoo.com.br
Received 04-VIII-2014. Corrected 17-I-2015. Accepted 12-II-2015.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}