










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.63 n.2 San José Apr./Jun. 2015
What happens to the soil seed bank 17 years after clear cutting of vegetations?
¿Qué ocurre con el banco de semillas del suelo 17 años despúes del corte de la vegetación?
¿Qué ocurre con el banco de semillas del suelo 17 años despúes del corte de la vegetación?
Leonardo Brasil Mendes1*, Kleber Andrade da Silva2*, Danielle Melo dos Santos1, Josiene Maria Falcão Fraga dos Santos1, Ulysses Paulino de Albuquerque1 & Elcida de Lima Araújo1
Abstract
Seed banks play an important role in the resilience of potential anthropogenic areas and are influenced by seasonal variation. The spatial and temporal heterogeneity of the forests influences the richness and density of the soil seed bank, but there is a hypothesis that such influence can be changed in anthropogenic areas, where we expect to find lower richness and seed density in relation to the seed bank of mature forests. The richness and seasonal density of seeds in the soil depth of a young caatinga forest, 17 years after an agricultural activity was abandoned, were evaluated and compared with those of a mature forest. The study was conducted at the Instituto Agronômico de Pernambuco - IPA (Estação Experimental José Nilson de Melo), located in Caruaru, Pernambuco, Brazil, in an area of tropical dry forest, known locally as “caatinga”. The local climate is seasonal, the dry season occurring from September to February and the rainy season concentrated in the remaining months. The average annual rainfall over time (time series of 30 years) is 692mm. In each climatic season (rainy and dry), the seed bank was sampled in 210 20x20cm plots (105 in the leaf litter and 105 at 5cm soil depth). The richness and seed density of the soil samples were evaluated by the method of seedling emergence. The seed bank had 47 species, with a predominance of herbaceous plants. Seasonal variation in richness and seed density in the soil (leaf litter + soil) was not significant, but 42 species of the mature forest were absent from the seed bank of the young forest, despite 17 years of natural regeneration and its proximity to the mature forest. On its own, the soil has greater richness and density of seeds than the leaf litter. The depth of seed deposition in the soil bank of the young forest significantly explained 36% of the species richness and 16% of the seed density, with a significant interaction effect with the climatic season only on species richness, explaining 4% of the variation recorded. The seed density of the young forest (1 277seeds/m2) was greater than that the mature forest indicating that the time abandoned had not yet been sufficient for complete recovery of plant diversity and there is no longer any seed limitation of pioneer species for regeneration of the young forest. Rev. Biol. Trop. 63 (2): 321-332. Epub 2015 June 01.
Key words: semiarid, young forests, savanna, natural regeneration, resilience.
Resumen
El banco de semillas juega un papel importante en el potencial de resiliencia de las áreas antropogénicas y sufre influencia de las variaciones estacionales. La heterogeneidad espacial y temporal de los bosques influye en la riqueza y la densidad del banco de semillas del suelo, pero hay una hipótesis de que tal influencia se puede cambiar en las zonas antropogénicas, donde esperaríamos encontrar menor riqueza y densidad de semillas en relación con el banco de semillas de los bosques maduros. La riqueza y la densidad estacional de semillas del espacio vertical del suelo de un bosque joven, con 17 años de abandono de cualquier actividad agrícola, fueron evaluadas y comparadas con las un bosque maduro. El estudio se realizó en el Instituto Agronômico de Pernambuco - IPA (Estação Experimental José Nilson de Melo), situado en Caruarú, Pernambuco, Brasil, en una zona de bosque seco tropical, conocida localmente como “caatinga”. El clima local es estacional, la estación seca ocurre de septiembre a febrero y la estación de lluvias en los meses restantes. La precipitación promedio anual (series de tiempo de 30 años) es 692mm. En cada estación climática (lluviosa y seca), el banco de semillas fue muestreado en 210 parcelas de 20x20cm (105 en la hojarasca a 5cm de profundidad del suelo). La riqueza y densidad de semillas de las muestras del suelo fueron evaluadas por el método de emergencia de plántulas. El banco de semillas presentó 47 especies, con una predominancia de las herbáceas. Las variaciones estacionales de riqueza y densidad de semillas del suelo (hojarasca+suelo) no fueron significativas, pero 42 especies del bosque maduro todavía estaban ausentes en el banco de semillas del bosque joven, apesar de los 17 años de regeneración natural y de su proximidad con el bosque maduro. Aisladamente, el suelo presentó mayor riqueza y densidad de semillas que la hojarasca. La profundidad de depósito de semillas en el banco del suelo del bosque joven explicó significativamente 36% de la riqueza de especies y 16% de la densidad de semillas, con efecto significativo de interacción con la estación climática apenas sobre la riqueza de las especies, explicando más de 4% de la variación registrada. La densidad de semillas del bosque joven (1 277semillas/m2) fue más elevada que la del bosque maduro indicando, que a pesar del poco tiempo de abandono del cultivo, fue suficiente para la recuperación completa de la densidad de semillas.
Palabras clave: semiárido, bosque joven, caatinga, regeneración natural, resiliencia.
The soil seed bank reflects the factors that affect plant reproduction and survival of seeds in time and space (Donohue, Casas, Burghardt, Kovach, & Willis, 2010; López, 2003; Pennington, Lavin, & Oliveira-Filho, 2009; Pugnaire, & Lázaro, 2000; Silva et al., 2013a), and may be depleted in disturbed areas (Mamede, & Araújo, 2008; Wang, Jiang, Toshio, & Zhou, 2013; Williams, Congdon, Grice, & Clarke, 2005). However, the influence of spatial and temporal heterogeneity on the soil seed bank of areas that have undergone the removal of vegetation is still poorly documented, making it difficult to assess the potential for restoration of anthropogenic forests, the resilience of which, among other factors, depends on the availability of seeds from the soil bank for recruitment (Baeten et al., 2009; Lopes et al., 2012; Wang et al., 2013).
In areas of tropical dry forests with no history of disturbance and with a marked dry season, the dispersion of the propagules is heterogeneous over the months of the year and may vary between years, depending on rainfall totals and the variation in rainfall distribution (Albuquerque et al., 2012; Amorim, Sampaio, & Araujo, 2009; Donohue et al., 2010; Souza, Ferraz, Albuquerque, & Araújo, 2014). Consequently, seasonal differences in richness and density of seed in the soil bank may not occur (López, 2003) or may occur and sometimes be higher either in the dry season (Williams et al., 2005) or in the rainy season (Facelli, Chesson, & Barnes, 2005; Santos et al., 2010), being evidence that precipitation explains 5% of the density of the soil seed bank (Santos et al., 2013a).
Moreover, the spatial heterogeneity (vertical and horizontal) of mature forests also induces differences in the characteristics of the soil seed bank (Pugnaire, & Lázaro, 2000). In some dry forests, the thin layer of leaf litter houses a small amount of seeds when compared to the layers of soil (Costa, & Araújo, 2003; Santos et al., 2010), as the quantity of seeds decreases with increasing soil depth (Williams et al., 2005). The distinct microhabitats of forests are also of relevance for harboring different amount of seeds, explaining 31% of the density of the soil seed bank (Santos et al., 2013a).
Undoubtedly, the spatial and temporal heterogeneity of the forests influences the richness and density of the soil seed bank, but there is a hypothesis that such influence can be changed in anthropogenic areas, where we expect to find lower richness and seed density in relation to the seed bank of mature forests. Since it displays lower richness, we expect higher floristic similarity to occur between climatic seasons in forests at early stages of succession. We also hope that the soil harbors greater richness and seed density when compared to the leaf litter, because in seasonally dry environments, the accumulation of leaf litter tends to be reduced when compared to forests in humid environments (Austin, & Vivanco, 2006), and the decomposition of the leaf litter tends to be faster.
Thus, this study aimed at characterizing the seed bank of an area that suffered clear cutting, by answering the following questions: 1) Are there seasonal and spatial differences in floristic composition, richness and density of the soil seed bank of an area abandoned after clear cutting?, and 2) Are the richness and density of soil seed banks lower in young forests compared to mature forests?
Material and methods
Study area: The study was conducted at the Instituto Agronômico de Pernambuco - IPA (Estação Experimental José Nilson de Melo), located in Caruaru, Pernambuco, Brazil (8o14’18” S - 35o55’20” W, 537masl), in an area of tropical dry forest, known locally as “caatinga”, that suffered clear cutting for the establishment of a culture of Opuntia ficus-indica (L.) Mill. (giant palm), without the use of either fire, fertilizer or an irrigation system. Soon after planting the palm, the area was abandoned and has been regenerating naturally for 17 years, currently holding a young forest in early stages of succession, with few adult individuals (Lopes et al., 2012; Souza et al., 2014).
The portion of the cropped area was part of a mature forest, preserved for more than 50 years, covering about 30ha, and currently there is a small distance of 7m between the young and mature forests, which enables the IPA staff to transit through them. The mature forest has a high richness of woody and herbaceous species, with a predominance of Asteraceae, Euphorbiaceae, Mimosaceae, Poaceae, and Malvaceae (Araújo, Silva, Ferraz, Sampaio, & Silva, 2005a; Reis, Araújo, Ferraz, & Moura, 2006), there being a high similarity between the vegetation and the soil seed bank (Santos et al., 2010; Santos et al., 2013b; Silva et al., 2013a).
The local climate is seasonal, the dry season occurring from September to February and the rainy season concentrated in the remaining months. The average annual rainfall over time (time series of 30 years) is 692mm. The average annual temperature is 22.7oC and the soil of the area is classified as Eutrophic Yellow Podzol (Reis et al., 2006).
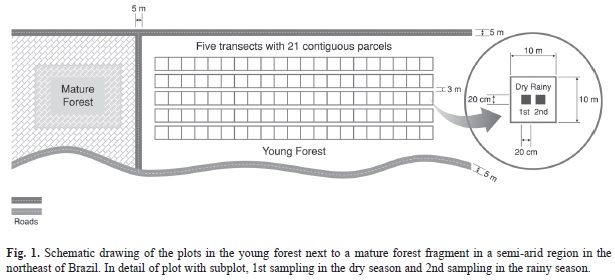
Sampling the seed bank: Five 210m transects were set up in the young forest, parallel to one another and perpendicular to the mature forest. The distance between transects was 3m. In each transect, 21 10x10m contiguous plots were defined, totaling 105 plots (Lopes et al., 2012). In the center of each plot a 20x20cm subplot was defined for the soil sampling (Fig. 1).
To evaluate the influence of spatial and temporal heterogeneity of the soil seed bank, the soil samples were taken at the end of each climatic season (rainy and dry in 2010), considering two levels of seed distribution in the vertical space: 1) leaf litter; 2) soil layer up to 5cm in depth. In total, 210 soil samples (105 in the leaf litter and 105 in the soil) were collected in each season, totaling 420 samples. In the dry season, the sub-plots to collect soil were set up next to the site collected in the rainy season to avoid reduction in the quantity of seeds due to sampling taken beforehand in the rainy season (Santos et al., 2013a; Silva et al., 2013a).
The samples were placed in polystyrene trays, watered daily and kept at room temperature in a greenhouse for six months, simulating the maximum duration of the seedling stage in the caatinga vegetation (Araújo, Martins, & Santos, 2005b). The richness and seed density of the soil samples were evaluated by the method of seedling emergence (Santos et al., 2013a; Thompson, Ceriani, Bakker, & Bekker, 2003), and the seedlings were removed from the tray after identification (Christoffoleti, & Caetano, 1998). A row of tray controls with sterilized soil was set up between the trays to monitor the presence of contamination (Santos et al., 2013a; Silva et al., 2013a). There were no seedlings in the sterilized soil, confirming the absence of contamination.
Species identification was made by comparison with herbarium specimens, from the herbaria Prof. Vasconcelos Sobrinho (PEUFR) and Dárdano de Andrade Lima (IPA), with the aid of specialized literature (Judd, Campbell, Kellogg, & Stevens, 2009) and, when necessary, the material was sent to specialists. Furthermore, when sampling the soil seed bank, seeds of species that were in the reproductive phase were also collected and these were left to germinate in a greenhouse to help identify the species of the seed bank sampled. The unidentified plants were classified by morphospecies (Santos et al., 2013a; Silva et al., 2013a).
The seed density was expressed per square meter. The sample size and method of analysis of the soil seed bank for the analysis of the seed bank of the young forest were the same as those used in previous studies on the seed bank of the mature forest (Santos et al., 2010; Silva et al., 2013a) to enable comparisons between forests.
Analyses of the seed bank: Seasonal differences in richness and average density of seeds in the soil between the leaf litter and 0-5cm soil layer were evaluated through a multiple regression analysis (GLM) with Tukey’s a posteriori test (Zar, 1996). There was no correlation between mean and variance.
To find out if the richness and seed density of the soil seed bank differed between the young forest and the mature forest remnant, the floristic list of the soil seed bank of the mature forest was compiled from a study by Silva et al. (2013a) for three consecutive years and compared to the richness and seed density of the sampled young forest.
The floristic similarity between forests was assessed using the Sorensen similarity index (Zar, 1996), for which three levels of similarity were adopted: 0-33%; 34-66% and 67-100%, considered low, medium and high, respectively. Differences in richness and average seed bank density between forests were verified by the one sample “t” test (Zar, 1996). The analyses were performed using the programs Statistica 7.0 and 5.0 BioEstat.
Results
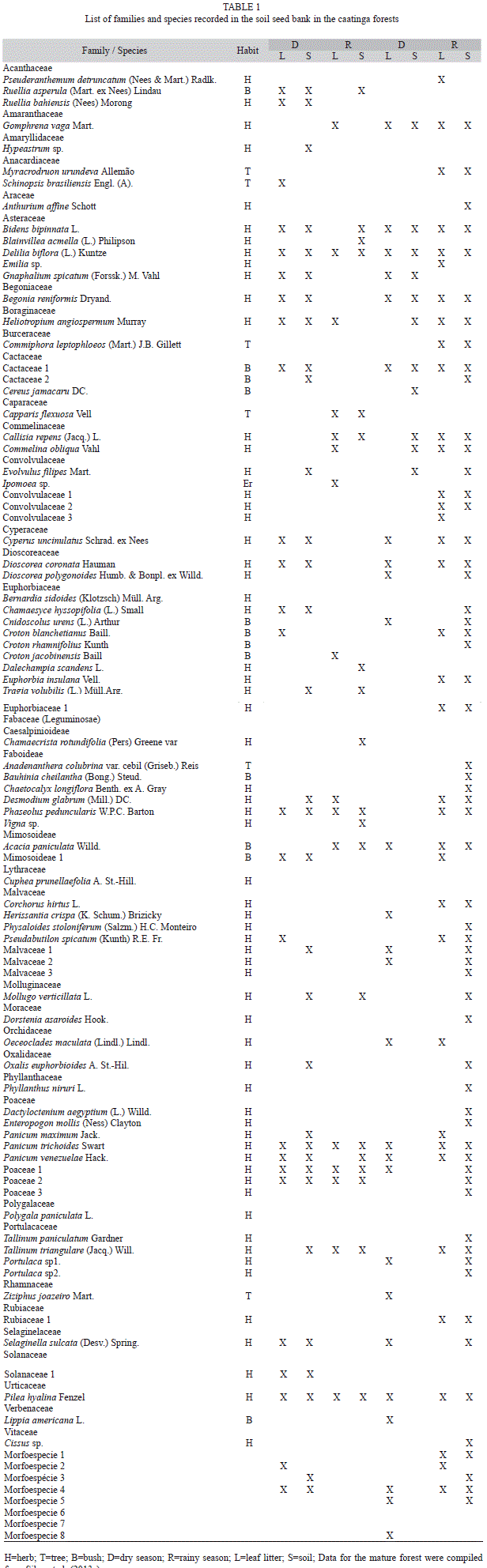
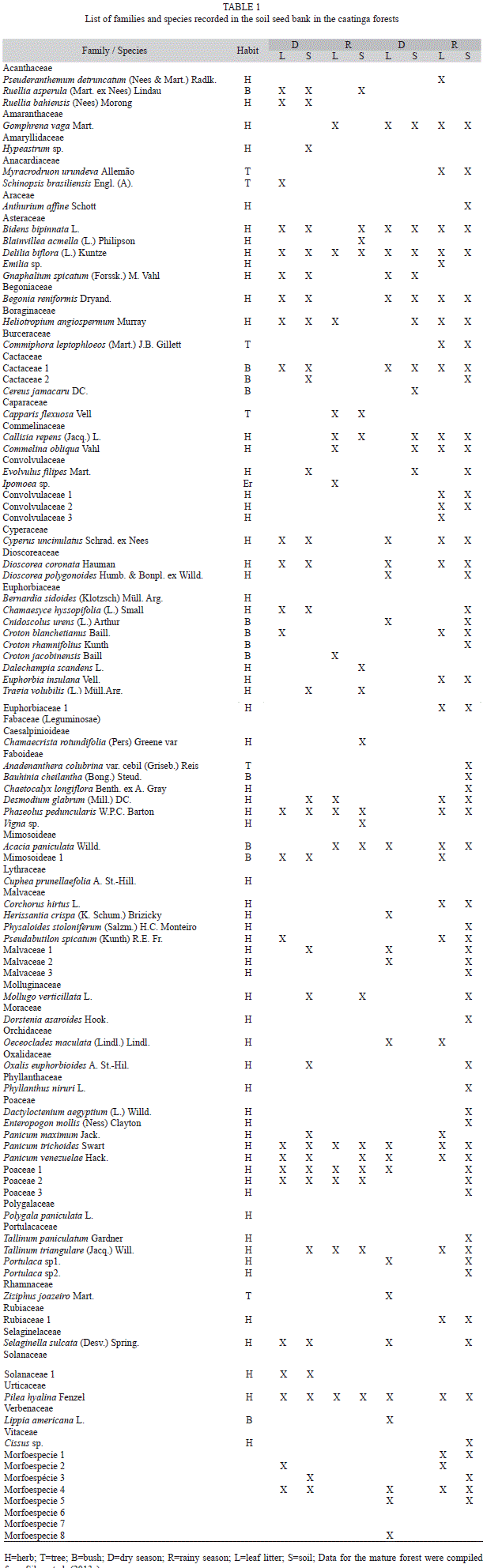
Richness of the soil seed bank of the young forest: A total of 47 species belonging to 24 families were recorded in the seed bank. Of the 47 species, three were identified to the genus level, seven to the family level and three only as morphospecies, due to the precocious death of the seedlings. Excluding the morphospecies, 80% of the species were herbaceous, 4% trees and 16% shrubs. Only one plant without seeds (the fern Selaginella sucata) was recorded in this study, occurring during the dry season. Of the total number of species recorded, 21 occurred only in the dry season, 12 occurred only in the rainy season and 14 species occurred in both seasons.
In the vertical space, considering the two seasons, eight species occurred only in the leaf litter, 13 only in the soil and 26 occurred in both the soil and leaf litter. The floristic similarity ranged from average (57% between seasons) to high (70% between leaf litter and soil). Only the species Delilia biflora, Phaseolus peduncularis, Panicum trichoides, Poaceae sp1, Poaceae sp2 and Pilea hyalina occurred in both seasons, both in the leaf litter and in the soil (Table 1).
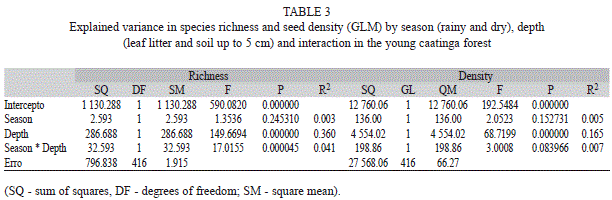
The species richness of the seed bank was significantly higher in the soil in both seasons. Between seasons, the average richness of the seed bank in the leaf litter was significantly higher in the dry season, while in the soil it was significantly higher in the rainy season (Table 2). Combining the leaf litter and the soil, the average richness of the seed bank was similar between seasons. The depth of seed deposition in the soil seed bank significantly explained 36% of the species richness and the interaction between season and depth explained only 4% of the species richness of the soil seed bank (Table 3).
Density of the soil seed bank of the young forest: A total of 1 277 (365 in the leaf litter and 912 in the soil) and 1 038 (101 in the leaf litter and 937 in the soil) seedlings emerged from the seed bank in the dry and rainy season samples, respectively. These values allow us to estimate a density of 304seeds/m2 (87seeds/m2 in the leaf litter and 217seeds/m2 in the soil), in the dry season and 247seeds/m2 (24seeds/m2 in the leaf litter and 223seeds/m2 the soil), in the rainy season.
The mean annual density of the seeds in the soil seed bank in the young forest was 276seeds/m2 (soil + leaf litter). Despite there were no seasonal differences in seed density in soil + leaf litter, it was significantly higher in the soil (Table 2) in both the rainy and dry seasons. Of the total number of seeds recorded in the soil seed bank of the young forest, 16% was significantly explained by the depth of deposition. The relationship between seasons and seed bank density was not significant (Table 3).
Differences in soil seed bank between young and mature forests: A total of 91 species were recorded in the soil seed bank of the young and mature forests (Table 1), with 12 species recorded exclusively in the young forest, 43 only in the mature forest and 36 in both forests. The floristic similarity between forests was average (53%).
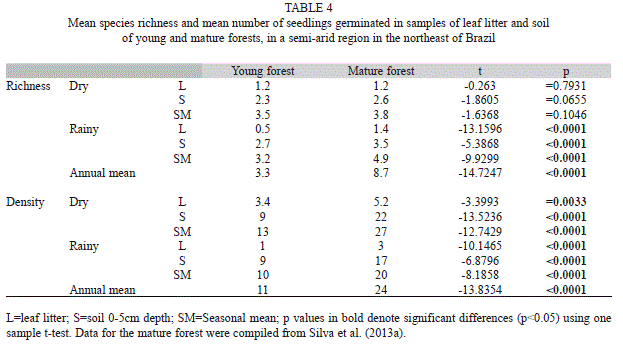
Regardless of the season, the species richness of the seed bank was lower in the young forest. However, separating the seasons it was found that only in the rainy season did the mature forest have the greatest richness in its seed bank. Moreover, the seed bank of the mature forest also harbored a significantly higher number of seeds in the soil and in the leaf litter in both seasons (Table 4).
Discussion
The spatial and seasonal differences in seed density and species richness of the soil seed bank of the young forest showed that spatial and temporal variations in the seed bank in the soil (Facelli et al., 2005; Santos et al., 2010; Santos et al., 2013a; Williams et al., 2005) continue to exist even after 17 years of human intervention. However, changes in the percentage of floristic similarity between the seasons occur, confirming the hypothesis of this study. The percentage (57%) of seasonal similarity was higher than that of mature forests that range from 22% to 53% (Caballero, Olano, Luzuriaga, & Escudero, 2005; López, 2003), possibly due to the influence of seasonality and its interaction with human disturbance on the reproductive period of plants (Albuquerque et al., 2012; Amorim et al., 2009; Donohue et al., 2010; Lima et al., 2007; Lima et al., 2012; Souza et al., 2014).
Lopes et al. (2012) and Souza et al. (2014) conducted studies on seed rain and vegetation structure in the same study area and found that the young caatinga forest was still in early stages of succession and had reduced species richness compared to the mature forest. Thus, perhaps the reduction in seasonal floristic dissimilarity in areas that suffered human disturbance is a reflection of the fact that the area has not yet regained the full floristic richness that existed before the disturbance event.
Allied to this, it is worth noting that some types of vegetation from arid and semiarid environments have high richness of herbaceous species (López, 2003; Reubens, Heyn, Gebrehiwot, Hermy, & Muys, 2007), as was the case of the study area (Santos et al., 2013b), and changes in species composition and assemblage structure may occur naturally depending on interannual variations in rainfall and soil types (Reis et al., 2006; Souza et al., 2014; Silva, Santos, Santos, Ferraz, & Araújo, 2013b). Therefore, it is also possible that, in the year of the study, the species composition of the area was naturally low, explaining the increase in the percentage of similarity recorded and the difference in seasonal richness of the soil depth.
Although there was similarity in richness and seed density (soil + leaf litter) between seasons, as had already been recorded by Silva et al. (2013a) and Lopez (2003), it is still not possible to make further comments on the existence of a seasonal pattern in the characteristics of the seed bank of forests in dry environments as there are studies showing that the richness and density of the seed bank may also be higher in the season that is more favorable (Caballero et al., 2005; Facelli et al., 2005; Kellerman, & Van Rooyen, 2007) or unfavorable (Williams et al., 2005) to seedling emergence.
Regarding the soil depth, some studies of dry forest environments have shown that, regardless of seasonality, density and richness of the seed bank are lower in the leaf litter (Santos et al., 2010; Santos et al., 2013a; Silva et al., 2013a; Yu, Bell, Sternberg, & Kutiel, 2008), as recorded in this study. At least two factors may explain the higher density of seeds in the soil. First, the vegetation of the study area is still in the initial stages of succession and there are many clearings and young trees with poorly developed crowns (Lopes et al., 2012), which naturally reduces the accumulation of leaf litter on the ground.
Second, the deposition of leaf litter in the young forest is higher in the transition between the rainy-dry seasons (Santos, Souza, Santos, Santos, & Araújo, 2011), but decomposition occurs rapidly, justifying the trend for greater richness and seed density in the soil. The high percentage (70%) of floristic similarity between the superficial layer of the soil and the leaf litter indicates that propagules of the few species that are dispersed are quickly incorporated into the soil seed bank.
The seasonal differences in richness and seed density recorded in the soil and leaf litter can be explained by the fact that the dispersal of seeds of many species occurs in the dry season or during the transition between the rainy and dry seasons. Also, the shedding of leaves occurs in response to the arrival of the dry season (Albuquerque et al., 2012; Lima et al., 2012; Souza et al., 2014), which increases the accumulation of leaf litter on the ground, especially in the first months of the dry season (Santos et al., 2011), contributing to greater richness in this season.
In the soil, however, the greater richness and seed density in the rainy season can be explained by the facty that the leaf litter layer is generally shallower (Austin, & Vivanco, 2006; Santos et al., 2011), thus leading to the seeds being incorporated into the soil quickly. Another fact that may help in understanding the greater richness and density of seeds in the soil during the rainy season is the loss of seeds that occurs in the dry season, since rainfall is erratic in the caatinga in that season, triggering the germination of seedlings that end up dying due to that lack of continuous rainfall (Albuquerque et al., 2012; Araújo et al., 2005b; Lima et al., 2007; Santos et al., 2009).
Apart from human disturbance (Mamede, & Araújo, 2008) and the depth of seed deposition in the soil (Yu et al., 2008), there are other variables, such as variation in microhabitats, predation, canopy openness, pathogens and precipitation, that can influence directly or indirectly the richness and density of the soil seed bank (Pugnaire, & Lazaro, 2000; Santos et al., 2010; Santos et al., 2013a; Yan, Zhu, & Gang, 2013). However, although these variables were not monitored in this study, the vertical space alone explained 36% of the variation in species richness and 16% of the variation in density of seeds in the soil bank. Therefore, this study shows that vertical space is a variable of great influence on the seed bank of abandoned agricultural areas and needs to be considered in developing predictive models of richness and seed density for these environments.
However, there are reports that interannual variations of precipitation can explain 48% of the variation in species richness of the seed bank of mature forests (Santos et al., 2013a), indicating that only one year of monitoring is not enough to generalize about a larger pattern of seasonal variation in the richness of the soil seed bank of young forests. Only long-term monitoring will allow us to make larger generalizations.
A significant quantity of species (42) of the soil seed bank of the mature forest were not present in the seed bank of the young forest, despite the contribution of the mature forest in the seed rain of the young forest (Souza et al., 2014), the elapsed time of abandonment (17 years) after agricultural activity, and the proximity between forests (Lopes et al., 2012; Souza et al., 2014).
The average floristic similarity between the seed banks, allied to the occurrence of forest species exclusive to the seed bank of the young forest, show that the seed bank of that forest is depleted by the impact of the action of removal of vegetation, providing a lower species diversity for the restoration of the forests. This confirms the changes in richness and floristic composition that have been recorded in the seed bank of other areas, after anthropic action (Mamede, & Araújo, 2008; Reubens et al., 2007), eventually leading to a reduction of the original plant community to a single population resistant to the disturbance that occurred in the area (Vieira, & Scariot, 2006).
According to Silva et al. (2013a), the density of the soil seed bank of the mature forest may vary between years, since an average of 1 168, 304 and 302seeds/m2 was recorded for three consecutive years. Despite the variation in the density of the soil seed bank of the young forest being as yet unknown, the density recorded in this study (1 277seeds/m2) was higher than that observed by Silva et al. (2013a), indicating that despite the time of abandonment not being sufficient for complete recovery of floristic diversity, there is no seed limitation of pioneer species for regeneration of the young forest.
In the mature forest, spatial variation of microhabitats explained 31% of the seed density of the soil seed bank (Santos et al., 2013a). This percentage, almost double that recorded in the young forest with regard to the explanatory power (16%) of the variable depth of seed deposition, indicates that the young forest may still be impoverished regarding the diversity of microhabitats, and this needs to be further evaluated in future studies.
Acknowledgments
The authors acknowledge the Agronomic Institute of Agricultural Research (IPA) for logistical support; the Federal Rural University of Pernambuco and Postgraduate Program in Ecology; the researchers of the Laboratory of Plant Ecology of Northeast Ecosystems (LEVEN) for their assistance in preparing this manuscript; the CNPq by the scholarship productivity of researchers and the financial support (477239/2009-9); the FACEPE by scholarship.
References
Albuquerque, U. P., Araújo, E. L., Asfora-Eldeir, A. C., Lima, A. L. A., Souto, A., Bezerra, B. M., & Severi, W. (2012). Caatinga Revisited: Ecology and Conservation of an Important Seasonal Dry Forest. The Scientific World Journal, 1-18. [ Links ]
Araújo, E. L., Silva, K. A., Ferraz, E. M. N., Sampaio, E. V. S. B., & Silva, S. I. (2005a). Diversidade de herbáceas em microhabitats rochoso, plano e ciliar em uma área de caatinga, Caruaru, PE, Brasil. Acta Botânica Brasilica, 19(2), 287-296. [ Links ]
Araújo, E. L., Martins, F. R., & Santos, F. A. M. (2005b). Establishment and death of two dry tropical forest woody species in dry and rainy seasons in northeastern Brazil. In R. J. M. C. Nogueira, E. L. Araújo, L. G. Willadino, & U. M. T. Cavalcante (Eds.), Estresses ambientais: danos e benefícios em plantas. Recife, Brazil: MXM Gráfica e Editora. [ Links ]
Amorim, I. L., Sampaio, E. V. S. B., & Araujo, E. L. (2009). Fenologia de Espécies Lenhosas da caatinga do Seridó, RN. Revista Árvore, 33, [ Links ] 491-499.
Austin, A. T., & Vivanco, L. (2006). Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation. Nature, 442(3), 555-558. [ Links ]
Baeten, L., Jacquemyn, H., Calster, H. V., Beek, E. V., Devlaeminck, R., Verheyen, K., & Hermy, M. (2009). Low recruitment across life stages partly accounts for the slow colonization of forest herbs. Journal of Ecology, 97, 109-117. [ Links ]
Caballero, I., Olano, J. M., Luzuriaga, A. L., & Escudero, A. (2005). Spatial coherence between seasonal seed banks in a semi-arid gypsum community: density changes but structure does not. Seed Science Research, 15, 153-160. [ Links ]
Christoffoleti, P. J., & Caetano, R. S. X. (1998). Soil seed bank. Scientia Agricola, 55, 74-78. [ Links ]
Costa, R. C., & Araújo, F. S. (2003). Densidade, germinação e flora do banco de sementes do solo no final da estação seca, em uma área de caatinga, Quixadá, CE. Acta Botanica Brasilica, 17, 259-264. [ Links ]
Donohue, K., Casas, R. R, Burghardt, L., Kovach, K., & Willis, C. G. (2010). Germination, postgermination adaptation, and species ecological ranges. Annual Review of Ecology, Evolution, and Systematics, 10, 293-319. [ Links ]
Facelli, J. M., Chesson P., & Barnes, N. (2005). Differences in seed biology of annual plants in arid lands: a key ingredient of the storage effect. Ecology, 86(11), 2998-3006. [ Links ]
Judd, W. S., Campbell, C. S., Kellogg, E. A., & Stevens, P. F. (2009). Sistemática vegetal: um enfoque filogenético. Porto Alegre: Artmed. [ Links ]
Kellerman, M. J. S., & Van Rooyen, M. W. (2007). Seasonal variation in soil seed bank size and species composition of selected habitat types in Maputaland, South Africa. Bothalia, 37(2), 249-258. [ Links ]
Lima, A. L. A., Sampaio, E. V. S. B., Castro, C. C., Rodal, M. J. N., Antonino, A. C. D., & Melo, A. L. (2012). Do the phenology and functional stem attributes of woody species allow for the identification of functional groups in the semiarid region of Brazil? Trees, 26, 1605-1616. [ Links ]
Lima, E. N., Araújo, E. L., Ferraz, E. M. N., Sampaio, E. V. S. B., Silva, K. A., & Pimentel, R. M. M. (2007). Fenologia e dinâmica de duas populações herbáceas da caatinga. Revista de Geografia, 24, 124-141. [ Links ]
López, R. P. (2003). Soil seed bank in the semi-arid Prepuna of Bolivia. Plant Ecology, 168, 85-92. [ Links ]
Lopes, C. G. R., Ferraz, E. M. N., Castro, C. C., Lima, E. N., Santos, J. M. F. F., Santos, D. M., & Araújo, E. L. (2012). Forest succession and distance from preserved patches in the Brazilian semiarid region. Forest Ecology and Management, 271, 115-123. [ Links ]
Mamede, M. A., & Araújo F. S. (2008). Effects of slash and burn practices on a soil seed bank of Caatinga vegetation in Northeastern Brazil. Journal of Arid Environments, 72, 458-470. [ Links ]
Pennington, R. T., Lavin, M., & Oliveira-Filho, A. (2009). Woody Plant Diversity, Evolution, and Ecology in the Tropics: Perspectives from Seasonally Dry Tropical Forests. Annual Review of Ecology, Evolution, and Systematics, 40, 437-457. [ Links ]
Pugnaire, F. I., & Lazaró, R. (2000). Seed bank and understorey species composition in a semi-arid environment: the effect of shrub age and rainfall. Annals of Botany, 86, 807-813. [ Links ]
Reis, A. M. S., Araújo, E. L., Ferraz, E. M. N., & Moura, A. N. (2006). Inter-annual variations in the floristic and population structure of an herbaceous community of “caatinga” vegetation in Pernambuco, Brazil. Brazilian Journal of Botany, 29(3), 497-508. [ Links ]
Rubens, B., Heyn, M., Gebrehiwot, K., Hermy, M., & Muys, B. (2007). Persistent soil seed banks for natural rehabilitation of dry tropical forests in northern Ethiopia. Tropicultura, 25(4), 204-214. [ Links ]
Santos, J. M. F. F., Silva, K. A., Lima, E. N., Santos, D. M., Pimentel, R. M. M., & Araújo, E. L. (2009). Dinâmica de duas populações herbáceas de uma área de caatinga, Pernambuco, Brasil. Revista de Geografia, 26, 142-160. [ Links ]
Santos, D. M., Silva, K. A., Santos, J. M. F. F., Lopes, C. G. R., Pimentel, R. M. M., & Araújo, E. L. (2010). Variação espaço-temporal do banco de sementes em uma área de floresta tropical seca (caatinga) – Pernambuco. Revista de Geografia, 27(1), 234-253. [ Links ]
Santos, P. S., Souza, J. T., Santos, J. M. F. F., Santos, D. M., & Araújo, E. L. (2011). Diferenças sazonais no aporte de serrapilheira em uma área de caatinga em Pernambuco. Revista Caatinga, 24(4), 94-101. [ Links ]
Santos, D. M., Silva, K. A., Albuquerque, U. P., Santos, J. M. F. F., Lopes, C. G. R., & Araújo, E. L. (2013a). Can spatial variation and inter-annual variation in precipitation explain the seed density and species richness of the germinable soil seed bank in a tropical dry forest in north-eastern Brazil? Flora, 208, 445-452. doi: 10.1016/j.flora.2013.07.006. [ Links ]
Santos, J. M. F. F., Santos, D. M., Lopes, C. G. R., Silva, K. A., Sampaio, E. V. S. B., & Araújo, E. L. (2013b). Natural regeneration of the herbaceous community in a semiarid region in Northeastern Brazil. Environmental Monitoring Assessement, 185, 8287-8302. doi: 10.1007/s10661-013-3173-8. [ Links ]
Silva, K. A., Santos, D. M., Santos, J. M. F. F., Albuquerque, U. P., Ferraz, E. M. N., & Araújo, E. L. (2013a). Spatio-temporal variation in a seed bank of a semi-arid region in northeastern Brazil. Acta Oecologica, 46, 25-32. doi: 10.1016/j.actao.2012.10.008 [ Links ]
Silva, K. A., Santos, J. M. F. F, Santos, D. M., Ferraz E. M. N., & Araújo E. L. (2013b). Spatial variation in the structure and composition of the herbaceous community in a semiarid region of northeastern Brazil. Brazilian Journal of Biology, 73(1), 135-148. doi: http://dx.doi.org/10.1590/S1519-69842013000100015. [ Links ]
Souza, J. T., Ferraz, E. M. N., Albuquerque, U. P., & Araújo, E. L. (2014). Does proximity to a mature forest contribute to the seed rain and recovery of an abandoned agriculture area in a semiarid climate? Plant Biology, 16, 748-756. doi: 10.1111/plb.12120. [ Links ]
Thompson, K., Ceriani, R. M., Bakker, J. P., & Bekker, R. M. (2003). Are seed dormancy and persistence in soil related? Seed Science Research, 13, 97-100. [ Links ]
Vieira, D. L. M., & Scariot, A. (2006). Principles of natural regeneration of tropical dry forests for regeneration. Restoration Ecology, 14, 11-20. [ Links ]
Wang, Y., Jiang, D., Toshio, O., & Zhou, Q. (2013). Recent Advances in Soil Seed Bank Research. Contemporary Problems of Ecology, 6(5), 520-524. doi: 10.1134/S1995425513050181. [ Links ]
Williams, P. R., Congdon, R. A., Grice, A. C., & Clarke, P. J. 2005. Germinable soil seed banks in a tropical savanna: seasonal dynamics and effects of fire. Austral Ecology, 30, 79-90. [ Links ]
Yan, Q., Zhu, J., & Gang, Q. (2013). Comparison of spatial patterns of soil seed banks between larch plantations and adjacent secondary forests in Northeast China: implication for spatial distribution of larch plantations. Trees, 27, 1747-1754. doi: 10.1007/s00468-013-0920-y. [ Links ]
Yu, S., Bell, D., Sternberg, M., & Kutiel, P. (2008). The effect of microhabitats on vegetation and its relationships with seedlings and soil seed bank in a Mediterranean coastal sand dune community. Journal of Arid Environments, 72, 2040-2053. [ Links ]
Zar, J. H. (1996). Bioestatistical analysis. New Jersey: Prentice Hall. [ Links ]
Araújo, E. L., Silva, K. A., Ferraz, E. M. N., Sampaio, E. V. S. B., & Silva, S. I. (2005a). Diversidade de herbáceas em microhabitats rochoso, plano e ciliar em uma área de caatinga, Caruaru, PE, Brasil. Acta Botânica Brasilica, 19(2), 287-296. [ Links ]
Araújo, E. L., Martins, F. R., & Santos, F. A. M. (2005b). Establishment and death of two dry tropical forest woody species in dry and rainy seasons in northeastern Brazil. In R. J. M. C. Nogueira, E. L. Araújo, L. G. Willadino, & U. M. T. Cavalcante (Eds.), Estresses ambientais: danos e benefícios em plantas. Recife, Brazil: MXM Gráfica e Editora. [ Links ]
Amorim, I. L., Sampaio, E. V. S. B., & Araujo, E. L. (2009). Fenologia de Espécies Lenhosas da caatinga do Seridó, RN. Revista Árvore, 33, [ Links ] 491-499.
Austin, A. T., & Vivanco, L. (2006). Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation. Nature, 442(3), 555-558. [ Links ]
Baeten, L., Jacquemyn, H., Calster, H. V., Beek, E. V., Devlaeminck, R., Verheyen, K., & Hermy, M. (2009). Low recruitment across life stages partly accounts for the slow colonization of forest herbs. Journal of Ecology, 97, 109-117. [ Links ]
Caballero, I., Olano, J. M., Luzuriaga, A. L., & Escudero, A. (2005). Spatial coherence between seasonal seed banks in a semi-arid gypsum community: density changes but structure does not. Seed Science Research, 15, 153-160. [ Links ]
Christoffoleti, P. J., & Caetano, R. S. X. (1998). Soil seed bank. Scientia Agricola, 55, 74-78. [ Links ]
Costa, R. C., & Araújo, F. S. (2003). Densidade, germinação e flora do banco de sementes do solo no final da estação seca, em uma área de caatinga, Quixadá, CE. Acta Botanica Brasilica, 17, 259-264. [ Links ]
Donohue, K., Casas, R. R, Burghardt, L., Kovach, K., & Willis, C. G. (2010). Germination, postgermination adaptation, and species ecological ranges. Annual Review of Ecology, Evolution, and Systematics, 10, 293-319. [ Links ]
Facelli, J. M., Chesson P., & Barnes, N. (2005). Differences in seed biology of annual plants in arid lands: a key ingredient of the storage effect. Ecology, 86(11), 2998-3006. [ Links ]
Judd, W. S., Campbell, C. S., Kellogg, E. A., & Stevens, P. F. (2009). Sistemática vegetal: um enfoque filogenético. Porto Alegre: Artmed. [ Links ]
Kellerman, M. J. S., & Van Rooyen, M. W. (2007). Seasonal variation in soil seed bank size and species composition of selected habitat types in Maputaland, South Africa. Bothalia, 37(2), 249-258. [ Links ]
Lima, A. L. A., Sampaio, E. V. S. B., Castro, C. C., Rodal, M. J. N., Antonino, A. C. D., & Melo, A. L. (2012). Do the phenology and functional stem attributes of woody species allow for the identification of functional groups in the semiarid region of Brazil? Trees, 26, 1605-1616. [ Links ]
Lima, E. N., Araújo, E. L., Ferraz, E. M. N., Sampaio, E. V. S. B., Silva, K. A., & Pimentel, R. M. M. (2007). Fenologia e dinâmica de duas populações herbáceas da caatinga. Revista de Geografia, 24, 124-141. [ Links ]
López, R. P. (2003). Soil seed bank in the semi-arid Prepuna of Bolivia. Plant Ecology, 168, 85-92. [ Links ]
Lopes, C. G. R., Ferraz, E. M. N., Castro, C. C., Lima, E. N., Santos, J. M. F. F., Santos, D. M., & Araújo, E. L. (2012). Forest succession and distance from preserved patches in the Brazilian semiarid region. Forest Ecology and Management, 271, 115-123. [ Links ]
Mamede, M. A., & Araújo F. S. (2008). Effects of slash and burn practices on a soil seed bank of Caatinga vegetation in Northeastern Brazil. Journal of Arid Environments, 72, 458-470. [ Links ]
Pennington, R. T., Lavin, M., & Oliveira-Filho, A. (2009). Woody Plant Diversity, Evolution, and Ecology in the Tropics: Perspectives from Seasonally Dry Tropical Forests. Annual Review of Ecology, Evolution, and Systematics, 40, 437-457. [ Links ]
Pugnaire, F. I., & Lazaró, R. (2000). Seed bank and understorey species composition in a semi-arid environment: the effect of shrub age and rainfall. Annals of Botany, 86, 807-813. [ Links ]
Reis, A. M. S., Araújo, E. L., Ferraz, E. M. N., & Moura, A. N. (2006). Inter-annual variations in the floristic and population structure of an herbaceous community of “caatinga” vegetation in Pernambuco, Brazil. Brazilian Journal of Botany, 29(3), 497-508. [ Links ]
Rubens, B., Heyn, M., Gebrehiwot, K., Hermy, M., & Muys, B. (2007). Persistent soil seed banks for natural rehabilitation of dry tropical forests in northern Ethiopia. Tropicultura, 25(4), 204-214. [ Links ]
Santos, J. M. F. F., Silva, K. A., Lima, E. N., Santos, D. M., Pimentel, R. M. M., & Araújo, E. L. (2009). Dinâmica de duas populações herbáceas de uma área de caatinga, Pernambuco, Brasil. Revista de Geografia, 26, 142-160. [ Links ]
Santos, D. M., Silva, K. A., Santos, J. M. F. F., Lopes, C. G. R., Pimentel, R. M. M., & Araújo, E. L. (2010). Variação espaço-temporal do banco de sementes em uma área de floresta tropical seca (caatinga) – Pernambuco. Revista de Geografia, 27(1), 234-253. [ Links ]
Santos, P. S., Souza, J. T., Santos, J. M. F. F., Santos, D. M., & Araújo, E. L. (2011). Diferenças sazonais no aporte de serrapilheira em uma área de caatinga em Pernambuco. Revista Caatinga, 24(4), 94-101. [ Links ]
Santos, D. M., Silva, K. A., Albuquerque, U. P., Santos, J. M. F. F., Lopes, C. G. R., & Araújo, E. L. (2013a). Can spatial variation and inter-annual variation in precipitation explain the seed density and species richness of the germinable soil seed bank in a tropical dry forest in north-eastern Brazil? Flora, 208, 445-452. doi: 10.1016/j.flora.2013.07.006. [ Links ]
Santos, J. M. F. F., Santos, D. M., Lopes, C. G. R., Silva, K. A., Sampaio, E. V. S. B., & Araújo, E. L. (2013b). Natural regeneration of the herbaceous community in a semiarid region in Northeastern Brazil. Environmental Monitoring Assessement, 185, 8287-8302. doi: 10.1007/s10661-013-3173-8. [ Links ]
Silva, K. A., Santos, D. M., Santos, J. M. F. F., Albuquerque, U. P., Ferraz, E. M. N., & Araújo, E. L. (2013a). Spatio-temporal variation in a seed bank of a semi-arid region in northeastern Brazil. Acta Oecologica, 46, 25-32. doi: 10.1016/j.actao.2012.10.008 [ Links ]
Silva, K. A., Santos, J. M. F. F, Santos, D. M., Ferraz E. M. N., & Araújo E. L. (2013b). Spatial variation in the structure and composition of the herbaceous community in a semiarid region of northeastern Brazil. Brazilian Journal of Biology, 73(1), 135-148. doi: http://dx.doi.org/10.1590/S1519-69842013000100015. [ Links ]
Souza, J. T., Ferraz, E. M. N., Albuquerque, U. P., & Araújo, E. L. (2014). Does proximity to a mature forest contribute to the seed rain and recovery of an abandoned agriculture area in a semiarid climate? Plant Biology, 16, 748-756. doi: 10.1111/plb.12120. [ Links ]
Thompson, K., Ceriani, R. M., Bakker, J. P., & Bekker, R. M. (2003). Are seed dormancy and persistence in soil related? Seed Science Research, 13, 97-100. [ Links ]
Vieira, D. L. M., & Scariot, A. (2006). Principles of natural regeneration of tropical dry forests for regeneration. Restoration Ecology, 14, 11-20. [ Links ]
Wang, Y., Jiang, D., Toshio, O., & Zhou, Q. (2013). Recent Advances in Soil Seed Bank Research. Contemporary Problems of Ecology, 6(5), 520-524. doi: 10.1134/S1995425513050181. [ Links ]
Williams, P. R., Congdon, R. A., Grice, A. C., & Clarke, P. J. 2005. Germinable soil seed banks in a tropical savanna: seasonal dynamics and effects of fire. Austral Ecology, 30, 79-90. [ Links ]
Yan, Q., Zhu, J., & Gang, Q. (2013). Comparison of spatial patterns of soil seed banks between larch plantations and adjacent secondary forests in Northeast China: implication for spatial distribution of larch plantations. Trees, 27, 1747-1754. doi: 10.1007/s00468-013-0920-y. [ Links ]
Yu, S., Bell, D., Sternberg, M., & Kutiel, P. (2008). The effect of microhabitats on vegetation and its relationships with seedlings and soil seed bank in a Mediterranean coastal sand dune community. Journal of Arid Environments, 72, 2040-2053. [ Links ]
Zar, J. H. (1996). Bioestatistical analysis. New Jersey: Prentice Hall. [ Links ]
1.Departamento de Biologia. Universidade Federal Rural de Pernambuco. Dois Irmãos. C.P. 52171-900. Brasil. Recife, Pernambuco; lbrasilmendes@yahoo.com.br, danmelo_bio@hotmail.com, enefalcaoo@hotmail.com, upa@db.ufrpe.br, elcida@db.ufrpe.br
2. Centro Acadêmico de Vitória. Universidade Federal de Pernambuco. Bela Vista. C.P. 55608-680. Brasil. Vitória de Santo Antão, Pernambuco; kleberandradedasilva@hotmail.com
Received 06-VI-2014. Corrected 30-X-2014. Accepted 26-XI-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}