










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.1 San José Feb. 2014
Fish community structure on coral habitats with contrasting architecture in the Tropical Eastern Pacific
Abstract
Reefscape architecture, shaped by dominant coral morphologies, can play a major role in determining the structure and composition of fish assemblages by affecting niche and resource availability and mediating interspecific interactions. To explore the role of dominant coral morphologies on reef fish communities, we carried out a comparative study of the fish community associated with a Massive Coral Community (MCC) and a Branching Coral Community (BCC) at Gorgona Island, Tropical Eastern Pacific (TEP). On each community, the benthic substrate was assessed through the “chain transect method” while the fish assemblage was evaluated through visual surveys on belt transects. We found differences between both fish assemblages in terms of the abundance, diversity (H’), and evenness (J’). The BCC, despite being formed by morphologically complex pocilloporid colonies, had a simple and relatively flat architecture that attracted principally small and territorial fishes. Significant higher abundances of Chromis atrilobata and Thalassoma lucasanum at the BCC boosted the total fish abundance but caused low fish evenness and diversity. Conversely the MCC, composed of massive coral species with considerable sizes and diversity of shapes, held a complex and high-relief reefscape capable of sustaining a more diverse and even fish community, although with the same species richness as the BCC. Fishes with large sizes, roving behavior and piscivore-feeding preferences were especially attracted to the MCC. Although, massive coral species are important in determining a diverse and complex reefscape architecture, both dominant coral morphologies (massive and branching) attract and provide resources to different types of fish according to their size, mobility and trophic group. Our results suggest that a loss of massive coral species and a community shift towards stress-resistant taxa (such as Pocillopora spp.), could alter the structure and function of fish assemblages in the TEP due to the habitat loss for large, mobile and piscivore species. Rev. Biol. Trop. 62 (Suppl. 1): 343-357. Epub 2014 February 01.
Key words: coral reef fishes, reefscape architecture, coral morphology, massive coral community, branching coral community, Tropical Eastern Pacific.
Resumen
La arquitectura del paisaje arrecifal, definida por la morfología de los corales dominantes, puede desempeñar un papel importante en la estructura y composición de las comunidades de peces al afectar la disponibilidad de nichos y recursos y modificar las interacciones interespecíficas. Hicimos un estudio comparativo entre la comunidad de peces asociada a una comunidad de corales masivos (CCM) y a una de corales ramificados (CCR) en la isla Gorgona, Pacífico Oriental Tropical. En cada formación coralina, el sustrato bentónico se evaluó a través de “transectos de cadena”, mientras que la comunidad de peces se valoró con el uso censos visuales en transectos de banda. Hubo diferencias en la abundancia, diversidad (H’) y equitatividad (J’) de las dos comunidades de peces. La CCR, a pesar de estar formada por colonias morfológicamente complejas de corales pocillóporidos, presentó una arquitectura simple y relativamente plana que atrajo principalmente peces territoriales y de talla pequeña. Abundancias significativamente altas de Chromis atrilobata y Thalassoma lucasanum en la CCR, aumentaron la abundancia total de peces, pero ocasionaron una baja diversidad y equitatividad de la comunidad. Por el contrario, la CCM constituida principalmente por especies de corales masivos con diversos tamaños y formas, presentó una arquitectura compleja y de alto relieve capaz de mantener una comunidad de peces mucho más diversa y equitativa, aunque con la misma riqueza de especies de peces que la CCR. Los peces de gran talla, con comportamiento errante y hábitos carnívoros fueron atraídos a la MCC. En general, nuestro estudio evidenció que aunque las especies de coral con crecimiento masivo son importantes en la formación de una arquitectura compleja, cada una de las morfologías de coral dominante (masivo y ramificado) atrae y brinda recursos a distintos grupos de peces según su tamaño y grupo trófico. La pérdida de corales masivos o un cambio en la comunidad hacia corales pocilóporidos (resistentes al estrés) podrían alterar la diversidad y función comunitaria de peces en el Pacífico Oriental Tropical (POT) debido a la pérdida de hábitat para las especies grandes, móviles y carnívoras.
Palabras clave: peces de arrecifes de coral, arquitectura del arrecife, morfología de corales, comunidad de corales masivos, comunidad de corales ramificados, Pacífico Oriental Tropical.
Habitat complexity can be a major determinant of community structure because it strongly influences the availability of niches and the ways in which species exploit environmental resources (Hutchinson, 1959; MacArthur & MacArthur, 1961; Tews et al., 2004). Although a strong positive relationship between habitat complexity and species diversity was first observed in terrestrial ecosystems (reviewed by Tews et al., 2004), many studies have also demonstrated strong linkages between properties of fish assemblages and the complexity of marine habitats (e.g., Risk, 1972; Luckhurst & Luckhurst, 1978; Friedlander & Parrish, 1998; Gratwicke & Speight, 2005; Alvarez-Filip, Gill & Dulvy, 2011a).
Tropical coral reefs, bio-constructed by scleractinian coral species, are among the most complex marine ecosystems due to their coral diversity, architectural complexity, variety of refuge sizes, percentage of live cover and vertical relief (Luckhurst & Luckhurst, 1978; Roberts & Ormond, 1987; Friedlander & Parrish, 1998; Gratwicke & Speight, 2005; Pratchett et al., 2008; Messmer et al., 2011). Among these habitat features, architectural complexity (defined here as the structure of the physical environment) usually plays the most important role in shaping the richness, diversity, abundance and biomass of reef fish assemblages (Luckhurst & Luckhurst, 1978; McCormick, 1994; Almany, 2004; Alvarez-Filip et al., 2011a). It is believed that a high structural complexity boosts fish diversity because it reduces interspecific competition by providing a greater spectrum of niches and resources. It also increases fish survivorship by diminishing prey-predator encounter rates and/or by offering additional prey refuges (Beukers & Jones, 1997; Friedlander & Parrish, 1998; Almany, 2004; Gratwicke & Speight, 2005).
The importance of dominant coral morphologies (e.g., tabular, massive, branching) has been highlighted in recent years, because it is an important coral reef attribute that shapes the reefscape architectural complexity and determines habitat usage by reef fishes (Alvarez-Filip et al., 2011a; Messmer et al., 2011; Kerry & Bellwood, 2012). For example, branching coral communities with many interstices are often used by small reef fishes for shelter (Beukers & Jones, 1997), while tabular corals congregate larger reef fishes that seek concealment for ambush predation or shade from sunlight (Kerry & Bellwood, 2012). Studying fish-habitat associations also provides insights into the effects of decreased structural complexity as a consequence of habitat degradation (Pratchett et al., 2008; Benfield, Baxter, Guzman & Mair, 2008; Alvarez-Filip, Dulvy, Côté, Watkinson & Gill, 2011b). Coral morphology is, however, a measure of architectural complexity usually not considered in coral reef fish studies, as most methods designed to assess the habitat complexity of coral communities (Risk, 1972; McCormick 1994; Wilson, Graham & Polunin, 2007; Wedding, Friedlander, McGranaghan, Yost & Monaco, 2008) involve the use of quantitative indices that conceal the role of dominant coral species and their associated morphological and functional traits (Luckhurst & Luckhurst, 1978; Jones & Syms, 1998; Kerry & Bellwood, 2012).
Although reef fish communities from the Tropical Eastern Pacific (TEP) have received increasing attention in the last decade (e.g., Edgar, Barrett & Morton, 2004; Alvarez-Filip, Reyes-Bonilla & Calderon-Aguilera, 2006; Dominici-Arosemena & Wolff, 2006; Benfield et al., 2008), few studies have considered coral morphology (massive or branching) as an important factor influencing habitat structure and thus the composition and structure of fish assemblages in the region (Zapata & Morales, 1997; Dominici-Arosemena & Wolff, 2006; Benfield et al., 2008). For instance, Zapata & Morales (1997) noted that changes in fish richness and evenness among reef zones within a fringing coral reef responded to the relative distribution of massive and branching coral types. As the dominant coral forms of a coral community determine substrate monotony and complexity, they appear to be strongly correlated with the richness and diversity of fish assemblages (Dominici-Arosemena & Wolff, 2006).
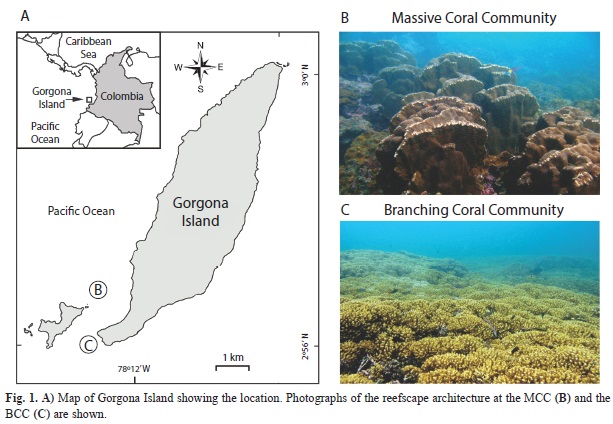
In this study, we examined how dominant coral morphologies influence fish community structure and composition by carrying out a comparative study of the fish assemblages on two coral formations with contrasting reefscape architecture. One formation was dominated by branching coral species, which formed a monotonous framework of low relief, while the other was composed of large massive coral colonies that made a high-relief, heterogeneous and complex seascape (Fig. 1). Considering a large spatial scale (the gross reefscape morphology) we addressed the following questions: (1) Are there differences in the fish assemblages inhabiting habitats dominated by branching and massive corals?, (2) which reefscape pattern or habitat architecture features the highest fish species richness, diversity, and abundance?, and (3) which types of fish species and trophic groups are most attracted to each of the two coral communities assessed?
Matherials and methods
Study area: Located in the Panamic province at 2°58’27”N - 78°11’13”W, Gorgona is a continental island 35 km off the Pacific coast of Colombia (Fig. 1A). Despite the adverse climatic and oceanographic conditions for coral reef development in the TEP (e.g., upwelling, El Niño events; Glynn & Ault, 2000), Gorgona holds more than 30ha of coral communities and reefs that regularly exhibit dense coral growth (80% live coral cover on reef crests; Zapata & Vargas-Ángel, 2003). The Massive Coral Community (MCC) considered here, is the largest grouping of massive and submassive coral colonies found in the island (~8ha), holding species such as Gardineroseris planulata, Pavona clavus, P. varians, P. frondifera and Porites lobata, which can reach considerable sizes (often 1m and occasionally =2m high and =3m in diameter) and occur at an average depth of 8m (Fig. 1B). The MCC is located in the south-western, windward side of the island in an area locally known as “La Camaronera”, which is characterized by strong waves and low light penetration due to high turbidity and sediment re-suspension (Zapata & Vargas-Ángel, 2003). In contrast, the Branching Coral Community (BCC) we studied lies on the south-eastern, leeward side of Gorgona (Fig. 1C) at a site known as “La Ventana”. It was first described by Glynn, Prahl & Guhl (1982) as a dense and continuous stand of Pocillopora spp. colonies which formed buildups of approximately 1m thick. The coral framework is shallow (~5m depth) and although it is often subjected to strong currents (Prahl & Erhardt, 1985), its waters are clearer than at the MCC site.
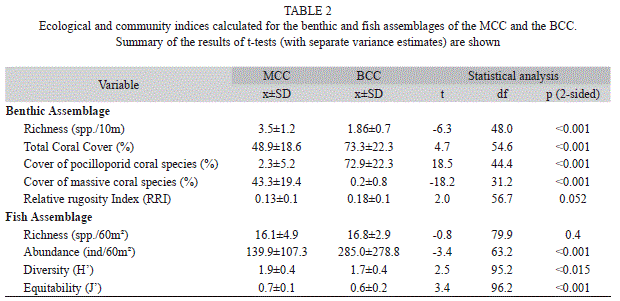
Benthic community: To assess the biological and physical characteristics of the benthic community at BCC and MCC, we applied the “chain-transect method” (Risk, 1972; Glynn et al., 1982; McCormick, 1994) to obtain measures of coral richness (number of species/10m), benthic cover (%) and substrate rugosity. Following the method of Garzón-Ferreira, Reyes-Nivia & Rodríguez-Ramírez (2002), we randomly installed 30 linear transects of 10m length on each community, where a diver extended a chain following the substrate contour and recorded the number of links in contact with each benthic component (e.g., coral, algae, sand). For each transect, the cover percentage of each benthic component was calculated by dividing the number of links of each component by the total number of chain links and multiplying by 100. The relative rugosity index (RRI) was calculated following Wellington (1982), who considers the horizontal linear distance between the two ends of a transect and the distance between the same two points following the contour of the bottom in the formula: RRI=1–(Linear distance/Contour distance). Values approaching 1 are indicative of very high rugosity while values close to 0 are typical of flat areas.
Fish community: To assess the fish communities present at both sites we carried out underwater visual surveys while SCUBA diving (Garzón-Ferreira et al., 2002; Edgar et al., 2004). At each site we randomly conducted 50 surveys along 30x2m belt transects. In each survey a diver recorded the identity and abundance of all the fishes observed while swimming just over the substrate and twice along the transect. During the first run, only mobile and conspicuous species were recorded, whereas in the second run (returning on the same transect) benthic, cryptic and small fishes were surveyed through a thorough examination of the coral colonies and sea bottom. The surveys were carried out at depths between 5 and 10.6m at the MCC and between 2.3 and 7m at the BCC. We used data from the fish surveys to create a listing of all the species recorded and calculated community variables such as fish species richness (number of species/60m2), frequency of observation (%) and density (number of individuals/60m2). Additionally, we computed ecological indices such as Shannon’s diversity index (H’) and Pielou’s evenness index (J’), and classified the fish into five trophic categories (planktivores, carnivores, omnivores, herbivores, or parasitic) following Robertson & Allen (2006). All fish surveys and benthic assessments were carried out in two time periods: (1) from November 23 to December 5 of 2010 and (2) from February 22 to February 30 of 2011.
We examined differences between the MCC and BCC by comparing different community and ecological measures: (1) coral richness (spp./10m), (2) coral cover percentage (arcsine-square root-transformed), (3) relative rugosity index (RRI), (4) fish richness (spp./60m2), (5) total fish abundance (ind/60m2), (6) fish diversity (H’) and (7) fish evenness (J’). Statistical analyses were made using two-sample t-tests with separate variance estimates as the data did not satisfy the assumption of homoscedasticity (Zar, 1996). Additionally, to compare the densities of individual fish species (ind/60m2) between both sites we performed two-sample randomization tests because the data did not fulfill the normality and homocedasticity assumptions required by traditional parametric tests. The homogeneity of proportions of fish species richness in different trophic categories between the two coral communities was examined with a chi2 test. Variation in terms of total abundance (ind/60m2; modified with a Box-Cox transformation) per trophic group was analyzed through a factorial Analysis of Variance (ANOVA). For both fish and benthic communities, variation in species abundance and composition was assessed using a Non-Metric Multidimensional Scaling (nMDS; Faith, Minchin & Belbin, 1987) and an Analyses of Similarity (ANOSIM; Clarke, 1993) based on the Jaccard and the Bray-Curtis similarity indices (Magurran, 2004). The Jaccard index is an occurrence measure that takes into account just the presence/absence of species in a fish assemblage, while the Bray-Curtis index additionally considers the relative abundance of each of the taxa. Within the ANOSIM tests, R-values close to +1 indicate high dissimilarity between the established groups, while values near 0 indicate absence of dissimilarity. Finally, Similarity Percentage Analyses (SIMPER) were used to examine which species contributed most to the dissimilarities between both sites in terms of the benthic components and the fish assemblage. All analyses were performed using PAST 2.17 (Hammer, Harper & Ryan, 2001), Statistica 8 (Statsoft Inc., 2007), and Rundom Pro 3.14 (Jadwiszczak, 2009) statistics software.
Results
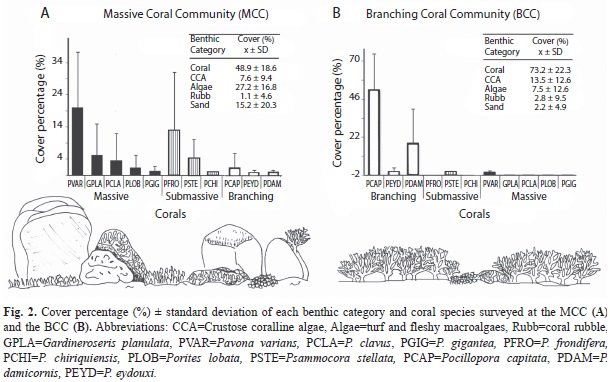
Benthic community comparisons: The MCC had a richer coral community and, as expected, a statistically higher cover of massive and submassive species than the BCC (Table 2; Fig. 2A). Among the eleven species of scleractinian corals recorded in the MCC, the massive coral P. varians had the highest substrate cover (20.1±17.2%; Fig. 2A). In contrast, the BCC had a higher total live coral cover and an expectedly greater abundance of branching coral colonies than the MCC (Table 2; Fig. 2B). In this case, the branching species P. damicornis and P. capitata dominated the coral assemblage with an average cover of 54.2±25.2% and 18.2±21.5%, respectively (Fig. 2B). However, despite the obvious differences between the two communities in dominant coral types and gross reefscape architecture (Fig. 2), the rugosity index obtained through the chain-transect method did not reflect significant differences in topographic complexity (Table 2).
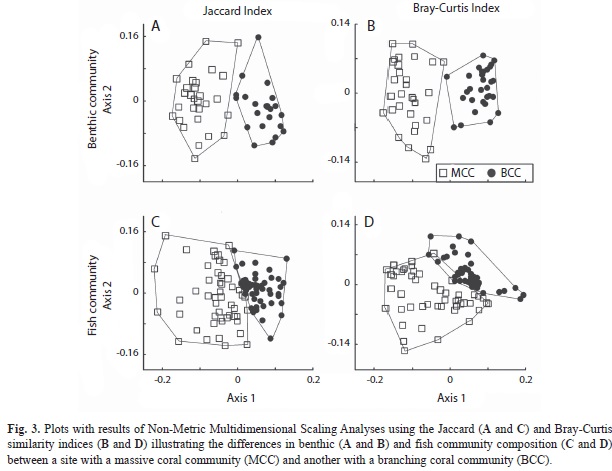
As suggested by the previous results, the BCC and MCC were very different in terms of both their benthic composition (Jaccard index, ANOSIM; R=0.79, p<0.001; Fig. 3A) and the relative abundance of those components (Bray-Curtis index, ANOSIM; R=0.87, p<0.001; Fig. 3B). The SIMPER analysis revealed that 39.3% of the dissimilarity between the two coral assemblages was due to variation in the abundance of the branching corals (P. capitata and P. damicornis) which covered 70% of the benthic cover at the BCC, but represented 3% of the MCC substrate. Other components having a major contribution to the dissimilarity between both communities were green algae, P. varians, P. frondifera, and sand, all of which had greater cover in the MCC (Fig. 2).
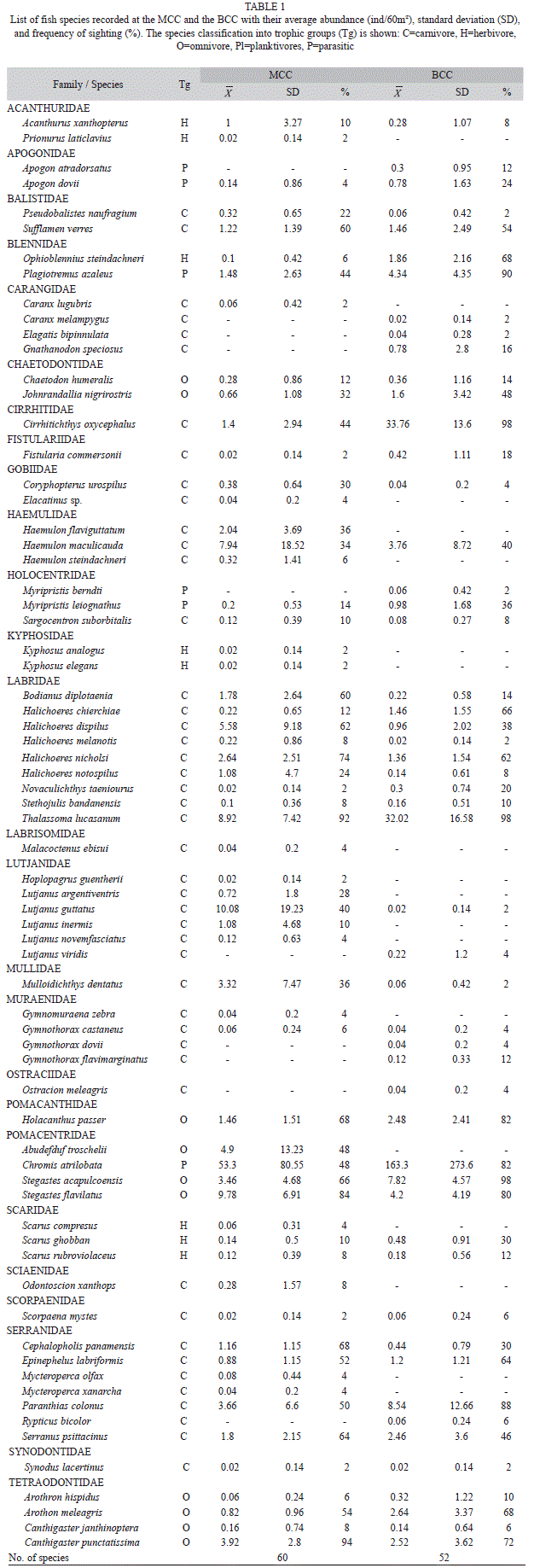
Fish community comparisons: A total of 21 249 individuals and 70 fish species from 26 families were surveyed at the MCC and BCC sites (Table 1). Both of these coral communities had similar total (67 vs 69 species, respectively) and mean fish species richness (~16 species/60m2; Table 2). However, the BCC sustained a more abundant, but less diverse and less even fish community than the MCC (Table 2). Low diversity and evenness of the BCC fish assemblage was due a the small number of species (Chromis atrilobata, Cirrhitichthys oxycephalus, Thalassoma. lucasanum, Stegastes acapulcoensis and Paranthias colonus) accounting for over 85% of all the individuals recorded in that community.
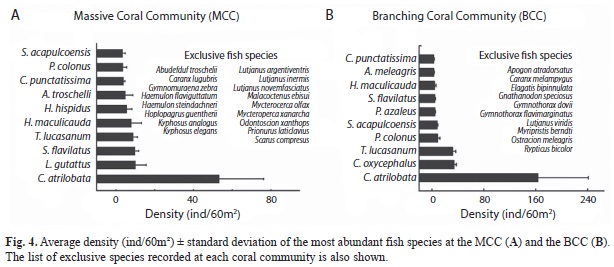
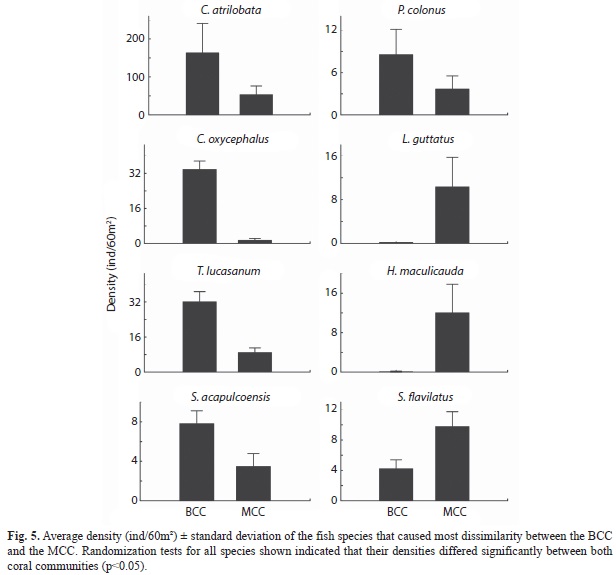
Although the MCC and BCC shared 42 fish species, the ANOSIM based on the Jaccard index indicated that the two fish assemblages differed significantly in species composition (R=0.39, p<0.001; Fig. 3C). The MCC sustained 17 exclusive species, 53% of them belonging to the families Lutjanidae, Haemulidae, Serranidae and Kyphosidae (Fig. 4A). Conversely, in the BCC the family Carangidae contributed three of the ten exclusive species (Fig. 4B). Considering the relative abundance of fish species, the ANOSIM test based on the Bray-Curtis index corroborated that the MCC and BCC were inhabited by distinct fish assemblages (ANOSIM; R=0.41, p<0.001; Fig. 3D). The SIMPER analysis revealed that 70% of the dissimilarity between the two fish assemblages inhabiting the two coral communities was due to differences in the abundance of C. atrilobata (28% contribution), C. oxycephalus (10%), T. lucasanum (7.5%), L. guttatus (2.7%) and S. flavilatus (2.6%; Fig. 4). The first three species in addition to P. colonus (represented mostly by juveniles and small adults) held significantly higher densities in the BCC than in the MCC (randomization tests: p?0.005; Fig. 5). In contrast, L. guttatus, H. maculicauda and S. flavilatus were more abundant in the MCC than in the BCC (randomization tests: p?0.05; Fig. 5). Interestingly, the congeneric damselfishes S. acapulcoensis and S. flavilatus exhibited opposite patterns (Fig. 5).
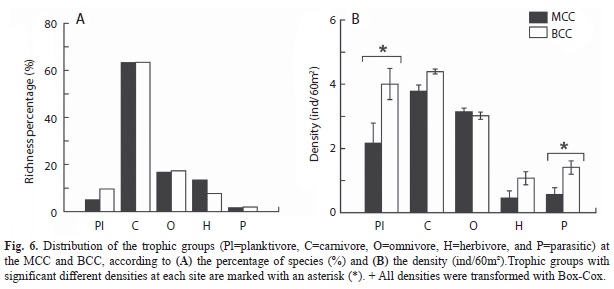
The species richness of the five trophic groups was nearly identical at both sites (?2=1.67, df=4, p=0.8; Fig. 6A), with carnivores being the group with the largest number of species. In contrast, we found a significant interaction effect of trophic group and coral community type on mean density per trophic group (Two-way ANOVA, F4,490=11.54, p?0.001). Which was explained by the higher abundances that planktivores and parasitic fish (represented by Plagiotremus azaleus) had in the BCC in comparison with the MCC (Fig. 6).
Discussion
By comparing the fish assemblages of two coral communities that differed mainly in their dominant coral morphologies (massive vs. branching), we examined how coral morphology and the resulting reefscape architecture affects the assemblage structure of conspicuous fish. Our results suggest that reefscape architecture, shaped by dominant coral morphologies, plays a major role structuring the fish assemblages that inhabit tropical coral communities. We particularly found that massive corals forge a complex reefscape architecture that sustains a more diverse and even fish community. However, detailed analysis of the fish species composition and relative abundance in each coral community revealed that preference towards massive or branching coral architectures depends on the fishes’ size and trophic preferences. Other authors have arrived at similar conclusions and have also noted the role that the mobility of fishes can play in determining species-habitat associations (Dominici-Arosemena & Wolf, 2006; Benfield et al., 2008; Alvarez-Filip et al., 2011a; Kerry & Bellwood, 2012).
Our finding that the MCC sustains a more diverse and even assemblage of conspicuous fish than the BCC, can be explained by the fact that this community was richer in coral species, and hence morphologies, than the BCC. Furthermore, dominant massive coral species, despite their simple and smooth morphology, exhibited a wide range of sizes (up to 2m height) and shapes (e.g., globular, plate) that increased the complexity of the reefscape architecture and offered a wide variety of microhabitats and resources for different types of fish species (Roberts & Ormond, 1987; Friedlander & Parrish, 1998; Gratwicke & Speight, 2005). The BCC, on the other hand, despite being dominated by branching species that are morphologically complex at a small scale (Beukers & Jones, 1997; Almany, 2004), at a larger scale (meters to tens of meters) have a very simple and plain topography as the colonies grow very tightly in a monotonous framework. Although this type of reefscape may enhance the abundance of small, cryptic fishes that inhabit small microhabitats within coral branches (see below), it cannot be directly used as refuge by large, conspicuous fishes and it therefore does not favor their richness, diversity or evenness (Dominici-Arosemena & Wolff, 2006; Benfield et al., 2008). Even though the dominant coral morphologies appear to shape the reefscape architecture, the reef complexity does not necessarily depend on the rugosity of the foundation taxa, but on the variety of morphological attributes that coral species have and the disposition or arrangement that they take within the reefscape (Alvarez-Filip et al., 2011a).
In our study, results of total fish density were not considered to be related to reef architecture, as pocilloporid frameworks in the TEP are known to have very high total fish abundances due to the particular oceanographic conditions in which they develop (shallow and exposed zones constantly subjected to relatively strong currents; Dominici-Arosemena & Wolff, 2006). Such conditions usually boost the densities of highly abundant species like C. atrilobata and T. lucasanum that in consequence bias the total abundance estimates of the fish community (Zapata & Morales, 1997; Dominici-Arosemena & Wolff, 2006; Alvarez-Filip et al., 2006). It is important to point out however, that the fact that the highest total fish abundance was recorded at the BCC (with the flattest reefscape architecture) emphasizes the influence that environmental conditions and dominant fish species have in shaping the fish assemblage (Bradbury & Young 1981; Dominici-Arosemena & Wolff, 2006).
As some community attributes and ecological indices mask the responses of particular fish species or groups, we considered the presence and abundance of each of the fishes at the MCC and BCC to detect associations of certain fish species with a particular architectural reefscape or dominant coral morphology. Fishes attracted to the BCC had a strong preference for the shelter and food that pocilloporid colonies offer. For instance, the branching colonies promote the presence of small territorial fish (e.g., C. oxycephalus, O. steindachneri) and juveniles of certain species (e.g., P. colonus) as the branches and interstices of ramose corals provide a complex fine-scale microhabitat that effectively shelters them from predators (Beukers & Jones, 1997; Almany, 2004; McCormick & Hoey, 2006; Alvarez-Filip et al., 2011a; Kerry & Bellwood, 2012). Although we did not consider fish sizes in our study, we hypothesize that the fish richness recorded at the BCC (comparable to the one at the MCC) might result from the presence of small juveniles from a wide variety of species that use this type of habitat as refuge during their early life stages. It is likely that the community of species associated with this type of microhabitat might be even richer than we observed as visual surveys are known to undestimate the richness and abundance of cryptobenthic fishes that seek shelter between coral branches. Additional to small fish, the BCC also tended to attract corallivore fishes such as the pufferfish Arothron meleagris, which feeds from the branch tips of Pocillopora spp. (Glynn et al., 1982) and is usually found in high densities on coral communities with high pocilloporid cover (Palacios, Muñoz & Zapata, in prep.).
In the MCC, the complex reefscape architecture, forged by the variety of shapes and sizes that massive coral species had, attracted mainly large fishes with roving behavior and piscivore-feeding preferences (e.g., species from the families Lutjanidae, Haemulidae and Serranidae). Large predator species are known to frequent massive or tabular coral forms in search of resting places, concealment or adequate grounds for their ambush predation (Dominici-Arosemena & Wolff, 2006; Kerry & Bellwood, 2012). In accordance with the decrease in the number of planktivores recorded at the MCC, Alvarez-Filip et al. (2011a) noted that at sites with a complex reefscape, the ratio of fish from high-trophic guilds (carnivores) to low trophic guilds (planktivores and herbivores) is reduced. The particularly high abundance of L. guttatus at the MCC (usually absent or at very low densities on TEP coral reefs; Zapata & Morales, 1997; Alvarez-Filip et al., 2006; Dominici-Arosemena & Wolff, 2006) along with the presence of large fishes usually associated with rocky habitats (e.g., Hoplopagrus guentherii, Mycteroperca spp., Lutjanus novemfasciatus; Robertson & Allen, 2006) may support the idea that structural reefscapes of massive corals and deep-rocky reefs can be analogous habitats with largely overlapping fish communities (Robertson, 1998; Bellwood, 1998; Dominici-Arosemena & Wolf, 2006).
When assessing the architectural complexity of the MCC and BCC, it was noteworthy that the relative rugosity index used in this study failed to demonstrate the evident structural differences between both coral communities. However, considering the critiques made by McCormick (1994) and Wedding et al. (2008) to the “chain transect method”, we ascribe our results to specific limitations that this method has when assessing large spatial scales and differentiating contrasting landscape patterns. McCormick (1994) pointed out that similar index values can be obtained from substrata with low/inclined corrugations (reefscape similar to the BCC) and substrata with tabular/globular outcrops (such as those at the MCC). Additionally, we must acknowledge that our results extend only to assemblages of large, mobile, conspicuous fish and that if assessed, other patterns and conclusion may be drawn from small, cryptobenthic fish assemblages.
Our study provides evidence that the dominant coral morphology of a coral community is an important habitat feature influencing the characteristics of the coral reef fish assemblage (Luckhurst & Luckhurst, 1978; Friedlander & Parrish, 1998; McCormick, 1994; Wilson et al., 2007; Alvarez-Filip et al., 2011a). For large, mobile and conspicuous fishes, massive coral species proved to be very important foundation taxa as they increase the gross reefscape complexity through the variety of morphological attributes they exhibit. Branching pocilloporid colonies, despite shaping a flat and monotonous framework, proved to attract small fish seeking shelter at a micro-scale level. It appears that the current loss of massive corals, the architectural flattening of the reefscape and community shifts towards monotonous stands of stress-resistant coral taxa (e.g., Pocillopora spp.), would pose a risk mainly to the large and carnivorous species in the fish assemblage (Pratchett et al., 2008; Alvarez-Filip et al., 2011b; Kerry & Bellwood, 2012). Massive coral species, which are in fact very vulnerable to natural and anthropogenic disturbances (Jones & Syms, 1998), require critical protection not only to guarantee the complexity of the reefscape architecture, but to support the large fishes from high trophic levels and thus maintain the diversity and evenness of the fish assemblage.
Acknowledgments
We thank Fondo para la Acción Ambiental y la Niñez, Conservación Internacional Colombia and Universidad del Valle for the funding that made this study possible. Additionally we are grateful to Parques Nacionales Naturales for providing all the logistical support at Gorgona Island and granting research permit PIBD-DTSO 011-10. Teammates Diego Lozano-Cortés, Carlos G. Muñoz, Ana Lucía Castrillón, and particularly Juan Sebastián Ayala are most gratefully acknowledged for their help with field work.
References
Almany, G. R. (2004). Does increased habitat complexity reduce predation and competition in coral reef fish assemblages? Oikos, 106: 275-285. [ Links ]
Alvarez-Filip, L., Reyes-Bonilla, H. & Calderon-Aguilera, L. E. (2006). Community structure of fishes in Cabo Pulmo reef, Gulf of California. Marine Ecology Progress Series, 27: 253-262. [ Links ]
Alvarez-Filip, L., Gill, J. A. & Dulvy, N. K. (2011a). Complex reef architecture supports more small-bodied fishes and longer food chains on Caribbean reefs. Ecosphere, 2: 10. [ Links ]
Alvarez-Filip, L., Dulvy, N. K., Côté, I. M, Watkinson, A. R. & Gill, J. A. (2011b). Coral identity underpins architectural complexity on Caribbean reefs. Journal of Applied Ecology, 21: 2223-2231. [ Links ]
Bellwood, D. R. (1998). What are reef fishes?-Comment on the report by DR Robertson: Do coral-reef fish faunas have a distinctive taxonomic structure? (Coral Reefs 17: 179-186). Coral Reefs, 17: 187-189. [ Links ]
Benfield, S., Baxter, L., Guzman, H. M. & Mair, J. M. (2008). A comparison of coral reef and coral communities fish assemblages in Pacific Panama and environmental factors governing their structure. Journal of the Marine Biological Association of the United Kingdom, 88: 1331-1341. [ Links ]
Beukers, J. S. & Jones, G. P. (1997). Habitat complexity modifies the impact of piscivores on a coral reef fish population. Oecologia, 114: 50-59. [ Links ]
Bradbury, R. H. & Young, P. C. (1981). The effects of a major forcing function, wave energy, on a coral reef ecosystem. Marine Ecology Progress Series, 5: 229-241. [ Links ]
Clarke, K. R. (1993). Non-parametric multivariate analysis of changes in community structure. Australian Journal of Ecology, 18: 117-143. [ Links ]
Dominici-Arosemena, A. & Wolff, M. (2006). Reef fish community structure in the tropical Eastern Pacific (Panamá): living on a relatively stable rocky reef environment. Helgoland Marine Research, 60: 287-305. [ Links ]
Edgar, G. J., Barrett, N. S. & Morton, A. J. (2004). Biases associated with the use of underwater visual census techniques to quantify the density and size-structure of fish populations. Journal of Experimental Marine Biology and Ecology, 308: 269-290. [ Links ]
Faith, D. P., Minchin, P. R. & Belbin, L. (1987). Compositional dissimilarity as a robust measure of ecological distance. Vegetation, 69: 57-68. [ Links ]
Friedlander, A. M. & Parrish, F. (1998). Habitat characteristics affecting fish assemblages on a Hawaiian coral reef. Journal of Experimental Marine Biology and Ecology, 224: 1-30. [ Links ]
Garzón-Ferreira, J., Reyes-Nivia, M. C. & Rodríguez-Ramírez, A. (2002). Manual de métodos del Sistema Nacional de Monitoreo de Arrecifes Coralinos en Colombia. INVEMAR, Santa Marta, Colombia. [ Links ]
Glynn, P. W. & Ault, J. S. (2000). A biogeographic analysis and review of the far eastern Pacific coral reef region. Coral Reefs, 19: 1-23. [ Links ]
Glynn, P. W., Prahl, H. v. & Guhl, F. (1982). Coral reefs of Gorgona Island, with special reference to corallivores and their influence on community structure and reef development. Anales del Instituto de Investigaciones Marinas de Punta de Betín, 12: 185-214. [ Links ]
Gratwicke, B. & Speight, M. R. (2005). The relationship between fish species richness, abundance and habitat complexity in a range of shallow tropical marine habitats. Journal of Fish Biology, 66: 650-667. [ Links ]
Hammer, O., Harper, D. A. T. & Ryan, P. D. (2001). PAST: Paleontological Statistics software package for education and data analysis, version 2.17. Retrieved from http://folk.uio.no/ohammer/past/ [ Links ]
Hutchinson, G. E. (1959). Homage to Santa Rosalia or why are there so many kinds of animals? American Naturalist, 93: 145-159. [ Links ]
Jadwiszczak, P. (2009). Rundom Pro, version 3.14. Retrieved from http://pjadw.tripod.com [ Links ]
Jones, G. P. & Syms, C. (1998). Disturbance, habitat structure and the ecology of fishes on coral reefs. Australian Journal of Ecology, 23: 287-297. [ Links ]
Kerry, J. T. & Bellwood, D. R. (2012). The effect of coral morphology on shelter selection by coral reef fishes. Coral Reefs, 31: 415-424. [ Links ]
Luckhurst, B. E. & Luckhurst, K. (1978). Analysis of the influence of substrate variables on coral reef fish communities. Marine Biology, 49: 317-323. [ Links ]
MacArthur, R. H. & MacArthur, J. W. (1961). On bird species diversity. Ecology, 42: 594-598. [ Links ]
Magurran, A. E. (2004). Ecological diversity and its measurement. Blackwell Science LTD, USA. [ Links ]
McCormick, M. I. (1994). Comparison of field methods for measuring surface topography and their associations with a tropical reef fish assemblage. Marine Ecology Progress Series, 112: 87-96. [ Links ]
McCormick, M. I. & Hoey, A. S. (2006). Biological and physical correlates of settlement and survival for a coral reef fish, Pomacentrus amboinensis (Pomacentridae). Coral Reef Symposium, 1: 425-430. [ Links ]
Messmer, V., Jones, G. P., Munday, P. L., Holbrook, S. J., Schmitt, R. J. & Brooks, A. J. (2011). Habitat biodiversity as a determinant of fish community structure on coral reefs. Ecology 92: 2285-2298. [ Links ]
Pratchett, M. S., Munday, P. L., Wilson, S. K., Graham N. A. J., Cinner, J. E., Bellwood, D. R., Jones, G. P., Polunin, N. V. C. & McClanahan, T. R. (2008). Effects of climate-induced coral bleaching on coral-reef fishes-ecological and economic consequences. Journal of Marine Biology & Oceanography, 46: 251-296. [ Links ]
Prahl, H. v. & Erhardt, H. (1985). Colombia: Corales y arrecifes coralinos. Fondo FEN. Editorial Presencia, Bogotá, Colombia. [ Links ]
Risk, M. J. (1972). Fish diversity on a coral reef in the Virgin Islands. Atoll Research Bulletin, 193: 1-6. [ Links ]
Roberts, C. M. & Ormond, R. F. (1987). Habitat complexity and coral reef fish diversity and abundance on Red Sea fringing reefs. Marine Ecology Progress Series, 41: 1-8. [ Links ]
Robertson, D. R. (1998). Do coral-reef fish faunas have a distinctive taxonomic structure? Coral Reefs, 17: 179-186. [ Links ]
Robertson, D. R. & Allen, G. R. (2006). Shorefishes of the tropical eastern Pacific: an information system. Version 2.0. Smithsonian Tropical Research Institute. Balboa, Panamá [ Links ].
StatSoft, Inc. 2007. STATISTICA (data analysis software system), version 8.0. Retrieved from www.statsoft.com [ Links ]
Tews, J., Brose U., Grimm, V., Tielborger, K., Wichmann, M. C., Schwager, M. & Jeltsch, F. (2004). Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. Journal of Biogeography, 31: 79-92. [ Links ]
Wedding, L. M., Friedlander, A. M., McGranaghan, M., Yost, R. S. & Monaco, M. E. (2008). Using bathymetric lidar to define nearshore benthic habitat complexity: Implications for management of reef fish assemblages in Hawaii. Remote Sensing of Environment, 112: 4159-4165. [ Links ]
Wellington, G. M. (1982). Depth zonation of corals in the Gulf of Panama: control and facilitation by resident reef fishes. Ecological Monographs, 52: 223-241. [ Links ]
Wilson, S. K., Graham, N. A. J. & Polunin, N. V. C. (2007). Appraisal of visual assessments of habitat complexity and benthic composition on coral reefs. Marine Biology, 151: 1069-1076. [ Links ]
Zapata, F. A. & Morales, Y. A. (1997). Spatial and temporal patterns of fish diversity in a coral reef at Gorgona island, Colombia. Proc. 8th Int. Coral Reef Symposium, 1: 1029-1034. [ Links ]
Zapata, F. A. & Vargas-Ángel, B. (2003). Corals and coral reefs of the Pacific coast of Colombia. In J. Cortés (Ed.), Coral reefs of Latin America (pp. 419- 447). Elsevier Science, Amsterdam. [ Links ]
Zar, J. H. (1996). Biostatistical Analysis. Prentice Hall. New Jersey. USA. [ Links ]
Alvarez-Filip, L., Reyes-Bonilla, H. & Calderon-Aguilera, L. E. (2006). Community structure of fishes in Cabo Pulmo reef, Gulf of California. Marine Ecology Progress Series, 27: 253-262. [ Links ]
Alvarez-Filip, L., Gill, J. A. & Dulvy, N. K. (2011a). Complex reef architecture supports more small-bodied fishes and longer food chains on Caribbean reefs. Ecosphere, 2: 10. [ Links ]
Alvarez-Filip, L., Dulvy, N. K., Côté, I. M, Watkinson, A. R. & Gill, J. A. (2011b). Coral identity underpins architectural complexity on Caribbean reefs. Journal of Applied Ecology, 21: 2223-2231. [ Links ]
Bellwood, D. R. (1998). What are reef fishes?-Comment on the report by DR Robertson: Do coral-reef fish faunas have a distinctive taxonomic structure? (Coral Reefs 17: 179-186). Coral Reefs, 17: 187-189. [ Links ]
Benfield, S., Baxter, L., Guzman, H. M. & Mair, J. M. (2008). A comparison of coral reef and coral communities fish assemblages in Pacific Panama and environmental factors governing their structure. Journal of the Marine Biological Association of the United Kingdom, 88: 1331-1341. [ Links ]
Beukers, J. S. & Jones, G. P. (1997). Habitat complexity modifies the impact of piscivores on a coral reef fish population. Oecologia, 114: 50-59. [ Links ]
Bradbury, R. H. & Young, P. C. (1981). The effects of a major forcing function, wave energy, on a coral reef ecosystem. Marine Ecology Progress Series, 5: 229-241. [ Links ]
Clarke, K. R. (1993). Non-parametric multivariate analysis of changes in community structure. Australian Journal of Ecology, 18: 117-143. [ Links ]
Dominici-Arosemena, A. & Wolff, M. (2006). Reef fish community structure in the tropical Eastern Pacific (Panamá): living on a relatively stable rocky reef environment. Helgoland Marine Research, 60: 287-305. [ Links ]
Edgar, G. J., Barrett, N. S. & Morton, A. J. (2004). Biases associated with the use of underwater visual census techniques to quantify the density and size-structure of fish populations. Journal of Experimental Marine Biology and Ecology, 308: 269-290. [ Links ]
Faith, D. P., Minchin, P. R. & Belbin, L. (1987). Compositional dissimilarity as a robust measure of ecological distance. Vegetation, 69: 57-68. [ Links ]
Friedlander, A. M. & Parrish, F. (1998). Habitat characteristics affecting fish assemblages on a Hawaiian coral reef. Journal of Experimental Marine Biology and Ecology, 224: 1-30. [ Links ]
Garzón-Ferreira, J., Reyes-Nivia, M. C. & Rodríguez-Ramírez, A. (2002). Manual de métodos del Sistema Nacional de Monitoreo de Arrecifes Coralinos en Colombia. INVEMAR, Santa Marta, Colombia. [ Links ]
Glynn, P. W. & Ault, J. S. (2000). A biogeographic analysis and review of the far eastern Pacific coral reef region. Coral Reefs, 19: 1-23. [ Links ]
Glynn, P. W., Prahl, H. v. & Guhl, F. (1982). Coral reefs of Gorgona Island, with special reference to corallivores and their influence on community structure and reef development. Anales del Instituto de Investigaciones Marinas de Punta de Betín, 12: 185-214. [ Links ]
Gratwicke, B. & Speight, M. R. (2005). The relationship between fish species richness, abundance and habitat complexity in a range of shallow tropical marine habitats. Journal of Fish Biology, 66: 650-667. [ Links ]
Hammer, O., Harper, D. A. T. & Ryan, P. D. (2001). PAST: Paleontological Statistics software package for education and data analysis, version 2.17. Retrieved from http://folk.uio.no/ohammer/past/ [ Links ]
Hutchinson, G. E. (1959). Homage to Santa Rosalia or why are there so many kinds of animals? American Naturalist, 93: 145-159. [ Links ]
Jadwiszczak, P. (2009). Rundom Pro, version 3.14. Retrieved from http://pjadw.tripod.com [ Links ]
Jones, G. P. & Syms, C. (1998). Disturbance, habitat structure and the ecology of fishes on coral reefs. Australian Journal of Ecology, 23: 287-297. [ Links ]
Kerry, J. T. & Bellwood, D. R. (2012). The effect of coral morphology on shelter selection by coral reef fishes. Coral Reefs, 31: 415-424. [ Links ]
Luckhurst, B. E. & Luckhurst, K. (1978). Analysis of the influence of substrate variables on coral reef fish communities. Marine Biology, 49: 317-323. [ Links ]
MacArthur, R. H. & MacArthur, J. W. (1961). On bird species diversity. Ecology, 42: 594-598. [ Links ]
Magurran, A. E. (2004). Ecological diversity and its measurement. Blackwell Science LTD, USA. [ Links ]
McCormick, M. I. (1994). Comparison of field methods for measuring surface topography and their associations with a tropical reef fish assemblage. Marine Ecology Progress Series, 112: 87-96. [ Links ]
McCormick, M. I. & Hoey, A. S. (2006). Biological and physical correlates of settlement and survival for a coral reef fish, Pomacentrus amboinensis (Pomacentridae). Coral Reef Symposium, 1: 425-430. [ Links ]
Messmer, V., Jones, G. P., Munday, P. L., Holbrook, S. J., Schmitt, R. J. & Brooks, A. J. (2011). Habitat biodiversity as a determinant of fish community structure on coral reefs. Ecology 92: 2285-2298. [ Links ]
Pratchett, M. S., Munday, P. L., Wilson, S. K., Graham N. A. J., Cinner, J. E., Bellwood, D. R., Jones, G. P., Polunin, N. V. C. & McClanahan, T. R. (2008). Effects of climate-induced coral bleaching on coral-reef fishes-ecological and economic consequences. Journal of Marine Biology & Oceanography, 46: 251-296. [ Links ]
Prahl, H. v. & Erhardt, H. (1985). Colombia: Corales y arrecifes coralinos. Fondo FEN. Editorial Presencia, Bogotá, Colombia. [ Links ]
Risk, M. J. (1972). Fish diversity on a coral reef in the Virgin Islands. Atoll Research Bulletin, 193: 1-6. [ Links ]
Roberts, C. M. & Ormond, R. F. (1987). Habitat complexity and coral reef fish diversity and abundance on Red Sea fringing reefs. Marine Ecology Progress Series, 41: 1-8. [ Links ]
Robertson, D. R. (1998). Do coral-reef fish faunas have a distinctive taxonomic structure? Coral Reefs, 17: 179-186. [ Links ]
Robertson, D. R. & Allen, G. R. (2006). Shorefishes of the tropical eastern Pacific: an information system. Version 2.0. Smithsonian Tropical Research Institute. Balboa, Panamá [ Links ].
StatSoft, Inc. 2007. STATISTICA (data analysis software system), version 8.0. Retrieved from www.statsoft.com [ Links ]
Tews, J., Brose U., Grimm, V., Tielborger, K., Wichmann, M. C., Schwager, M. & Jeltsch, F. (2004). Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. Journal of Biogeography, 31: 79-92. [ Links ]
Wedding, L. M., Friedlander, A. M., McGranaghan, M., Yost, R. S. & Monaco, M. E. (2008). Using bathymetric lidar to define nearshore benthic habitat complexity: Implications for management of reef fish assemblages in Hawaii. Remote Sensing of Environment, 112: 4159-4165. [ Links ]
Wellington, G. M. (1982). Depth zonation of corals in the Gulf of Panama: control and facilitation by resident reef fishes. Ecological Monographs, 52: 223-241. [ Links ]
Wilson, S. K., Graham, N. A. J. & Polunin, N. V. C. (2007). Appraisal of visual assessments of habitat complexity and benthic composition on coral reefs. Marine Biology, 151: 1069-1076. [ Links ]
Zapata, F. A. & Morales, Y. A. (1997). Spatial and temporal patterns of fish diversity in a coral reef at Gorgona island, Colombia. Proc. 8th Int. Coral Reef Symposium, 1: 1029-1034. [ Links ]
Zapata, F. A. & Vargas-Ángel, B. (2003). Corals and coral reefs of the Pacific coast of Colombia. In J. Cortés (Ed.), Coral reefs of Latin America (pp. 419- 447). Elsevier Science, Amsterdam. [ Links ]
Zar, J. H. (1996). Biostatistical Analysis. Prentice Hall. New Jersey. USA. [ Links ]
1. Coral Reef Ecology Research Group, Department of Biology, Universidad del Valle, A.A. 25360, Cali, Colombia.School of Marine & Tropical Biology, James Cook University, Townsville, Queensland 4811, Australia;mariadelmar.palacios@gmail.com
2. Coral Reef Ecology Research Group, Department of Biology, Universidad del Valle, A.A. 25360, Cali, Colombia; fernando.zapata@correounivalle.edu.co
Recibido 18-X-2013. Corregido 20-XI-2013. Aceptado 19-XII-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}