










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.63 n.1 San José Jan./Mar. 2015
Parámetros de crecimiento y mortalidad de Orthopristis ruber (Perciformes: Haemulidae) en el archipiélago Los Frailes, Venezuela
Growth and mortality parameters of Orthopristis ruber (Perciformes: Haemulidae) from Los Frailes Archipielago, Venezuela
Growth and mortality parameters of Orthopristis ruber (Perciformes: Haemulidae) from Los Frailes Archipielago, Venezuela
Abstract
Orthopristis ruber is a species in high demand in Eastern Venezuela, but production has been decreasing in recent years. For this reason, our objective was to estimate the growth and mortality parameters of this resource. Monthly samples were collected from June 2011 to May 2012, obtaining 2 980 specimens in El Tirano and Puerto Abajo. Data on total length (cm), total weight (g), and sex were recorded for each specimen. No sexual dimorphism was shown with respect to length (ts=1.113, p>0.05), so one length-weight ratio was established for both sexes (Wt=0.0612*Lt2.54); and they both exhibited minor allometric growth. Growth was estimated by analysis of length frequency distributions using FiSAT software. The estimated growth parameters (L∞=39.03cm, W∞=679.60g, k=0.48/year and to=-0.32year) showed moderately rapid growth. Length frequency data were adjusted to the von Bertalanffy model, and indicated an exponential tendency of accelerated growth during the first years of life, followed by slow growth until the fish reached its maximum length. The coefficient of variation of the growth index (Ø’) demonstrated no differences in growth pattern. The natural mortality rate (M=0.97/year), from fishing (F=1.57/year), and total mortality (Z=2.54/year), were high, as well as the exploitation rate (E=0.62/year). We concluded that O. ruber has been fully exploited by artisanal fishers, and suggest a continuous study on population dynamics, to recommend optimum management techniques for the fishery.
Key words: growth, mortality, population parameters, Orthopristis ruber, Los Frailes Archipelago, Venezuela.
Resumen
Orthopristis ruber es una especie de gran demanda en el oriente de Venezuela cuya producción se ha visto disminuida en los últimos años. Por esta razón, se planteó como objetivo estimar los parámetros de crecimiento y mortalidad del recurso. Se realizaron muestreos mensuales de junio 2011 a mayo 2012, para un total de 2 980 ejemplares recolectados en El Tirano y Puerto Abajo. A cada ejemplar se le tomó datos de longitud total (cm), peso total (g) y se determinó el sexo. Se comprobó que no existe diferencia sexual con respecto a longitud (ts=-1.113; p>0.05), la relación longitud-peso para ambos sexos fue: Pt=0.0612*Lt2.54, lo que evidencia un crecimiento alométrico minorante. La estimación del crecimiento se basó en el análisis de la distribución de frecuencia de longitudes, con uso del software FiSAT. Los parámetros de crecimiento estimados (L∞=39.03cm, P∞=679.60g, k=0.48/año y to=-0.32año) mostraron un crecimiento moderadamente rápido. Los datos de frecuencias de longitudes fueron ajustados al modelo de von Bertalanffy, e indica una tendencia de tipo exponencial con crecimiento acelerado los primeros años de edad, que luego se hizo lento hasta que el pez alcanzó la longitud máxima. El coeficiente de variación del índice de crecimiento (Ø’) demostró que no hay diferencias en el patrón de crecimiento. La tasa de mortalidad natural (M=0.97/año), por pesca (F=1.57/año), y total (Z=2.54/año) fueron altas así como la tasa de explotación (E=0.62/año). Se concluye que O. ruber está plenamente explotado por la pesca artesanal; por lo que se sugiere continuar con los estudios de dinámica de poblaciones para evaluar la pesquería con fines de manejo.
Palabras clave: crecimiento, mortalidad, parámetros poblacionales, Orthopristis ruber, archipiélago Los Frailes, Venezuela.
Los estudios de crecimiento de peces son esenciales para la mayor parte de los objetivos de la evaluación pesquera, tanto en las regiones tropicales como en las templadas, debido a que el crecimiento individual de los organismos constituye, a través del tiempo, la fuente de suministro de las capturas extraídas por una pesquería (Pauly, 1983). En pesquerías la mortalidad total es interpretada como la disminución en el número de individuos de una cohorte a causa de diversos factores. Los coeficientes de mortalidad que normalmente se usan en el estudio de la dinámica de las poblaciones están relacionados con las dos principales causas de mortalidad a que están expuestos los peces a lo largo de su vida: la mortalidad natural, que depende de las características biológicas de cada stock y de las cohortes que la componen, y la mortalidad por pesca, causada por el hombre, que se utiliza en la administración pesquera (Sparre & Venema, 1997). La mortalidad natural puede considerarse siempre como un factor constante que afecta por igual y en forma permanente a todos los peces, mientras que la mortalidad por pesca no siempre puede ser considerada como una constante, ya que en gran parte está sujeta a los deseos o habilidades del hombre (Csirke, 1980); de tal manera, la pesca actúa sobre la población como un factor de mortalidad más, que se suma a las otras causas de mortalidad y hace que las clases anuales disminuyan rápidamente, y también a la población.
En el Atlántico Centro Occidental las especies de la familia Haemulidae están entre los peces más capturados por la pesca deportiva y pesca artesanal de subsistencia, siendo las especies más numerosas en los arrecifes tropicales del Atlántico (Courtenay & Sahlman, 1978; Darcy, 1983; Rasotto & Sadovy, 1995; Lindeman, Diaz, Serafy, & Aula, 1998). Dentro de las pesquerías artesanales multiespecíficas que se desarrollan en el oriente de Venezuela, Orthopristis ruber (Cuvier, 1830) ocupa un lugar importante en las capturas comerciales y tiene una gran demanda, sobre todo por familias de escasos recursos económicos (Eslava, 1988). Se captura con nasa, cordel y redes todo el año sin ninguna medida de manejo (González, Eslava, & Guevara, 2006), y en el 2012, representó el 0.45% (514t) de la producción pesquera marítima venezolana (115 414t), según datos aportados por el Instituto Socialista de la Pesca y Acuicultura (INSOPESCA), lo que indica su importancia en las capturas comerciales para el consumo en fresco.
En Venezuela, se han realizado varios estudios de biología y dinámica poblacional de O. ruber, tales como los de reproducción (Carvajal, 1973), alimentación (Gil & Bastardo, 1975; González, 1981), relación longitud-peso (Etchevers, 1975; Parra, 1996), isópodos parásitos (Eslava, 1988), índices ecológicos y parámetros biométricos (Parra, Ruiz, & Prieto, 2007), distribución, abundancia y parámetros pesqueros (Acosta, 2011), que no permiten evaluar el tamaño de la población explotable. En tal sentido, se planteó estimar los parámetros de crecimiento y mortalidad de O. ruber del archipiélago Los Frailes, Venezuela, con la finalidad de contribuir con la ordenación del recurso, cuya captura se ha visto disminuida en los últimos años al pasar de 3 480t en 2003 a 514t en 2012 según el INSOPESCA.
Materiales y Métodos
Los ejemplares de O. ruber procedieron de la captura artesanal con nasa por los pescadores de la comunidad de El Tirano que faenan en el archipiélago Los Frailes de lunes a sábado seis horas diarias, desde las 03:00 (hora de zarpe) hasta las 09:00 (hora de llegada al puerto de desembarque), en seguida comercializan la pesca en fresco directamente al consumidor y en algunos casos a intermediarios. El archipiélago Los Frailes está ubicado al noreste de la isla de Margarita del estado Nueva Esparta está constituido por diez islotes de 192ha² (1.92km²) de superficie (11º11’30” N - 63º42’00” W) (Fig. 1), siendo la de mayor tamaño Fraile Grande o Puerto Real que tiene una longitud máxima de 2 200m con una superficie de 0.75km² y forman parte de las Dependencias Federales de Venezuela (Villa, 1967; Cervigón, 1995). Los pescadores realizan faenas diarias. Los muestreos se realizaron mensualmente en las playas El Tirano y Puerto Abajo, de junio 2011 a mayo 2012. El número de individuos muestreados por mes oscilaron entre 48 y 419. A cada ejemplar se le tomó la longitud total (Lt), peso total (Pt) y la identificación del sexo se realizó mediante la observación de las gónadas según Billings y Munro (1974).
Para determinar diferencias significativas entre las longitudes por sexo, se realizó la prueba de ts (Zar, 2009). Se determinó la relación longitud-peso mediante la ecuación propuesta por Ricker (1975): Pt=a*Ltb; donde Pt es el peso total sin eviscerar en g, Lt es la longitud total en cm, a es la intersección con el eje de las ordenadas y b es la pendiente de la curva. Las constantes “a” y “b” fueron estimadas por el método de los mínimos cuadrados, previa linearización de los datos mediante una transformación logarítmica (ln). Para determinar la existencia de diferencias significativas entre los coeficientes de machos y hembras (a y b), se aplicó la hipótesis del crecimiento isométrico (Ricker, 1975) mediante una prueba ts (Walpole, Myers, Myers, & Ye, 2007) con un nivel de significación de α=0.05.
La estimación del crecimiento se basó en el análisis de la distribución de frecuencia de longitudes. Para ello se usó el software FiSAT (Gayanilo, Sparre, & Pauly, 1996). En primer lugar, se estimó una longitud asintótica (L∞) aplicando el método de Powell (1979) y Wetherall (1986), que permitió determinar el coeficiente de crecimiento (k) a través de la rutina ELEFAN I; para la optimización de los parámetros de L∞ y de k se empleó el procedimiento de Gulland y Holt (1959) asociado en la misma rutina, previo análisis de la progresión modal utilizando para ello la descomposición de frecuencia de longitudes según el Método de Bhattacharya (1967). En segundo lugar, se calculó el to de acuerdo a la fórmula anotada por Pauly (1979): Log10 (-to)=-0.3922–0.2752*Log10 L∞ -1.038*Log10 k; donde to es la edad teórica en que el organismo tiene talla cero, L∞ es la longitud asintótica promedio máximo y k es el coeficiente de crecimiento o coeficiente de curvatura. Para el cálculo de la longevidad se utilizó la ecuación de Taylor (1958): A0.95=to+2.996/k; donde A0.95 es la edad límite o tiempo requerido para alcanzar el 95% de la longitud máxima del pez (L∞). El peso asintótico se estimó mediante la fórmula anotada por Csirke (1980): P∞=a*L∞b; donde P∞ es el peso asintótico o peso máximo promedio. Una vez ajustados L∞, P∞, k y to, se determinaron las curvas de crecimiento en longitud y en peso adecuadas al modelo de von Bertalanffy (1938), según las siguientes ecuaciones: Lt=L∞[1-e–k(t-to)]; Pt =P∞[1-e–k(t-to)]b. Las curvas de crecimiento, de longitud fueron comparadas a través de un análisis de varianza de doble vía con un nivel de significación de 0.05 (Zar, 2009). Los parámetros de crecimiento estimados de O. ruber, fueron comparados con resultados obtenidos por otros investigadores en diferentes regiones, mediante el índice phi prima (Ø’) de Munro y Pauly (1983): Ø’=Log10k+2Log10L∞ y el criterio de Sparre y Venema (1997), quienes establecen que el coeficiente de variación (CV) no debe exceder del 4% para poder asumir que los resultados son estadísticamente similares.
De la curva de captura linearizada se derivaron los valores de mortalidad total aparente (Z), la mortalidad por pesca (F) y la mortalidad natural (M), con el fin de estimar la tasa de explotación según la ecuación anotada por Cushing (1981): E=F/Z*(1–e–Z). La mortalidad natural (M) se evaluó a través de la ecuación de Pauly (1980): Log10 M=-0.0066–0.279*Log10 L∞+0.6543*Log10 k+0.4632*Log10 T, utilizando una temperatura (T) media anual del área de 25ºC. La mortalidad por pesca (F) se calculó de la diferencia entre Z y M.
Resultados
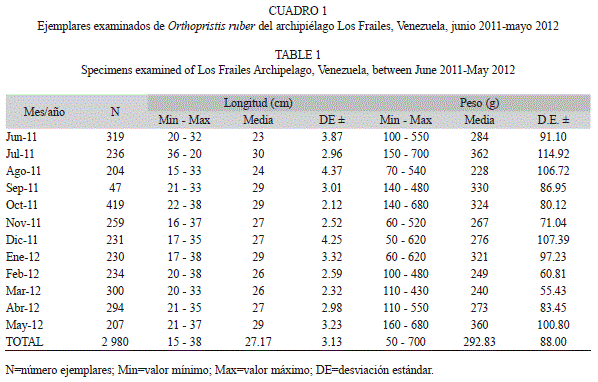
Se examinaron un total de 2 980 individuos, con longitudes comprendidas de 15 a 38cm con un promedio de 27.17cm (±3.13), con pesos que oscilaron entre 50 y 700g con un promedio de 292.83g (±88); el menor número de organismos se recolectó en el septiembre 2011 porque hubo poca pesca, debido a las festividades religiosas (Virgen del Valle), mientras que el mayor fue registrado en octubre 2011 (Cuadro 1). No se encontraron diferencias estadísticamente significativas para la longitud entre hembras y machos (α=95% y 99 grados de libertad, ts=1.113; p>0.05), por lo que se trataron los sexos combinados, para la determinación de crecimiento y de la mortalidad. Durante el período junio 2010-mayo 2011, las longitudes con mayor frecuencia estuvieron comprendidas entre los 19 y 30cm, siendo el de mayor abundancia 29cm, observándose la mayor abundancia en marzo y octubre que corresponden a las época de surgencia y post surgencia (relajación).
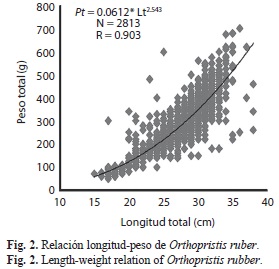
Crecimiento: La relación longitud-peso quedó establecida como Pt=0.0612*Lt2.54 siendo la constante “b” significativamente diferente a 3 (ts=20.045; p<0.05), demostrando un crecimiento alométrico minorante (Fig. 2), lo que indica que los ejemplares son menos pesados para la longitud que tienen.
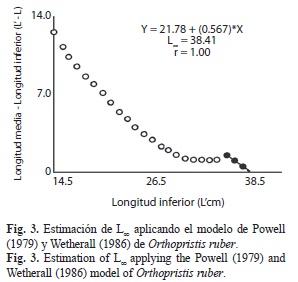
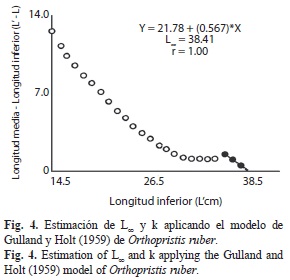
Se obtuvo una primera estimación de la longitud asintótica (L∞) igual a 38.41cm (Fig. 3) la cual fue utilizada como dato de entrada en el ELEFAN I, obteniendo una primera estimación de k=0.35/año, y luego de varias corridas con la superficie de respuesta con la rutina ELEFAN I, se determinó que la combinación que presentó el mejor ajuste (Rn=0.160) fue L∞=39.03cm y k=0.48/año. Posteriormente, se aplicó la rutina de Gulland y Holt (1959) la cual proporcionó valores adicionales de L∞ y k, los que fueron descartados debido a que el valor de k estimado no responde a las características biológicas de la especie (Fig. 4). Se estimó el to=-0.32años lo que permitió finalmente determinar la A0.95 ≈ 7años.
Con la frecuencia de tallas reestructuradas con el ELEFAN I, se pudo observar que el crecimiento de O. ruber es continuo durante todo el año. El peso asintótico (P∞=679.60g) se obtuvo utilizando las constantes “a” y “b” de la ecuación de la relación longitud-peso y se reemplazó la variable Lt por el parámetro L∞ obtenido en la rutina ELEFAN I.
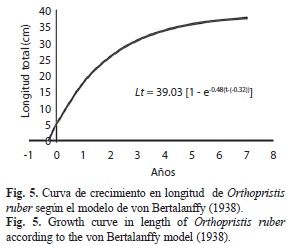
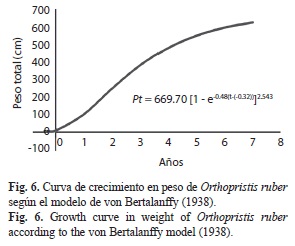
La curva de crecimiento en longitud describió una curva de tipo exponencial, acelerado en el primer año de vida, alcanzando aproximadamente el 47% de su longitud asintótica, y luego disminuyó lentamente hasta hacerse casi constante hasta llegar a la longitud máxima: Lt=39.03[1-e–0.48(t+0.32)] (Fig. 5); mientras que la curva de crecimiento en peso demostró un comportamiento sigmoideo, lento en el primer año de vida, y luego, lo hace en forma exponencial hasta la edad 3, para finalmente alcanzar un punto en la cual la curva se inflexiona, en este punto la especie va aumentando su peso hasta alcanzar su peso asintótico: Pt=679.60[1-e–0.48 (t +0.32)]2.543 (Fig. 6).
La comparación de los parámetros de crecimiento de la ecuación de von Bertalanffy y el coeficiente de variación (CV) se presentan en el Cuadro 2.
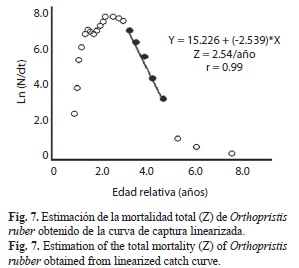
Mortalidad: La mortalidad natural establecida fue M=0.97/año y la mortalidad total fue Z=2.54/año (Fig. 7). Con las mortalidades natural y total estimadas, se pudo determinar la mortalidad por pesca F=1.57/año y la tasa de explotación E=0.62/año.
Discusión
Las longitudes mínimas y máximas encontradas en esta especie están dentro del rango de 12 y 52cm para especies de la familia Haemulidae (Billings & Munro, 1974). La ausencia de longitudes menores reportadas durante este estudio pueden deberse, probablemente, a la selectividad de la nasa, arte de pesca utilizado por los pescadores, y la disponibilidad del recurso en el área de pesca tal como lo señala Bravo, Eslava, y González (2009). Es factible que los juveniles se encuentren en zonas de aguas tranquilas y someras, siendo las mismas, áreas de crianza; en relación a esto, Lindeman (1989) señala que los juveniles se dirigen a los estuarios en busca de albergue así como para el desarrollo de las gónadas, permaneciendo fuera del área de pesca hasta alcanzar cierta talla en la cual se reclutan al stock de adultos. No se observó diferencia estadísticamente significativa entre las longitudes de hembras y machos, resultado que concuerda con González (1981) y Parra et al. (2007).
La relación longitud-peso de O. ruber indica un crecimiento alométrico minorante, similar al determinado por Etchevers (1975) y Acosta (2011), en el Golfo de Cariaco, Venezuela y a los reportados por Haimovici y Velasco (2000) en el sur de Brasil para especies de la familia Haemulidae, pero difiere con lo reportado por Parra et al. (2007), en el sur de Margarita, Venezuela, quienes determinaron un crecimiento isométrico, y a lo estimado por Vianna y Verani (2002) y do Santos, Pessanha y da Costa (2004) en aguas brasileñas, quienes establecieron un crecimiento alométrico mayorante. Las diferencias observadas responden a la tasa de crecimiento en peso, que puede variar en peces de diferentes localidades, sexos, edades y fases de crecimiento (Weatherley, 1972).
La longitud asintótica estimada para O. ruber fue diferente a la encontrada por Acosta (2011) (L∞=11.52cm) en aguas del Golfo de Cariaco, y similar a lo reportado por Vianna y Verani (2002), quienes aseguran que en O. ruber en aguas brasileñas, puede crecer hasta 41cm. Las diferencias en la longitud asintótica pueden deberse a que Acosta (2011) trabajó con una alta proporción de juveniles (talla máxima 17.1cm); mientras que en este estudio y en el realizado por Vianna y Verani (2002) se colectaron ejemplares que en su mayoría fueron adultos. El coeficiente de crecimiento k reportado en este trabajo fue mayor al estimado por Vianna y Verani (2002), y menor a lo reportado por Acosta (2011), lo que indica que este es un organismo que posee un crecimiento moderado. A menudo las especies pelágicas son más activas que las demersales, y poseen valores de k más altos (Sparre & Venema, 1997). La fracción juvenil menor a 15cm no apareció regularmente en las capturas comerciales y esto podría ser la causa principal de la subestimación del coeficiente de crecimiento k debido a la correlación estadística que existe entre los parámetros de crecimiento. Björn (2005), señala que las especies de la familia Haemulidae, a lo largo de su vida, experimentan una serie de migraciones a las áreas de reclutamiento asociadas a los cambios en la dieta y comportamiento alimenticio, aparentemente, correlacionadas con la longitud del pez, es decir, mientras los más pequeños se alimentan del plancton, los más grandes comienzan a alimentarse de algunos otros organismos marinos, y a medida que van creciendo, migran hacia aguas más profundas agregándose a los arrecifes coralinos, y a las áreas de pesca. González (1981) demostró que existen variaciones de la cantidad de los componentes alimentarios a través del año y talla de los individuos de O. ruber. Las diferencias de crecimiento de una especie, en diferentes regiones, posiblemente, se deben a factores ambientales, oceanográficos y al tipo de fondo que controla la distribución de la fauna González, Eslava, y Silva (1998).
La diferencia de los parámetros de crecimiento obtenidas en distintas áreas geográficas puede deberse, a diferencias genéticas y ambientales (temperatura del agua, disponibilidad de alimento, y niveles de explotación, entre otros.). El valor del índice de crecimiento (Ø’) estimado fue similar al obtenido por Vianna y Verani (2002), pero diferente con el de Acosta (2011); sin embargo, el coeficiente de variación sugiere que son estadísticamente similares el O. ruber de Brasil y Venezuela y presentan el mismo patrón de crecimiento.
El valor de la mortalidad natural de O. ruber estimado (M=0.97/año) fue parecido al determinado por Acosta (2011) en aguas del Golfo de Cariaco (M=1.01/año), pero superior a la encontrada por Vianna y Verani (2002) en aguas brasileñas (M=0.73/año). Los hemúlidos tienen una alta tasa de mortalidad natural, causada principalmente por depredación, enfermedad y vejez; y una alta tasa de mortalidad por pesca, la que actúa sobre una población haciendo descender el número de supervivientes (Billings & Munrro, 1974). Los individuos de una misma especie en diferentes áreas pueden tener diversas causas de mortalidad natural, una de ellas es la temperatura del agua de las regiones tropicales que aceleran los procesos biológicos provocando elevadas mortalidades (Pauly, 1979), otra es la cantidad de depredadores y competidores, cuya abundancia está influenciada por la pesca (Sparre & Venema, 1997).
La mortalidad total determinada para O. ruber fue menor a lo reportado por Acosta (2011), quien estimó una alta tasa mortalidad total (Z=4.324/año), y mayor a la calculada por Vianna y Verani (2002) para esta especie en aguas brasileñas (Z=2.46/año). Valores de mortalidad, menores a los estimados en otras regiones, puede suponer que la misma no es tan alta, lo que indica una condición saludable para la especie. Una alta tasa de mortalidad total, también, sugiere un fuerte impacto de la pesca, cuyo efecto directo puede causar cambios en la abundancia, en relación a un ecosistema equilibrado, y también en la estructura de tallas, con una reducción en las tallas máximas (Sparre & Venema, 1997).
El valor de la tasa de explotación obtenido en el presente trabajo fue alto (E=0.62/año) similar para aguas brasileñas (E=0.70/año) estimado por Vianna y Verani (2002), por lo que se establece que esta especie está sobre explotada. La tasa de explotación es un parámetro que permite conocer cuál es la situación de un stock respecto a la presión de pesca. Cada especie tiene una tasa de explotación máxima en donde no hay interferencia con la reproducción. Pasado el límite los mecanismos compensatorios no permiten que la población se recupere (Tresierra & Culquichicón, 1993). Por otro lado, Gulland (1971) menciona que la tasa de explotación óptima (E óptima) se da cuando F=M, de manera general se considera que E óptima ≈ 0.5. En tal sentido, estos resultados preliminares permiten proponer medidas de ordenación, como vedas basadas en el comportamiento reproductivo o cuotas de captura.
Se concluye que O. ruber no evidencia dimorfismo sexual con respecto a la longitud y presenta un crecimiento moderadamente rápido con una alta tasa de explotación lo cual demuestra que el recurso está plenamente explotado por la pesca artesanal; por lo que se sugiere continuar con los estudios de dinámica de poblaciones para evaluar la pesquería con fines de manejo.
Agradecimientos
A Juan Miguel Rodríguez, Presidente de la Cooperativa Virgen del Carmen de El Tirano y a José Gregorio Arcay del camión cava “Bonanza” por el apoyo logístico en el muestreo, facilitando los ejemplares antes de su comercialización. A Clark Casler por la lectura y sugerencias al manuscrito. Al Consejo de Investigación de la Universidad de Oriente por el financiamiento parcial del Proyecto de Grupo: “Evaluación de la pesquería artesanal de la comunidad de El Tirano, isla de Margarita” Código CI-6-030603-1587/2009.
Referencias
Acosta, J. (2011). Distribución, abundancia y parámetros poblacionales de cuatro especies de peces de la familia Haemulidae en el Golfo de Cariaco, Venezuela (Tesis de Licenciatura). Universidad de Oriente, Cumaná, Venezuela. Recuperado de http://ri.biblioteca.udo.edu.ve/bitstream/123456789/2440/1/TESIS_JA.pdf. [ Links ]
Bhattacharya, C. (1967). A simple method of resolution of a distribution into Gaussian components. Biometrics, 23, 115-135. [ Links ]
Billings, C., & Munro, J. (1974). The biology, ecology and bionomics of Caribbean reef fishes: Pomadasydae (Grunts). Part 5. Research Report from the Zoology Department, University of the West Indies, 3, 1-128. [ Links ]
Björn, K. (2005). Ontogenetic migration and growth of French grunt (Teleostei: Haemulon flavolineatum) as determined by coded wire tags (Tesis de Maestría). Universidad de Puerto Rico, Mayagüez, Puerto Rico. Recuperado de http://grad.uprm.edu/oeg/ENGLISH/ThesesDissertations/CienciasMarinas/#2005. [ Links ]
Bravo, E., Eslava, N., & González, L. W. (2009). Crecimiento y mortalidad del pez Haemulon aurolineatum (Teleostei: Haemulidae) en el suroeste de la isla de Margarita, Venezuela. Revista de Biología Tropical, 57(1-2), 105-112. [ Links ]
Carvajal, J. (1973). Capturas comerciales y reproducción de Orthopristis ruber al Nor-este de la isla de Margarita, Venezuela. Boletín del Instituto Oceanográfico de Venezuela Universidad de Oriente, 12(1), 33-40. [ Links ]
Cervigón, F. (1995). Las Dependencias Federales. Caracas, Venezuela: Academia Nacional de la Historia. [ Links ]
Courtenay, W., & Sahlman, H. (1978). Pomadasyidae. Vol. 4, unpaginated. In W. Fischer (Ed.), FAO species identification sheets for fishery purposes, Western Central Atlantic (Fishing Area 31). Rome: FAO. [ Links ]
Csirke, J. (1980). Introducción a la dinámica de poblaciones de peces (Documento Técnico de Pesca No. 192). Roma: FAO. [ Links ]
Cushing, D. H. (1981). Fisheries biology, a study in population dynamics. Madison, USA: Univ. Wisconsin System. [ Links ]
Darcy, G. H. (1983). Synopsis of the biological data on the pigfish, Orthopristis crysoptera (Pisces: Haemulidae) (NOAA Technical Report NMSF, FAO Fisheries, USA Synopsis, 134). USA: FAO. [ Links ]
do Santos, A., Pessanha, A., da Costa, M., & Araújo, F. (2004). Relação peso-comprimento de Orthopristis ruber (Cuvier, 1830) (Teleostei, Haemulidae) na Baia de Sepetiba, Rio de Janeiro, Brasil. Revista Brasilera de Zoología, 21(2), 185-187. [ Links ]
Eslava, N. (1988). Ocurrencia de isópodos parásitos en el Corocoro, Orthopristis ruber (Cuvier, 1830) (Pisces: Haemulidae). Memoria de la Sociedad de Ciencias Naturales La Salle, 48(129), 85-92. [ Links ]
Etchevers, S. (1975). La relación longitud-peso en 7 especies de interés comercial en el nororiente de Venezuela. Boletín del Instituto Oceanográfico de Venezuela Universidad de Oriente, 14(2), 243-246. [ Links ]
Gayanilo, F., Sparre, P., & Pauly, D. (1996). The FAO-ICLARM Stock Assessment Tools (FiSAT) User’s Manual. FAO Computerized Information Series (Fisheries) No. 8. Rome: FAO. [ Links ]
Gil, T., & Bastardo, J. (1975). Investigación de enterobacterias en el contenido gastrointestinal de corocoro, Orthopristis ruber (Pisces: Perciformes, Pomadasyidae) [contaminación de agua, Venezuela]. Boletín del Instituto Oceanográfico de Venezuela Universidad de Oriente, 14(2), 149-156. [ Links ]
González, L. W. (1981). Régimen alimentario del corocoro, Orthopristis ruber (Cuvier, 1830) (Pisces: Pomadasydae) en las zonas adyacentes a la isla de Margarita, Venezuela. Boletín del Instituto Oceanográfico de Venezuela Universidad de Oriente, 20(1-2), 23-32. [ Links ]
González, L. W., Eslava, N., & Silva, C. (1998). Edad, crecimiento y mortalidad de Lutjanus purpureus Poey, 1867 (Pisces: Lutjanidae) de la región de Guayana. Boletín Investigaciones Marinas y Costeras, 27, 7-20. [ Links ]
González, L. W., Eslava, N., & Guevara, F. (2001). Análisis de la pesca artesanal del pargo gallo (Lachnolaimus maximus) (Teleostei: Labridae) en el archipiélago Los Frailes, Venezuela. Boletín del Centro de Investigaciones Biológicas, 35(3), 272-282. [ Links ]
González, L. W., Eslava, N., & Guevara, F. (2006). Catálogo de la pesca artesanal del estado Nueva Esparta, Venezuela. Cumaná, Venezuela: Editoriales Radoca, C. A. [ Links ]
Gulland, J. (1971). Manual de métodos para la evaluación de las poblaciones de peces. Zaragoza, España: FAO. [ Links ]
Gulland, J., & Holt, S. (1959). Estimation of growth parameters for data at unequal time intervals. Journal Conseil International pour L´Exploitation de la Mer, 25, 47-49. [ Links ]
Haimovici, M., & Velasco, G. (2000). Length-weight relationships of marine fish from southern Brazil. Naga, Worldfish Center Quarterly, 23(1), 19-23. [ Links ]
Lindeman, K. (1989). Coastal construction, larval settlement, and early juvenile habitat use in grunts, snappers and other coastal fishes of southeast Florida. Bulletin of Marine Science, 44(2), 1068-1089. [ Links ]
Lindeman, K., Diaz, G., Serafy, J., & Aula, J. (1998). A spatial framework for assessing cross-shelf habitat use among newly settled grunts and snappers. Proceedings Gulf Caribbean Fisheries Institute, 50, 385-416. [ Links ]
Munro, J., & Pauly, D. (1983). A simple method for comparing the growth of fishes and invertebrates. Fishbyte, 1(1), 5-6. [ Links ]
Parra, B. (1996). Relación longitud-peso, factor de condición e índice hepático de Haemulon aurolineatum (Cuvier, 1830) (Pisces: Haemulonidae) de la isla de Cubagua, Venezuela. Acta Científica de Venezuela, 47(1), 122-134. [ Links ]
Parra, B., Ruiz, L., & Prieto, A. (2007). Índices ecológicos y parámetros biométricos de Haemulidae (Pisces: Perciformes) en la zona costera de la Isla de Cubagua, Venezuela. Zootecnia Tropical, 25(1), 51-61. [ Links ]
Pauly, D. (1979). Theory and management of tropical multispecies stocks: a review, with emphasis on the Southeast Asian demersal fisheries. Studies and Reviews No 1. Manila, Filipinas: ICLARM. [ Links ]
Pauly, D. (1980). On the interrelationships between mortality growth parameters and mean environmental temperature in 175 fish stocks. Journal Conseil International pour L´Exploitation de la Mer, 39, 175-192. [ Links ]
Pauly, D. (1983). Algunos métodos simples para la evaluación de recursos pesqueros tropicales (Fishering Technical Paper, 234). FAO. [ Links ]
Powell, D. (1979). Estimation of mortality and growth parameters for the length frequency of a catch. Raport process–V. Réunion Conseil International pour L´Exploitation de la Mer, 175, 167-169. [ Links ]
Rasotto, M., & Sadovy, Y. (1995). Peculiarities of the male urogenital apparatus of two grunt species (Teleostei: Haemulidae). Journal Fisheries Biology, 46, 936-948. [ Links ]
Ricker, W. (1975). Computation and interpretation of biological statistic of fish population. Department of Environmental Fisheries and Marine Service. Bulletin of the Fisheries Researchs Board of Canada, 191: 382. [ Links ]
Sparre, P., & Venema, S. (1997). Introducción a la evaluación de recursos pesqueros tropicales (Parte 1. Manual. Documento Técnico de Pesca Nº 306. 1 Revisión 2. 420 pp.). Roma: FAO. [ Links ]
Taylor, C. (1984). Cod growth and temperature. Journal Conseil International pour L´Exploitation de la Mer, 23, 366-370. [ Links ]
Tresierra, A., & Culquichicón, Z. (1993). Biología Pesquera. Trujillo, Perú: Editorial Libertad. [ Links ]
Vianna, M., & Verani, J. R. (2002). Biologia reprodutiva de Orthopristis ruber (Teleostei, Haemulidae) espécie acompanhante da pesca de arrasto do camarão-rosa, no sudeste Brasileiro. Atlántica, 23(1), 27-36. [ Links ]
Villa, M. (1967). Aspectos geográficos de las Dependencias Federales. Caracas: Corporación Venezolana de Fomento. [ Links ]
von Bertalanffy, L. (1938). A quantitative theory of organic growth. Human Biology, 10(2), 181-213. [ Links ]
Walpole, R., Myers, R., Myers, S., & Ye, K. (2007). Probability statistics for engineer scientists. 8th Edition. New Jersey, USA: Prentice Hall. [ Links ]
Weatherley, A. (1972). Growth and ecology of fish populations. London: Academic Press. [ Links ]
Wetherall, J. (1986). A new method for estimating growth and mortality parameters from length-frequency data. Fishbyte, 4, 12-14. [ Links ]
Zar, J. (2009). Biostatistical Analysis. New Jersey, USA: Prentice Hall. [ Links ]
Bhattacharya, C. (1967). A simple method of resolution of a distribution into Gaussian components. Biometrics, 23, 115-135. [ Links ]
Billings, C., & Munro, J. (1974). The biology, ecology and bionomics of Caribbean reef fishes: Pomadasydae (Grunts). Part 5. Research Report from the Zoology Department, University of the West Indies, 3, 1-128. [ Links ]
Björn, K. (2005). Ontogenetic migration and growth of French grunt (Teleostei: Haemulon flavolineatum) as determined by coded wire tags (Tesis de Maestría). Universidad de Puerto Rico, Mayagüez, Puerto Rico. Recuperado de http://grad.uprm.edu/oeg/ENGLISH/ThesesDissertations/CienciasMarinas/#2005. [ Links ]
Bravo, E., Eslava, N., & González, L. W. (2009). Crecimiento y mortalidad del pez Haemulon aurolineatum (Teleostei: Haemulidae) en el suroeste de la isla de Margarita, Venezuela. Revista de Biología Tropical, 57(1-2), 105-112. [ Links ]
Carvajal, J. (1973). Capturas comerciales y reproducción de Orthopristis ruber al Nor-este de la isla de Margarita, Venezuela. Boletín del Instituto Oceanográfico de Venezuela Universidad de Oriente, 12(1), 33-40. [ Links ]
Cervigón, F. (1995). Las Dependencias Federales. Caracas, Venezuela: Academia Nacional de la Historia. [ Links ]
Courtenay, W., & Sahlman, H. (1978). Pomadasyidae. Vol. 4, unpaginated. In W. Fischer (Ed.), FAO species identification sheets for fishery purposes, Western Central Atlantic (Fishing Area 31). Rome: FAO. [ Links ]
Csirke, J. (1980). Introducción a la dinámica de poblaciones de peces (Documento Técnico de Pesca No. 192). Roma: FAO. [ Links ]
Cushing, D. H. (1981). Fisheries biology, a study in population dynamics. Madison, USA: Univ. Wisconsin System. [ Links ]
Darcy, G. H. (1983). Synopsis of the biological data on the pigfish, Orthopristis crysoptera (Pisces: Haemulidae) (NOAA Technical Report NMSF, FAO Fisheries, USA Synopsis, 134). USA: FAO. [ Links ]
do Santos, A., Pessanha, A., da Costa, M., & Araújo, F. (2004). Relação peso-comprimento de Orthopristis ruber (Cuvier, 1830) (Teleostei, Haemulidae) na Baia de Sepetiba, Rio de Janeiro, Brasil. Revista Brasilera de Zoología, 21(2), 185-187. [ Links ]
Eslava, N. (1988). Ocurrencia de isópodos parásitos en el Corocoro, Orthopristis ruber (Cuvier, 1830) (Pisces: Haemulidae). Memoria de la Sociedad de Ciencias Naturales La Salle, 48(129), 85-92. [ Links ]
Etchevers, S. (1975). La relación longitud-peso en 7 especies de interés comercial en el nororiente de Venezuela. Boletín del Instituto Oceanográfico de Venezuela Universidad de Oriente, 14(2), 243-246. [ Links ]
Gayanilo, F., Sparre, P., & Pauly, D. (1996). The FAO-ICLARM Stock Assessment Tools (FiSAT) User’s Manual. FAO Computerized Information Series (Fisheries) No. 8. Rome: FAO. [ Links ]
Gil, T., & Bastardo, J. (1975). Investigación de enterobacterias en el contenido gastrointestinal de corocoro, Orthopristis ruber (Pisces: Perciformes, Pomadasyidae) [contaminación de agua, Venezuela]. Boletín del Instituto Oceanográfico de Venezuela Universidad de Oriente, 14(2), 149-156. [ Links ]
González, L. W. (1981). Régimen alimentario del corocoro, Orthopristis ruber (Cuvier, 1830) (Pisces: Pomadasydae) en las zonas adyacentes a la isla de Margarita, Venezuela. Boletín del Instituto Oceanográfico de Venezuela Universidad de Oriente, 20(1-2), 23-32. [ Links ]
González, L. W., Eslava, N., & Silva, C. (1998). Edad, crecimiento y mortalidad de Lutjanus purpureus Poey, 1867 (Pisces: Lutjanidae) de la región de Guayana. Boletín Investigaciones Marinas y Costeras, 27, 7-20. [ Links ]
González, L. W., Eslava, N., & Guevara, F. (2001). Análisis de la pesca artesanal del pargo gallo (Lachnolaimus maximus) (Teleostei: Labridae) en el archipiélago Los Frailes, Venezuela. Boletín del Centro de Investigaciones Biológicas, 35(3), 272-282. [ Links ]
González, L. W., Eslava, N., & Guevara, F. (2006). Catálogo de la pesca artesanal del estado Nueva Esparta, Venezuela. Cumaná, Venezuela: Editoriales Radoca, C. A. [ Links ]
Gulland, J. (1971). Manual de métodos para la evaluación de las poblaciones de peces. Zaragoza, España: FAO. [ Links ]
Gulland, J., & Holt, S. (1959). Estimation of growth parameters for data at unequal time intervals. Journal Conseil International pour L´Exploitation de la Mer, 25, 47-49. [ Links ]
Haimovici, M., & Velasco, G. (2000). Length-weight relationships of marine fish from southern Brazil. Naga, Worldfish Center Quarterly, 23(1), 19-23. [ Links ]
Lindeman, K. (1989). Coastal construction, larval settlement, and early juvenile habitat use in grunts, snappers and other coastal fishes of southeast Florida. Bulletin of Marine Science, 44(2), 1068-1089. [ Links ]
Lindeman, K., Diaz, G., Serafy, J., & Aula, J. (1998). A spatial framework for assessing cross-shelf habitat use among newly settled grunts and snappers. Proceedings Gulf Caribbean Fisheries Institute, 50, 385-416. [ Links ]
Munro, J., & Pauly, D. (1983). A simple method for comparing the growth of fishes and invertebrates. Fishbyte, 1(1), 5-6. [ Links ]
Parra, B. (1996). Relación longitud-peso, factor de condición e índice hepático de Haemulon aurolineatum (Cuvier, 1830) (Pisces: Haemulonidae) de la isla de Cubagua, Venezuela. Acta Científica de Venezuela, 47(1), 122-134. [ Links ]
Parra, B., Ruiz, L., & Prieto, A. (2007). Índices ecológicos y parámetros biométricos de Haemulidae (Pisces: Perciformes) en la zona costera de la Isla de Cubagua, Venezuela. Zootecnia Tropical, 25(1), 51-61. [ Links ]
Pauly, D. (1979). Theory and management of tropical multispecies stocks: a review, with emphasis on the Southeast Asian demersal fisheries. Studies and Reviews No 1. Manila, Filipinas: ICLARM. [ Links ]
Pauly, D. (1980). On the interrelationships between mortality growth parameters and mean environmental temperature in 175 fish stocks. Journal Conseil International pour L´Exploitation de la Mer, 39, 175-192. [ Links ]
Pauly, D. (1983). Algunos métodos simples para la evaluación de recursos pesqueros tropicales (Fishering Technical Paper, 234). FAO. [ Links ]
Powell, D. (1979). Estimation of mortality and growth parameters for the length frequency of a catch. Raport process–V. Réunion Conseil International pour L´Exploitation de la Mer, 175, 167-169. [ Links ]
Rasotto, M., & Sadovy, Y. (1995). Peculiarities of the male urogenital apparatus of two grunt species (Teleostei: Haemulidae). Journal Fisheries Biology, 46, 936-948. [ Links ]
Ricker, W. (1975). Computation and interpretation of biological statistic of fish population. Department of Environmental Fisheries and Marine Service. Bulletin of the Fisheries Researchs Board of Canada, 191: 382. [ Links ]
Sparre, P., & Venema, S. (1997). Introducción a la evaluación de recursos pesqueros tropicales (Parte 1. Manual. Documento Técnico de Pesca Nº 306. 1 Revisión 2. 420 pp.). Roma: FAO. [ Links ]
Taylor, C. (1984). Cod growth and temperature. Journal Conseil International pour L´Exploitation de la Mer, 23, 366-370. [ Links ]
Tresierra, A., & Culquichicón, Z. (1993). Biología Pesquera. Trujillo, Perú: Editorial Libertad. [ Links ]
Vianna, M., & Verani, J. R. (2002). Biologia reprodutiva de Orthopristis ruber (Teleostei, Haemulidae) espécie acompanhante da pesca de arrasto do camarão-rosa, no sudeste Brasileiro. Atlántica, 23(1), 27-36. [ Links ]
Villa, M. (1967). Aspectos geográficos de las Dependencias Federales. Caracas: Corporación Venezolana de Fomento. [ Links ]
von Bertalanffy, L. (1938). A quantitative theory of organic growth. Human Biology, 10(2), 181-213. [ Links ]
Walpole, R., Myers, R., Myers, S., & Ye, K. (2007). Probability statistics for engineer scientists. 8th Edition. New Jersey, USA: Prentice Hall. [ Links ]
Weatherley, A. (1972). Growth and ecology of fish populations. London: Academic Press. [ Links ]
Wetherall, J. (1986). A new method for estimating growth and mortality parameters from length-frequency data. Fishbyte, 4, 12-14. [ Links ]
Zar, J. (2009). Biostatistical Analysis. New Jersey, USA: Prentice Hall. [ Links ]
1. Escuela de Ciencias Aplicadas del Mar de la Universidad de Oriente. Boca del Río, Isla de Margarita, Venezuela; atenaszarai@gmail.com
2. Instituto de Investigaciones Científicas de la Universidad de Oriente. Boca del Río, Isla de Margarita, Venezuela; neslava20@yahoo.es, lwgc25@gmail.com, franciscogm2002e@hotmail.com
Recibido 12-III-2014. Corregido 20-IX-2014. Aceptado 22-X-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}