










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 suppl.1 San José Feb. 2014
Estructura y diversidad de la vegetación del Parque Nacional Natural de la Isla Gorgona, Colombia
Structure and diversity of the vegetation of the Gorgona Island National Natural Park, Colombia
Structure and diversity of the vegetation of the Gorgona Island National Natural Park, Colombia
Abstract
Gorgona is a continental island in the Pacific of Colombia. We studied the structure and diversity of its vegetation with 16 Gentry transects (total area 0.16ha) in nine sites on the island. With these data, we calculate Importance Value Index (IVI), distribution of individuals according to their height and diameter at breast height (DBH), cumulative species curves, and species diversity. The vegetation is mainly secondary forest, with remnants of primary forest on the hills. By density, frequency, and coverage, the most important family is Rubiaceae, with 0.275 IVF. The species with the highest IVI (0.157) is Lacistema aggregatum (Lacistemataceae). In height and diameter classes (DBH), individuals have an inverted “J” distribution. The plant community has a Shannon-Wiener diversity index of 3.98. The cumulative species curve of the plant community does not level off even after more than ten transects. Transects were grouped in the similarity analysis but not strongly. There were species richness differences among transects. Rev. Biol. Trop. 62 (Suppl. 1): 13-26. Epub 2014 February 01.
Key words: Horizontal structure, vertical structure, plant diversity, Gentry transects, Gorgona Island, Chocó biogeographic region.
Resumen
En este trabajo, se estudió la estructura y diversidad de la vegetación en isla Gorgona, a través de 16 transectos de Gentry (área total de 0.16ha), distribuidos en nueve sitios de la isla. Se calculó el índice de Valor de Importancia para la especie y la familia (IVI e IVF respectivamente), la distribución de los individuos en función de su altura y diámetro a la altura del pecho (DAP ), las curvas de especies acumuladas , y la diversidad de especies. La vegetación de isla Gorgona estuvo conformada principalmente por bosque secundario, con restos de bosque primario en las partes altas de los cerros. De acuerdo con la densidad, la frecuencia, y la cobertura, la familia más importante fue Rubiaceae (IVF=0.275), mientras que la especie con el IVI más alto fue Lacistema aggregatum (Lacistemataceae, IVI=0.157). De acuerdo con la altura y clases de diámetro (DAP), la distribución de los individuos describió una forma de “J” invertida, con un valor de diversidad (H’) calculado de 3.98. Incluso después de más de diez transectos de muestreo, la curva de acumulación de especies no se niveló. Con base en el análisis de similitud, se estableció que los transectos estuvieron agrupados en zonas, de las cuales cuatro tuvieron un soporte estadístico, siendo incluso la riqueza de especies diferente entre los transectos.
Palabras clave: Estructura horizontal, estructura vertical, diversidad vegetal, transectos de Gentry, Isla Gorgona, Diámetro a la altura del pecho (DAP), Índice de valor de importancia (IVI).
El estudio de estructura y diversidad de la flora de un lugar en particular, proporciona información sobre la composición de la vegetación, permitiendo comprender los procesos bióticos y abióticos que podrían estar modulando esa formación vegetal particular. En general para conocer la vegetación de un lugar determinado, es necesario realizar estudios florísticos que suministren información rápida y fácil de manejar, de tal manera que se puedan llegar a comparar con la vegetación de diferentes localidades (Villareal et al., 2006; Rangel, 2004a; Vásquez-Vélez, Garzón & Ascencio-Santofinio, 2013).
Los bosques del neotrópico se pueden estudiar desde el punto de vista de su organización y su diversidad biológica. En el primer caso, se evalúan como están construidos, tras la mezcla aparentemente desordenada de los árboles y las especies, es decir definir su estructura. Generalmente, la estructura de los bosques describe la geometría de las poblaciones presentes, que parecen seguir ciertas leyes matemáticas (UNESCO, 1980), y se puede considerar como estructura vertical cuando corresponde a los patrones de distribución de los individuos entre la superficie del suelo y el dosel del bosque, definiendo pisos o estratos (Melo, 1994). Por otra parte, se puede considerar la estructura horizontal, como la variación de la distribución de las especies en el plano del bosque, como resultado de los gradientes ambientales y edáficos (Brunic, 1983). Esta última cuantificación se basa en parámetros tales como frecuencia, abundancia y dominancia o cobertura, cuya suma relativa genera el índice de importancia (IVI), índice que expresa el peso o importancia ecológica de la especie dentro del ecosistema (Villareal et al., 2006).
El otro aspecto que se utiliza para estudiar los ecosistemas boscosos es la diversidad biológica (Vélez & Fresneda, 1992; Magurran, 1988). La diversidad biológica se refiere a la variedad y abundancia de especies, a su composición genética, a las comunidades, ecosistemas y paisajes en los que ocurre; igualmente se refiere a las funciones y procesos en todos estos niveles. Este elemento puede manifestarse en diferentes escalas espaciales que van desde lo puntual, como los microhábitats, pasando por lo regional (diversidad-beta) hasta lo global (diversidad- gama) (Magurran, 1988). En ambientes insulares, la flora depende primordialmente de la cercanía o lejanía al continente y del tamaño de la misma, estas se dividen en islas oceánicas y en islas continentales. Las islas oceánicas son aquellas que se encuentran distantes de la plataforma continental, la única isla oceánica del Pacifico colombiano es la isla de Mapelo está conformada por musgos, un helecho rastrero y algunos parches de pastos conforman las angiospermas de la isla. Las islas continentales son aquellas que están próximas al continente y están vinculadas geológicamente por la plataforma submarina, este tipo de islas son más comunes en el Pacifico colombiano y entre estas se encuentra la isla Gorgona (Giraldo, 2012). La vegetación del sistema insular de Gorgona hace parte de la provincia del Chocó biogeográfico, caracterizada por ser una de las regiones más lluviosas del mundo, con una extensa red hidrológica y alberga una alta diversidad de especies a nivel de todos los grupos. Las plantas de la isla están directamente correlacionadas con la altitud, la distribución de franjas de vegetación y la presión por efecto de la intervención antrópica en sectores particulares de la isla, que permiten reconocer tres grupos principales: vegetación a lo largo de la playa, vegetación sucesional en estados de desarrollo y niveles de intervención y comunidades climáticas, correspondientes a la flora en su condición nativa, en mejor estado de conservación (Rangel, 2004a).
Hasta la fecha han sido publicados tres inventarios taxonómicos sobre la flora de isla Gorgona. El primero fue hecho por Cabrera (1983), seguido por el inventario de Murillo & Lozano (1989), y el último fue el publicado por Rangel (1990a). Adicionalmente, Vázquez-Vélez et al., (2012), realizaron recientemente un estudio general sobre la flora de isla Gorgona, registraron 723 especies entre Briofitas, Pteridofitas, Gimnospermas y Angiospermas, sin embargo estos resultados no han sido aun publicados. Considerando que la flora de isla Gorgona está catalogada como un valor objeto de conservación, toda vez que la porción insular de esta localidad fue declarada en 1984 como área natural protegida bajo la figura de Parque Nacional Natural (UAESPNN, 1998), en el presente estudio, se describe la estructura y diversidad vegetal en isla Gorgona, después de cerca de 30 años de un proceso de sucesión natural, teniendo en cuenta el último trabajo sistemático publicado sobre la flora de esta localidad por Rangel (1990a; 1990b; 1990c), 20 años atrás.
Materiales y métodos
Áreas de estudio: Isla Gorgona es una isla de origen volcánico ubicada a 35km al occidente de la costa pacífica colombiana. Tiene 9km de largo por 2.5km de ancho, y una extensión de 26km2. Administrativamente pertenece al municipio de Guapi, departamento del Cauca. El relieve es montañoso, siendo el punto de mayor altura el Cerro La Trinidad, con 338m sobre el nivel del mar. Este cerro, junto con los cerros Los Micos, La Esperanza y el Mirador, conforman la columna vertebral orográfica de la isla (Giraldo, 2012).
La vegetación del sistema insular de Gorgona hace parte de la provincia del Chocó biogeográfico, caracterizada por ser una de las regiones más lluviosas del mundo, con una extensa red hidrológica y alberga una alta diversidad de especies a nivel de todos los grupos. Las plantas de la isla están directamente correlacionadas con la altitud, la distribución de franjas de vegetación y la presión por efecto de la intervención antrópica en sectores particulares de la isla, que permiten reconocer tres grupos principales: vegetación a lo largo de la playa, vegetación sucesional en estados de desarrollo y niveles de intervención y comunidades climáticas, correspondientes a la flora en su condición nativa, en mejor estado de conservación (Rangel, 2004a).
En términos generales la vegetación natural en isla Gorgona está compuesta por tres asociaciones vegetales y dos comunidades, las cuales están caracterizadas por sus rasgos florísticos, ecológicos, fisionómicos y corológicos. A pesar de no ser muy alta, la isla presenta una diferenciación altitudinal, en las cuales dos asociaciones se presentan en la parte baja y media de la isla, entre 40 y 200m y la otra en la parte alta, de 220 a 300m. Éstas asociaciones están descritas como: Ossaeo sessilifoliae-Anaxagoretum phaedocarpae; Malphighio glabrae-Cespedesietum spathulatae y Cassipoureo ellipticae-Ryanetum speciosae (Rangel, 1990b; 1990c). Estructura y diversidad vegetal: Se utilizó el método de transecto de cinturón o método Gentry (1982) con algunas modificaciones. Se censó en un área de 0.1ha, todos los individuos cuyo tallo tenga un diámetro a la altura del pecho (DAP medido a 1.3m desde la superficie del suelo) mayor o igual a 1 cm, con el propósito de tener una mejor representación de los estratos inferiores del bosque (sotobosque) (Villareal et al., 2006). Se realizaron 16 transectos de 50m de largo por 2m de ancho, que se distribuyeron ordenadamente, manteniendo una distancia de separación mínima de 20m entre transectos. Los transectos no se interceptaron y fueron dispuestos de manera que se concentraran en un solo tipo de hábitat o unidad de paisaje.
Se realizaron dos esfuerzos de muestreo. El primer esfuerzo fue entre el 4 al 8 de marzo de 2011, y el segundo esfuerzo entre 24 al 29 de junio de 2011. Durante el primer esfuerzo de muestreo se recorrieron siete transectos que fueron dispuestos en el sendero hacia La Chonta (Transecto 1), el sendero hacia El Acueducto (Transecto 2), en el sector medio del sendero hacia playa Palmeras (Transecto 3), en el sector inicial del sendero al cerro El Mirador (Transecto 4), en el sector inicial del sendero al cerro de La Trinidad (Transecto 5), en la parte media del sendero al Cerro Los Micos (Transecto 6), y en la parte media del sendero que conduce hacia Yundigüa (Transecto 7). Durante el segundo esfuerzo de muestreo se recorrieron nueve transectos que fueron dispuestos en la parte final del sendero hacia Yundigüa (Transecto 8), en la parte final (Transecto 9) y baja (Transecto 10) del sendero al cerro Los Micos, en el sector de Playa Yundigüa (Transecto 11), en la parte final del sendero de playa Palmeras (Transecto 12), en la zona de El Mirador (Transecto 13), en la inmediaciones a la Cabaña de Playa Palmeras(Transecto 14), en la Playa de La Mancora (Transecto 15), y en la Playa de La Camaronera (Transecto 16).
Cada transecto se trazó con una cuerda, y con una varita de 1 m se estableció la distancia efectiva de censo a cada lado. Se censaron todos los individuos con DAP mayor o igual a 1cm, incluyendo árboles, arbustos, lianas, algunas hemiepífitas, y hierbas grandes. Se estimó su altura, se registró su hábito de crecimiento y todas las características que permitan posteriormente su reconocimiento, como son los colores de la corteza, madera, y exudado si está presente (Gentry, 1982; Villareal et al., 2006). Se realizó la determinación de las familias en el sitio utilizando la guía de campo de Gentry (1993) y el listado de flora realizada por Murillo & Lozano (1989). Para confirmar la identificación el material recolectado fue fotografía y se comparó con el material recolectado en el 2009 durante la expedición científica de Gorgona que se encuentra incluido en el herbario CUVC de la Universidad del Valle. Todo el material procesado se encuentra depositado en el herbario CUVC de la Universidad del Valle.
Análisis de datos: Con base en los registros e identificaciones se construyó un listado taxonómico con la mayor resolución posible (familia, género o especie). Se estimó la riqueza de especies como el número de especies presentes en un área muestreada de 0.8ha que corresponde a la sumatoria del área de los 16 trayectos que se trabajaron durante esta investigación. Adicionalmente se estimó la densidad total como el número total de individuos en 0.8ha y el área basal total como la sumatoria de las áreas basales de todos los individuos en 0.8ha.
Se calcularon los parámetros estructurales de Frecuencia de la especie i, Frecuencia relativa de la especie i, Densidad de una especie i, Densidad relativa de especie i, Cobertura de especie i, y Cobertura relativa de especie i. Con esta información se estimó el índice de valor de importancia para cada una de las especies en el área de estudio (IVI) y el índice de valor de importancia para cada una de las familias en el área de estudio (IVF). Los índices de valor de importancia proporcionan información de cuan dominante es cada especie o cada familia con respecto a la totalidad de las especies o familias registradas en un muestreo. Adicionalmente, con base en los registros de altura y diámetro de los tallos se evaluó el patrón de distribución de frecuencias por clases de alturas y por clases diamétricas de los tallos.
Para evaluar la representatividad de la riqueza de especies establecida con el esfuerzo de muestreo se utilizó el estimador no paramétrico de Chao-1 estimado con base en el esfuerzo de muestreo realizado, utilizando el programa EstimateS versión 7.5.0 (Estimate 2005). Este índice utiliza los registros de las especies únicas y especies dobles para calcular, mediante un proceso de re-muestreo estandarizado el valor esperado de la riqueza de especies en el área de estudio a partir de la información recolectada. Adicionalmente, se estimó la diversidad alfa por medio del índice de Shannon-Wiener, a partir de los registros de abundancia relativa generados para cada especie considerando cada transecto como una unidad de muestreo, siguiendo a Feinsinger (2003). Para evaluar si hay diferencias de composición vegetal entre los transectos se realizó un análisis de similaridad con presencia ausencia de especies con el programa PAST 2.15 (Hammer, Harper & Ryan, 2001) y se realizó una comparación de la riqueza de especies entre transectos mediante un análisis de rarefracción utilizando el programa iNEXT (considerando como punto de referencia el 52% de complejidad o cobertura de muestreo) siguiendo a Chao & Jost (2012) y Hsieh, Ma & Chao (2013), toda vez que los estimadores no paramétricos de riqueza, como Chao-1, no deben ser comparados directamente ya que son dependientes del esfuerzo de muestreo y de la presencia de especies únicas o raras.
Resultados
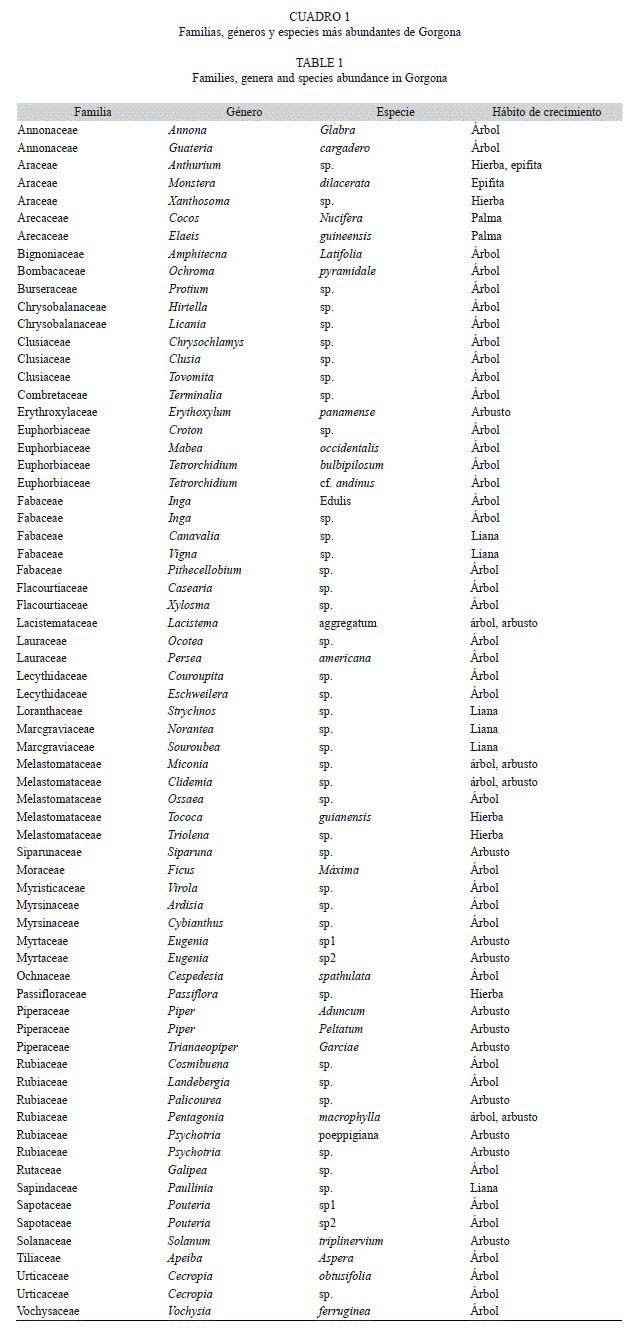
Estructura horizontal: En los 16 transectos efectuados se contabilizaron 1 138 individuos, clasificados en 231 especies, de 39 familias y 65 géneros (Cuadro 1). En términos generales la vegetación presente en el PNN Gorgona corresponde en su mayoría a un bosque secundario con restos de bosque primario en las lomas, con árboles pequeños a medianos (Fig. 1), seguido de árboles juveniles y arbustos. También fue común observar lianas de la familia Marcgraviaceae de gran grosor que colgaban y envolvían a los árboles como soporte para subir hasta el dosel. También se registraron hemiepifitas de la familia Clusiaceae, las cuales empiezan su etapa de crecimiento sobre otro árbol y luego lanzan sus raíces al suelo para continuar con su crecimiento.
Aunque se registraron algunas palmas naturales e introducidas en isla Gorgona, su abundancia fue relativamente baja en comparación con la zona continental (10 individuos en isla Gorgona contra 200 individuos en la localidad continental de Piangüita, ubicada en el sector externo de Bahía Buenaventura), probablemente por la ausencia de dispersores grandes y por la separación al continente. Sin embargo, estas especies fueron muy visitadas por los primates (Cebus capucinus curtus) presentes en la isla. En los claros de bosque fueron abundantes los especímenes de la familia Rubiaceae, con dominancia de Pentagonia macrophylla. También se identificaron especies típicas de sucesión secundaria como Cespedesia spathulata, Cecropia obtusifolia, Trema micrantha y Miconia sp. Las hierbas que predominaron fueron de los generos Selaginella, Triolena,Dieffenbachia y Mabea. Esta última estuvo en fruto durante junio y se escuchaba su dehiscencia cuando lanzaba las semillas.
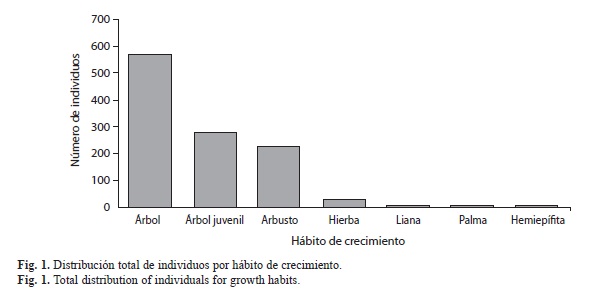
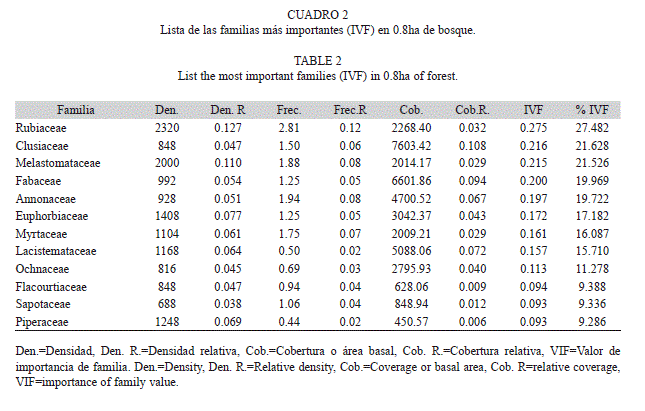
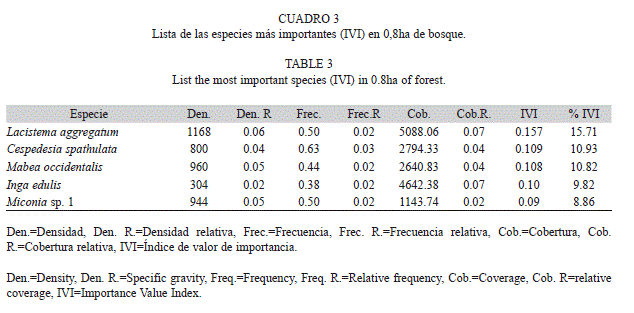
De acuerdo a la densidad, frecuencia y cobertura, las cinco familias más importante en isla Gorgona fueron Rubiaceae (IVF=0.28), Clusiaceae (IVF=0.22), Melastomataceae (IVF=0.22), Fabaceae (IVF=0.20) y Annonaceae (IVF=0.20) (Cuadro 2). Lacistema aggregatum (Familia Lacistemataceae) fue la especie con mayor valor de importancia (IVI=0.16), siendo registrada en el 50% de los transectos muestreados (Cuadro 3). En zonas de alta perturbación, C. spathulata (Familia Ochnaceae) y Miconia sp1 (familia Melastomataceae) fueron las especies con mayor valor de importancia, con IVI de 0.11 y 0.09 respectivamente (Cuadro 3). En los senderos hacia Yundigua y los cerros de Los Micos y La Trinidad, M. occidentalis (Familia Euphorbiaceae) fue la especie con mayor valor de importancia (IVI=0.11) (Cuadro 3).
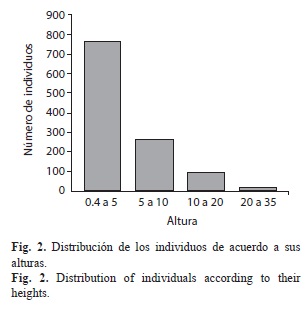
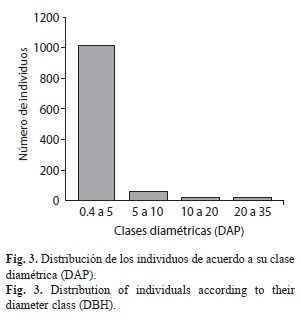
Estructura vertical: Isla Gorgona se caracterizó por presentar abundante vegetación de sotobosque distribuida principalmente en la parte baja de la isla, y árboles de crecimiento rápido pero de talla baja, condiciones que son características de vegetación de sucesión temprana. Por otro lado, hacia las cimas de los cerros Los Micos y La Trinidad se encuentran árboles de gran porte de la familia Clusiaceae, Myristicaceae y Fabaceae, aunque en las laderas hay alta perturbación por la caída natural de estos árboles. De acuerdo a las alturas y a las clases diamétricas (DAP) (Fig. 2, Fig. 3), se estableció una distribución de frecuencias de tipo “J” invertida, condición común en bosques perturbados, predominando individuos con altura entre 0.4 a 5m, y una clase diamétrica (DAP) entre 1 a 10cm.
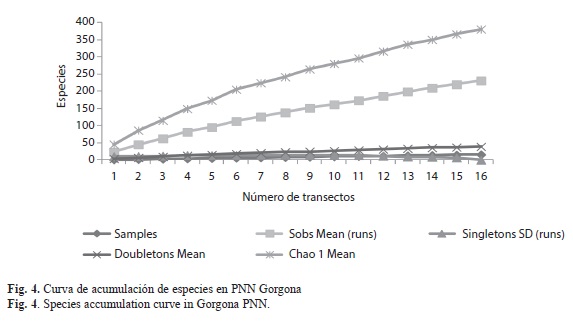
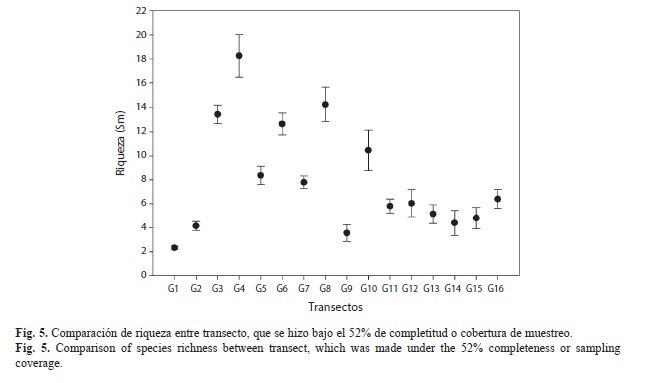
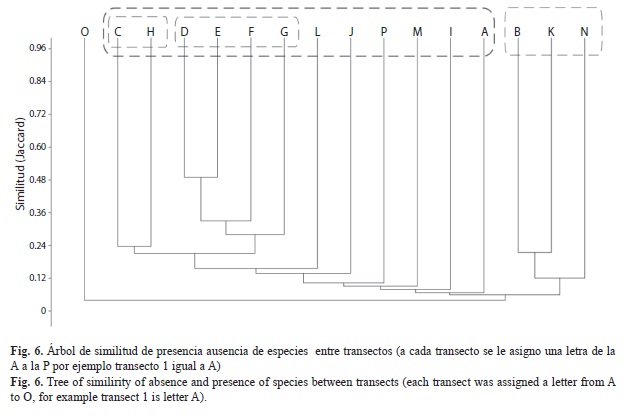
Diversidad y riqueza de especies: El número de especies registradas en Gorgona durante este trabajo fue de 231 especies, siendo el estimado de diversidad (H’) de 3.98. De acuerdo con el análisis no paramétrico de riqueza (Fig. 4), aunque se realizaron más de 10 transectos en el área de estudio, no se alcanzó el 100% de representatividad, lo que sugiere que aun hay especies de flora por registrar en esta localidad. Se establecieron diferencias significativas para la riqueza entre los transectos (Fig. 5), siendo los de mayor riqueza de especies los transectos 3 (G3), 4 (G4), 5 (G5), 6 (G6), 7 (G7) , 8 (G8), y 10 (G10), con el mismo número de especies, aunque algunas fueron exclusivas en un transecto en particular.
Discusión
La flora del PNN Gorgona es muy similar a la del continente, resultado que refuerza la hipótesis de que isla Gorgona estuvo unido al continente en el pasado geológico, con lazos biogeogáficos especialmente relacionados con la biota de la región pacífica (Vásquez-Vélez, Cortez, Calero, Soto & Torres, 2012). De acuerdo con Rangel (1990a), el proceso de separación fue reciente debido a la similitud de la flora entre la región Chocoana y también por el bajo porcentaje de endemismo presente en la isla. En este mismo sentido, también se podría suponer que isla Gorgona mantuvo condiciones ambientales óptimas durante los cambios climáticos fuertes del pleistoceno, por lo que podría ser considerada un refugio plestocénico (ver Gentry, 1982). Sin embargo, la alta riqueza de especies de algunos grupos como Melastomataceae (entre 4200-4500 especies; Clausing & Renner, 2001) y Rubiaceae (cerca de 5000 especies; Davis et al., 2009), depende principalmente por la alta diversidad de polinizadores y dispersores presentes en el neotrópico. Ambas familias tienen un alto valor ecológico, principalmente los géneros Miconia sp. Psychotria sp. y Palicourea sp., géneros que son son fuente importante de alimento para animales frugívoros y nectarívoros (Villareal et al., 2006). Adicional a estas dos familia, la familia Clusiaceae también posee un alto valor trófico para el ecosistema terrestre de isla Gorgona, toda vez que en la isla hay 14 especies, que son aprovechadas por los monos y las aves que la habitan.
Los resultados de este trabajo de investigación concuerdan con lo reportado por Rangel (1990a, 1990b). En términos generales la vegetación de Gorgona puede ser dividida en dos grandes grupos: una vegetación ruderal y/o intervenida y una vegetación natural. La vegetación intervenida comprende las zonas de cultivo que van a lo largo de la isla, cerca de la playa en Palmeras, en el acueducto y en la playa de Yundigüa donde predominan las palmas de coco, palma africana, como estrato arboreo. La vegetación intervenida también incluye vegetación secundaria por claros donde predominan C. spathulata, C. garciae, Vismia baccifera, y Pentagonia spathulata en el estrato arboreo, en el estrato arbustivo se encuentra Trema micrantha, Tetrorchidium andinum, Erythoxylum panamense, entre otras. Por medio de la gráfica de similaridad (Fig. 6) se puede observar que los transectos se agrupan de acuerdo a las caracteristicas de composición o al lugar donde se trasaron los transectos.
La vegetación natural comprende a tres asociaciones y dos comunidades (Fig. 6) (Rangel, 1990b; 1990c), asemejándose el sector de casa de Palmeras, la Mancora y la camaronera a la comunidad Simphonia globulifera-Hyeronima oblonga y Terminalia amozonia y a la asociación vegetal Cespedesio-Symphonion Globuliferae, donde en el estrato arboreo predomina C. spathulata, Symphonia globulosa, Apeiba glabra, el estrato arbustivo está dominado por Solanum sp3, Margaritaria sp. y Eugenia sp1, y el estrato herbaceo está dominado por el género Dieffenbachia sp. Los transectos del sector hacia Yundigüa y los transectos dispuestos por el sendero hacia Palmeras se encuentran o se asemejan a las asociaciones Cespedesio-Symphonion Globuliferae y Ossaeo sessilifoliae Anaxagoretum, predominando en el estrato arbóreo C. spathulata, Ossaea sp., Lacistema aggregatum y en el arboreo inferior predomina M. occidentalis, en el estrato arbustivo predominó Psychotria sp. Es importante destacar que en estas localidades, se registró abundantemente M. occidentalis y L. aggregatum, especies que de acuerdo con Rangel (1990c) no fueron tan importante para estas asociaciones vegetales. Los transectos del Mirador, y los cerros Trinidad y los Micos se asemejaron a la asociación Cassipoureo Ellipticae-Ryanetum speciosae. La vegetación de esta asociación se caracteriza por establecerse en pendientes promedio de 20° y a una altura sobre el nivel del mar entre 220 y 300m. La vegetación predominante del estrato arboreo fue C. spathulata, L. aggregatum, Protium sp. y M. occidentalis, mientras que en el estrato arboreo inferior predominaron Annona glabra, Tovomita sp., y en el estrato arbustivo las especies más abundantes fueron Solanum sp 2 y Trianaeopiper garciae.
Los suelos que son permanentemente lavados, como sucede en Gorgona, tienden a tener bajo contenido de nutrientes (Faber-Langendoen & Gentry, 1991), por lo que no pueden sostener árboles de gran tamaño, favoreciendo la presencia de una alta densidad de arboles y arbustos de tallos pequeños. En este tipo de ambientes, adoptar una estrategia de crecimiento rápido, que favorece la altura antes que el porte del tronco, puede ser ventajoso, como lo presenta C. spathulata en isla Gorgona. Adicionalmente, en el estrato inferior del bosque se encuentran una gran cantidad de hierbas y plántulas de especies de árboles tolerantes a la sombra. Por lo general sus hojas son delgadas, con bajo volumen de células por unidad de área y con un número también bajo de cloroplastos por unidad de área. Esta concentración reducida ayuda a mantener bajos los “costos” metabólicos de construcción foliar y de mantenimiento de las funciones fisiológicas (Sims & Pearcy, 1989; Chazdon & Kauffmann, 1993, citado por Chazdon & Montgomery, 2003), estando adaptadas para aprovechar principalmente los destellos de luz.
Considerar en esta investigación individuos con clases diamétricas (DAP) mayores o igual a 1cm, provocó el incrementó de los registros de más hábitos de crecimiento, como hierbas, hemiepífitas, lianas, lo que conlleva a un incremento en la riqueza de especies. De acuerdo con los resultados de este trabajo, se hace necesario incrementar el esfuerzo de muestreo para alcanzar una adecuada representación de la diversidad vegetal de isla Gorgona. Para esto es necesario considerar no solo realizar campañas en la región noroccidental de la isla, la cual no estuvo representada en este trabajo, sino considerar la necesidad de realizar los esfuerzos de muestreo por lo menos durante las temporadas de mayor y menor precipitación en la zona, con el propósito de tener la mayor cantidad de registros de individuos fértiles, condición requerida para tener una aceptable resolución taxonómica.
Sin duda, las perturbaciones causadas por la extracción de árboles para leña durante la época de la prisión y las caidas de árboles por procesos naturales actualmente, podrían estar modulando la riqueza y diversidad de especies vegetales presentes en isla Gorgona, al incrementar la heterogeneidad del bosque por la presencia de claros y aumentar la disponibilidad de bordes de bosque. De otro lado, Gentry (1982), planteó que hay una relación positiva entre precipitación y la diversidad de especies vegetales, de tal manera que a medida que se incrementa el registro medio de precipitación la diversidad de especies vegetales va creciendo hasta cuando se alcanzan los 4000-4500mm de lluvia, momento en el cual la diversidad vegetal tiende a estabilizarse. Adicionalmente, la abundante presencia de corrientes de agua en isla Gorgona, favorecen el desarrollo de vegetación riparia, incrementando la riqueza general de especies.
La similitud en la estructura de la vegetación entre los sitios de muestreo en isla Gorgona (Fig. 6) parecen estar relacionadas con el grado de perturbación por actividades humanas pasadas y presentes. En este sentido, la vegetación en zonas cercanas al poblado y a las playas como Palmeras y Yondigua, se caracterizaron por tener introducidas y de cultivo. Los bordes de camino o claros del bosque, se encuentran inmersos en procesos de sucesión, encontrándose especies pioneras como C. sphatulata, L. aggregatum, entre otros. Mientras que la zona de los cerros, parece tener una estructura vegetal más diversa y una menor perturbación, aunque también en estos sectores se registran especies pioneras de árboles y arbustos a lo largo de los senderos. Bajo estas características, se puede concluir que la vegetación terrestre del PNN Gorgona corresponde en su mayoría a bosque secundario con restos de bosque primario, concentradas hacia el sector de las partes altas de los cerros, predominando árboles pequeños a medianos, con la presencia de gran cantidad de arbustos.
Por su carácter insular, es prioritario evaluar en isla Gorgona las relaciones ecológicas particulares entre animales y plantas de esta localidad. Preguntas como ¿cuál es el mecanismo que utilizan las plantas que en otros sitios dependen de mamíferos terrestres para dispersar sus semillas, ante la ausencia de este grupo en isla Gorgona?, o ¿Cuál es el papel del mono cariblanco (Cebus capucinus curtus) o el perezoso de tres dedos (Bradypus variegatus gorgon) en los procesos de estructuración de la comunidad vegetal en isla Gorgona?, son preguntas que deben ser abordadas en el corto tiempo, con el propósito de estructurar y fortalecer las estrategias de manejo de esta área protegida.
En conclusión, la vegetación del PNN Gorgona tiene un grado particular de importancia no solo por su alta diversidad de especies, sino porque de ella depende la calidad del hábitat, el aporte de nutrientes al suelo, es la base en la cadena trófica del área terrestre protegida y probablemente, esté aportando materiales y energía al ambiente marino adyacente. Adicionalmente, la presencia de especies vegetales amenazadas o en peligro de extinción, como Podocarpus guatemalensis, convierten a isla Gorgona como un enclave natural de reserva del acervo génico.
Agradecimientos
A Elizabeth Sánchez, Milena Astorquiza y Víctor Eduardo Calero R. por toda su colaboración en campo. A Carlos A. Cultid por su apoyo para los análisis estadísticos. Al Grupo de Investigación en Ecología Animal de la Universidad del Valle por la invitación a participar en el componente de Mamíferos Arbóreos del proyecto «Evaluación del estado actual de los objetos de conservación faunísticos en isla Gorgona: una aproximación holística a la valoración ecológica de PNN Gorgona», en el marco del cual se realizó la presente investigación. Este proyecto fue financiado por la Universidad del Valle, la Fundación Squalus, el Fondo para la Acción Ambiental y la Niñez, Conservación Internacional Colombia y Parques Nacionales Naturales, amparado por el permiso de investigación PIBD-DTSO 011-10.
Referencias
Brunic, E. F.(1983). Vegetation structure and growth. In F. B. Golley (Ed.), Tropical rain forest ecosystems. Estructure and function (pp. 49-73). Elsevier, New York. [ Links ]
Cabrera, I. (1983). Inventario florístico preliminar de la Isla Gorgona. Boletín del mes. 11-20. [ Links ]
Chao, A. & Jost, L. (2012). Coverage-based rarefaction and extrapolation: standardizing samples by completeness rather than size. Ecology, 93(12): 2533-2547. [ Links ]
Chazdon, R. L. & Kauffmann, S. (1993). Plasticity of leaf anatomy of two rain forest shrubs in relation to photosynthetic light acclimation. Functional. Ecology, 7: 385-394. [ Links ]
Chazdon, R. L. & Montgomery, R. A. (2003). La adquisición de Carbono en las Plantas. In M. R. Guariguata & G. H. Kattan (Ed.), Ecología y conservación de bosques neotropicales (pp. 225-250). Libro Universitario Regional, Cartago, Costa Rica. [ Links ]
Clausing G. & Renner, S. S. (2001). Molecular phylogenetics of Melastomataceae and Memecylaceae: implications for character evolution. American Journal of Botany, 88: 486–498. [ Links ]
Davis, A. P., Govaerts, R., Diane., Bridson, M., Ruhsam, M.., Moat, J., Brummitt, N. A. (2009). A global assessment of distribution, diversity, endemism, and taxonomic effort in the Rubiaceae. Annals of the Missouri Botanical Garden, 96(1): 68–78. [ Links ]
Faber-Langendoen, D. & Gentry, A. H. (1991). The structure and diversity of rain forests at Bajo Calima, Chocó Region, western Colombia. Biotropica 23: 2-11. [ Links ]
Feisinger, P. (2003). El diseño de estudios de campo para la conservación de la biodiversidad. Editorial FAN, Santa Cruz de la Sierra, Bolivia. [ Links ]
Gentry, A. H. (1982). Patterns of neotropical plant species diversity. Evolution Biology, 15: 1-84. [ Links ]
Gentry, A. H. (1993). A Field guide to the familias and genera of woody plants of Northwest south america (Colombia, Ecuador, Peru) with suplementary notes on herbaceous taxa. Conservation International. Washington, D.C. [ Links ]
Giraldo, A. (2012). Geomorfologia e hidroclimatologia de Isla Gorgona. In A. Giraldo & B. Valencia (Ed.), Isla Gorgona Paraíso de biodiversidad y ciencia (pp. 17-23). Cali, Colombia: Programa editorial Universidad del Valle. [ Links ]
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica, 4: 1-9. [ Links ]
Hsieh, T. C., Ma, K. H., & Chao, A. (2013). iNEXT online: interpolation and extrapolation (Version 1.0) [Software]. Retrieved from http://chao.stat.nthu.edu.tw/blog/software-download/ [ Links ]
Magurran, A. E. (1988). Ecological diversity and its measurement. Princeton University Press, New Jersey. [ Links ]
Melo, O. A. (1994). Estructura y biodiversidad de los bosques húmedos tropicales de colinas bajas del litoral pacífico colombiano. In I Congreso Nacional sobre Biodiversidad. 4 al 7 de diciembre de 1994 (pp. 101-107). Universidad del Valle, Cali. Colombia. [ Links ]
Murillo, M. T., & Lozano, C. (1989). Hacia la realización de una florula del Parque Nacional Natural islas de Gorgona y Gorgonilla Cauca-Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 12: 277-304. [ Links ]
Rangel, J. O. (1990a). Tipos de Vegetación. In J. Aguirre & J. O. Rangel (Eds.), Biota y Ecosistemas de Gorgona (pp. 109-126). Fondo FEN. Bogotá, Colombia. [ Links ]
Rangel, O., Aguirre., J. & Sánchez, R. (1990b). Aspectos estructurales, dinámicos y fisionómicos de la vegetación (Aproximación preliminar). En J. Aguirre, & J.O. Rangel (Eds.). Biota y Ecosistemas de Gorgona (pp. 127-140). Fondo FEN. Bogotá, Colombia. [ Links ]
Rangel., J. O., Fuentes, C. & Uribe, J. (1990c). Aspectos biotipologicos y morfoecologicos de la vegetación. In J. Aguirre & J. O. Rangel (Eds.). Biota y Ecosistemas de Gorgona (pp. 152-169). Fondo FEN. Bogotá, Colombia. [ Links ]
Rangel, J.O. & Rivera-Díaz, O. (2004a). Diversidad y Riqueza de Espermatófitos en el Chocó Biogeográfico. En J.O. Rangel (Ed.), Colombia Diversidad Biótica IV: El Chocó Biogeográfico/Costa Pacífica (pp. 83-104). Instituto de Ciencias Naturales. Bogotá, Colombia [ Links ]
Rangel, J. O. & Arellano, H. (2004b). Clima del Chocó Biogeográfico de Colombia. En J.O. Rangel (Ed.), Colombia Diversidad Biótica IV: El Chocó Biogeográfico/Costa Pacífica (pp. 39-82). Instituto de Ciencias Naturales. Bogotá, Colombia. [ Links ]
Sims, D. A., & Pearcy, R. W. (1989). Photosynthetic characteristics of a tropical forest understory herb, Alocasia macrorrhiza, and related crop species, Colocasia esculenta grown in contrasting light environment. Oecologia, 79:53-59. [ Links ]
UAESPNN. (1998). El sistema de Parques Nacionales Naturales de Colombia. Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales, Ministerio del Medio Ambiente. Editorial Nomos, Bogotá, Colombia. [ Links ]
UNESCO. (1980). Ecosistemas de los bosques tropicales. Organización de las Naciones Unidas para la Educación, la Cultura y la Ciencia. Madrid. [ Links ]
Vásquez-Vélez, A. I., Cortez, L., Calero, V. E., Soto, E. & Torres, A. M. (2012). Vegetación de la Isla Gorgona. In A. Giraldo & B. Valencia (Eds.), Isla Gorgona Paraíso de biodiversidad y ciencia. Cali, Colombia: Programa editorial Universidad del Valle. [ Links ]
Vásquez-Vélez, A. I., Garzón, S. & Ascencio-Santofinio, H. (2013). Caracterización florística asociada al hábitat de Dendrobatidae (Amphibia:Anurade), en la localidad de Piangüita (Bahía de Buenaventura), Pacifico colombiano. Boletín Científico Museos de Histroia Natural, 17(1): 17-32. [ Links ]
Vélez, G., & Fresneda, H. (1992). Diversidad florística en las comunidades Robledal y Rastrojo Alto, en la cuenca de la quebrada Piedras Blancas, Antioquia. Revista Facultad Nacional de Agronomía Medellín, 45: 3-27. [ Links ]
Villareal H., Alvarez, M., Córdoba, S., Escobar, F., Fagua, G., Gast, F., Mendoza, H., Ospina M., & Umaña, A. M. (2006). Manual de métodos para el desarrollo de inventarios de biodiversidad. Programa de inventarios de biodiversidad. Instituto de investigación de recursos biológicos Alexander von Humboldt. Bogotá, Colombia. [ Links ]
Cabrera, I. (1983). Inventario florístico preliminar de la Isla Gorgona. Boletín del mes. 11-20. [ Links ]
Chao, A. & Jost, L. (2012). Coverage-based rarefaction and extrapolation: standardizing samples by completeness rather than size. Ecology, 93(12): 2533-2547. [ Links ]
Chazdon, R. L. & Kauffmann, S. (1993). Plasticity of leaf anatomy of two rain forest shrubs in relation to photosynthetic light acclimation. Functional. Ecology, 7: 385-394. [ Links ]
Chazdon, R. L. & Montgomery, R. A. (2003). La adquisición de Carbono en las Plantas. In M. R. Guariguata & G. H. Kattan (Ed.), Ecología y conservación de bosques neotropicales (pp. 225-250). Libro Universitario Regional, Cartago, Costa Rica. [ Links ]
Clausing G. & Renner, S. S. (2001). Molecular phylogenetics of Melastomataceae and Memecylaceae: implications for character evolution. American Journal of Botany, 88: 486–498. [ Links ]
Davis, A. P., Govaerts, R., Diane., Bridson, M., Ruhsam, M.., Moat, J., Brummitt, N. A. (2009). A global assessment of distribution, diversity, endemism, and taxonomic effort in the Rubiaceae. Annals of the Missouri Botanical Garden, 96(1): 68–78. [ Links ]
Faber-Langendoen, D. & Gentry, A. H. (1991). The structure and diversity of rain forests at Bajo Calima, Chocó Region, western Colombia. Biotropica 23: 2-11. [ Links ]
Feisinger, P. (2003). El diseño de estudios de campo para la conservación de la biodiversidad. Editorial FAN, Santa Cruz de la Sierra, Bolivia. [ Links ]
Gentry, A. H. (1982). Patterns of neotropical plant species diversity. Evolution Biology, 15: 1-84. [ Links ]
Gentry, A. H. (1993). A Field guide to the familias and genera of woody plants of Northwest south america (Colombia, Ecuador, Peru) with suplementary notes on herbaceous taxa. Conservation International. Washington, D.C. [ Links ]
Giraldo, A. (2012). Geomorfologia e hidroclimatologia de Isla Gorgona. In A. Giraldo & B. Valencia (Ed.), Isla Gorgona Paraíso de biodiversidad y ciencia (pp. 17-23). Cali, Colombia: Programa editorial Universidad del Valle. [ Links ]
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica, 4: 1-9. [ Links ]
Hsieh, T. C., Ma, K. H., & Chao, A. (2013). iNEXT online: interpolation and extrapolation (Version 1.0) [Software]. Retrieved from http://chao.stat.nthu.edu.tw/blog/software-download/ [ Links ]
Magurran, A. E. (1988). Ecological diversity and its measurement. Princeton University Press, New Jersey. [ Links ]
Melo, O. A. (1994). Estructura y biodiversidad de los bosques húmedos tropicales de colinas bajas del litoral pacífico colombiano. In I Congreso Nacional sobre Biodiversidad. 4 al 7 de diciembre de 1994 (pp. 101-107). Universidad del Valle, Cali. Colombia. [ Links ]
Murillo, M. T., & Lozano, C. (1989). Hacia la realización de una florula del Parque Nacional Natural islas de Gorgona y Gorgonilla Cauca-Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 12: 277-304. [ Links ]
Rangel, J. O. (1990a). Tipos de Vegetación. In J. Aguirre & J. O. Rangel (Eds.), Biota y Ecosistemas de Gorgona (pp. 109-126). Fondo FEN. Bogotá, Colombia. [ Links ]
Rangel, O., Aguirre., J. & Sánchez, R. (1990b). Aspectos estructurales, dinámicos y fisionómicos de la vegetación (Aproximación preliminar). En J. Aguirre, & J.O. Rangel (Eds.). Biota y Ecosistemas de Gorgona (pp. 127-140). Fondo FEN. Bogotá, Colombia. [ Links ]
Rangel., J. O., Fuentes, C. & Uribe, J. (1990c). Aspectos biotipologicos y morfoecologicos de la vegetación. In J. Aguirre & J. O. Rangel (Eds.). Biota y Ecosistemas de Gorgona (pp. 152-169). Fondo FEN. Bogotá, Colombia. [ Links ]
Rangel, J.O. & Rivera-Díaz, O. (2004a). Diversidad y Riqueza de Espermatófitos en el Chocó Biogeográfico. En J.O. Rangel (Ed.), Colombia Diversidad Biótica IV: El Chocó Biogeográfico/Costa Pacífica (pp. 83-104). Instituto de Ciencias Naturales. Bogotá, Colombia [ Links ]
Rangel, J. O. & Arellano, H. (2004b). Clima del Chocó Biogeográfico de Colombia. En J.O. Rangel (Ed.), Colombia Diversidad Biótica IV: El Chocó Biogeográfico/Costa Pacífica (pp. 39-82). Instituto de Ciencias Naturales. Bogotá, Colombia. [ Links ]
Sims, D. A., & Pearcy, R. W. (1989). Photosynthetic characteristics of a tropical forest understory herb, Alocasia macrorrhiza, and related crop species, Colocasia esculenta grown in contrasting light environment. Oecologia, 79:53-59. [ Links ]
UAESPNN. (1998). El sistema de Parques Nacionales Naturales de Colombia. Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales, Ministerio del Medio Ambiente. Editorial Nomos, Bogotá, Colombia. [ Links ]
UNESCO. (1980). Ecosistemas de los bosques tropicales. Organización de las Naciones Unidas para la Educación, la Cultura y la Ciencia. Madrid. [ Links ]
Vásquez-Vélez, A. I., Cortez, L., Calero, V. E., Soto, E. & Torres, A. M. (2012). Vegetación de la Isla Gorgona. In A. Giraldo & B. Valencia (Eds.), Isla Gorgona Paraíso de biodiversidad y ciencia. Cali, Colombia: Programa editorial Universidad del Valle. [ Links ]
Vásquez-Vélez, A. I., Garzón, S. & Ascencio-Santofinio, H. (2013). Caracterización florística asociada al hábitat de Dendrobatidae (Amphibia:Anurade), en la localidad de Piangüita (Bahía de Buenaventura), Pacifico colombiano. Boletín Científico Museos de Histroia Natural, 17(1): 17-32. [ Links ]
Vélez, G., & Fresneda, H. (1992). Diversidad florística en las comunidades Robledal y Rastrojo Alto, en la cuenca de la quebrada Piedras Blancas, Antioquia. Revista Facultad Nacional de Agronomía Medellín, 45: 3-27. [ Links ]
Villareal H., Alvarez, M., Córdoba, S., Escobar, F., Fagua, G., Gast, F., Mendoza, H., Ospina M., & Umaña, A. M. (2006). Manual de métodos para el desarrollo de inventarios de biodiversidad. Programa de inventarios de biodiversidad. Instituto de investigación de recursos biológicos Alexander von Humboldt. Bogotá, Colombia. [ Links ]
1. Herbario CUVC; Programa de Postgrado en Ciencias Biología, Departamento de Biología, Facultad de Ciencias Naturales y Exactas, Universidad del Valle, Cali, Colombia; anaisabelvasvel@gmail.com
Recibido 18-X-2013. Corregido 20-XI-2013. Aceptado 19-XII-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}