










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.4 San José Oct./Dec. 2014
Composición de especies leñosas en comunidades invadidas en montañas del centro de Argentina: su relación con factores ambientales locales
Abstract
Woody species composition in invaded communities from mountains of central Argentina: their relations with local environmental factors. Invasions by exotic woody species are threatening ecosystem functions worldwide. The spread and subsequent replacement of native forest by exotic dominated stands is particularly evident nearby urban centers were exotic propagule pressure is highest. Yet, there is a lack of information on the environmental factors that underlie these replacements. In this study we addressed the following questions: (1) is there a local spatial segregation between the dominant native and exotic woody species? and (2) if this local segregation does exist, is it driven by environmental features?. For this, in 2010 we established 31 plots distributed along 16 sampling sites where we surveyed the composition and abundance of all woody species with a basal diameter ≥ 5cm. To characterize the environment of each plot, we measured the topographic position (slope, exposure) and different properties such as soil physics (bulk density, soil impedance), structure (soil deep, texture) and chemical characteristics (pH, nutrient and water content). Through a cluster analysis we were able to identify five different woody communities in coexistence: (1) Woodlands dominated by the exotic Ligustrum lucidum; (2) Mixed woodlands dominated by the native Lithraea molleoides and the exotic Celtis australis; (3) Scrublands dominated by the native Condalia buxifolia; (4) Scrublands dominated by the exotic Cotoneaster glaucophyllus, and (5) Scrubby grasslands with the exotic Pyracantha angustifolia. These communities were all associated with different local topographic and edaphic features. The environmental segregation among the identified communities suggests that woody invaders have the potential to colonize almost all the environments of the study site (though varying in the identity of the dominant exotic species). The observed patterns, even being restricted to a single well invaded area of mountain Chaco, may posit the spread of woody invaders towards native communities in the region.
Key words: biological invasions, cluster analysis, environmental segregation, Mountain Chaco, subtropical forest, woody aliens.
Resumen
En todo el mundo, las invasiones de especies leñosas exóticas están amenazando las funciones ecosistémicas. La dispersión y el subsecuente reemplazo de bosques de especies nativas por comunidades dominadas por exóticas es evidente, particularmente, en proximidad a centros urbanos donde la presión de propágulos de especies exóticas es alta. Sin embargo, existe una falta de información sobre los factores ambientales que subyacen este reemplazo. En este estudio nos propusimos responder las siguientes preguntas: (1) ¿se observa una segregación espacial a escala local entre especies leñosas exóticas y/o nativas dominantes?, y (2) si existe esa segregación, ¿está asociada a variables ambientales? En el 2010 se establecieron 31 parcelas distribuidas en 16 laderas en los bosques Chaqueños de las Sierras de Córdoba, en Argentina central. En cada uno de los sitios de muestreo se relevó la composición y abundancia de todas las especies leñosas con un diámetro a la altura de la base superior a 5cm. Para caracterizar el ambiente en cada una de las parcelas medimos la posición topográfica (pendiente y orientación) y algunas propiedades asociadas con la física (densidad aparente y compactación), estructura (profundidad y textura) y con la química (pH y el contenido de nutrientes y agua) del suelo. A través, de un análisis jerárquico de agrupamiento, se identificaron cinco comunidades de leñosas coexistiendo: (1) Bosques dominados por Ligustrum lucidum, (2) Bosques mixtos dominados por Lithraea molleoides y Celtis australis, (3) Matorrales de Condalia buxifolia, (4) Matorrales de Cotoneaster glaucophyllus, y (5) Pajonales con emergentes de Pyracantha angustifolia. Estas comunidades se asociaron diferencialmente a las variables topográficas y edáficas locales. La segregación ambiental observada sugiere que las especies invasoras tienen una capacidad potencial para colonizar casi todos los ambientes en el área de estudio (variando la identidad de la invasora). En conjunto, los patrones descritos, aunque circunscriptos a un área de Chaco Serrano con un avanzado grado de invasión, plantearían un escenario de posible expansión de las leñosas exóticas sobre las comunidades nativas.
Palabras clave: análisis jerárquicos de agrupamiento, bosque subtropical, Chaco Serrano, invasiones biológicas, leñosas exóticas, segregación ambiental.
En la provincia de Córdoba, el Bosque Chaqueño Serrano adquiere gran importancia, no sólo porque contiene la mitad de especies de plantas vasculares de toda la provincia (Zuloaga, Morrone, & Belgrado, 2008; Giorgis, 2011), sino también porque un 13.5% de su flora corresponde a taxones endémicos del centro de Argentina (Giorgis et al., 2011a). A su vez, provee importantes servicios ecosistémicos, particularmente la provisión de agua, ya que los territorios que ocupa forman parte de las cuencas de los principales ríos que abastecen al menos al 80% de la población de la provincia. No obstante, la vegetación del Bosque Chaqueño Serrano se encuentra amenazada debido a cambios en el uso del suelo, los cuales se incrementaron notablemente desde tiempos prehispánicos (Schofield & Bucher, 1986) hasta la actualidad (Zak, Cabido, & Hodgson, 2004; Brown, Martínez-Ortiz, Acerbi, & Corcuera, 2006).
Sumado a la deforestación, uno de los principales factores de cambio de la vegetación en el área es la invasión de especies leñosas exóticas (Hoyos et al., 2010; Giorgis et al., 2011a, b). Particularmente, la especie invasora Ligustrum lucidum (“siempreverde”) ha experimentado una explosión demográfica en los últimos años, constituyéndose en la especie dominante en ciertas áreas de las Sierras Chicas (Gavier-Pizarro et al., 2012). Al mismo tiempo, existen otras especies leñosas exóticas inva- diendo el área (Gurvich, Tecco, & Díaz, 2005; Giorgis et al., 2011a; Tecco, Urcelay, Díaz, Cabido, & Pérez-Harguindeguy, 2013), aunque, al menos por el momento, con una menor abundancia regional que Ligustrum lucidum. Se ha observado que algunas de estas invasoras afectan no sólo al reclutamiento, composición y abundancia de especies nativas (Tecco, Gurvich, Díaz, Pérez-Harguindeguy, & Cabido, 2006; Giantomasi, Tecco, Funes, Gurvich, & Cabido, 2008; Hoyos et al., 2010; Giorgis, 2011), sino también a procesos directamente vinculados al ciclado de nutrientes (Furey, Tecco, Pérez-Harguindeguy, Giorgis, & Grossi, 2014) y con el ciclo del agua (Cingolani, Gurvich, Zeballos, & Renison, 2010; Jobbágy, Acosta, & Nosetto, 2013).
A escala regional se ha observado que la invasión de Ligustrum lucidum está fuerte- mente asociada con la proximidad a centros poblados (Gavier-Pizarro, Radeloff, Stewart, Huebner, & Keuler, 2010). Esta relación se debería principalmente a dos factores: por un lado, a la fuerte presión de propágulos que suponen estos centros urbanos y, por otra parte, al disturbio asociado a la urbanización (Colautti, Grigorovich, & MacIsaac, 2006; Pauchard et al., 2009; Simberloff, 2009). En conjunto, estos factores aumentan las probabilidades de colonización y establecimiento de especies exóticas (Simberloff, 2009; Gavier-Pizarro et al., 2010). Este patrón se ha observado también en las Sierras Grandes de Córdoba (Giorgis et al., 2011b) y en otras regiones montañosas del mundo (Pauchard et al., 2009). Si bien existen numerosos estudios que abordan esta problemática a escala regional, no hemos encontrado trabajos que analicen la relación entre la presencia y abundancia de especies leñosas exóticas con variables ambientales que actúan a escala local, tales como la posición topográfica, las características físico-químicas y la humedad del suelo, además de la pendiente y la orientación de las laderas. En sistemas montañosos estos factores son importantes estructuradores de las características de la vegetación, ya que afectan la erosión y deposición de sedimentos, el balance térmico y las condiciones de humedad y fertilidad de los suelos (Clark, Palmer, & Clark, 1999; Bledsoe & Shear, 2000; Cingolani, Cabido, Renison, & Solís, 2003). Es esperable, en consecuencia, que esta heterogeneidad local, propia de ambientes montañosos, condicione la presencia y la abundancia de las diferentes leñosas exóticas y nativas (Davis, Grime, & Thompson, 2000; Kakembo, Rowntree, & Palmer, 2007). En este contexto, adquiere relevancia teórica y aplicada entender los factores que subyacen al reemplazo del bosque nativo por comunidades dominadas por leñosas exóticas. Las Sierras Chicas de Córdoba ofrecen un experimento natural apropiado para poner a prueba los efectos de factores locales sobre la composición de leñosas nativas y exóticas, particularmente en remanentes de vegetación próximos a urbanizaciones y en sitios con una alta presión de propágulos. Por lo tanto, el objetivo del presente trabajo es explorar, a escala local, si existe alguna asociación entre la presencia y abundancia de especies leñosas, tanto exóticas como nativas, con ciertas variables ambientales locales en un área invadida de las Sierras Chicas de Córdoba. Específicamente, en este trabajo nos propusimos responder a las siguientes preguntas: 1) en un área con altas densidades de leñosas exóticas, ¿se observa una segregación espacial a escala local entre especies leñosas exóticas y/o nativas dominantes?, y 2) si existe esa segregación, ¿está asociada a variables ambientales?
Materiales y Métodos
Área de estudio y diseño experimental: El área de estudio forma parte de la Estancia Los Potreros (31º07’00”-31º07´31”S - 64º23’191”- 64º22’59” W), ubicada en las Sierras Chicas de Córdoba, cerca de la localidad de Salsipuedes. El área comprende 280ha y se encuentra entre los 900 y 1 000msnm, correspondiendo al piso del Bosque Serrano dentro del Distrito Chaqueño Serrano (Cabrera, 1976). El clima es templado cálido con una estación seca en los meses más fríos. La temperatura media anual es de 15°C, y la precipitación media anual de 850mm, con concentración estival de las lluvias (de Fina, 1992). La vegetación del área de estudio se compone de un mosaico de fisonomías en distintos estados sucesionales, con bosques abiertos a cerrados, matorrales y, en menor medida, pastizales inducidos por fuego y pastoreo (Cabido & Zak, 1999; Gavier-Pizarro & Bucher, 2004). Además, este mosaico de fisonomías vegetales se encuentra sometido a distintos grados de invasión por plantas exóticas (Giorgis, 2011; Giorgis et al., 2011a). El área de estudio constituye un escenario apropiado para abordar nuestro objetivo, ya que la proximidad a los centros urbanizados en las Sierras Chicas ha propiciado una alta presión de propágulos de leñosas exóticas. En consecuencia, la distribución de las especies exóticas en el área de estudio no estaría limitada por una falta de propágulos sino que podría estarlo por las interacciones bióticas o los requerimientos ambientales propios de cada especie. Esto permite abarcar un elenco de especies leñosas que, no sólo es representativo de las principales invasoras de la región (Giorgis et al., 2011a; Tecco et al., 2013), sino que coexiste en un área acotada, afectada por un macroclima y variables regionales homogéneas, en la cual se puede indagar sobre su relación con variables ambientales a escala local.
Se establecieron 32 parcelas permanentes de 10×20m distribuidas en 16 laderas. Una de las parcelas se perdió por motivos ajenos a esta investigación, por lo cual el número considerado en los análisis corresponde a las 31 parcelas restantes. Las laderas se concentraron dentro de un radio de un kilómetro alrededor del casco principal de la estancia y a unos 500m de distancia promedio entre ellas, ya que el objetivo del estudio fue analizar cuál era el patrón de la vegetación y su relación con las variables ambientales a escala local. La selección de la ubicación de las parcelas no fue al azar, sino que se procuró tener una representación de todas las especies leñosas nativas y exóticas, y de las diferentes fisonomías presentes en el área de estudio (desde bosques bien conser- vados a pastizales dominados por gramíneas en mata, llamados comúnmente pajonales). Además, debido a la gran heterogeneidad de la vegetación dentro de cada ladera (por ej., en una misma ladera se podía encontrar un pajonal y un bosque conservado) se decidió establecer dos parcelas por ladera a los efectos de considerar dicha heterogeneidad. En cada parcela se registró la presencia y número de individuos, y se estimó visualmente el porcentaje de cobertura de todas las especies leñosas. Además, se midió el área basal a todos los individuos que superaran los 5cm de diámetro en la base, durante abril a noviembre de 2010. Para la nomenclatura de las especies se siguió a Zuloaga et al. (2008) y su actualización online (www2.darwin.edu.ar). Las parcelas se describieron fisonómicamente siguiendo una modificación del sistema de clasificación pro- puesto por Foti y Blaney (1994) y el protocolo propuesto por la Organización de las Naciones Unidas para la Educación, la Ciencia y la Cultura (UNESCO, 1973). En este caso, se definieron a priori las distintas fisonomías de acuerdo al porcentaje de cobertura de árboles y arbustos que presentó cada parcela. De esta forma se identificaron cinco tipos de fisonomías: Bosque cerrado, Bosque abierto, Matorral cerrado, Matorral abierto y Pajonal con emergentes leñosos. Con el propósito de identificar distintas comunidades de especies leñosas, las parcelas fueron clasificadas en base a su fisonomía y composición (presencia y abundancia) (Nichols, 1923; Foti & Blaney, 1994). Dado que el relevamiento florístico de las parcelas no contempló otras formas de vida (hierbas, gramíneas, entre otras), los grupos de especies identificados no constituyen “comunidades” en el sentido estricto (Whittaker, 1970; Begon, Harper, & Twonsend, 1999), sino un conjunto de leñosas que se distribuyen en el área de estudio conformando más de un tipo fisonómico. Por practicidad, de aquí en más se hará referencia a estos ensambles de leñosas como “comunidades leñosas”.
Caracterización de factores ambientales a escala local: Las parcelas fueron caracterizadas según su posición topográfica, pendiente y orientación, y distintas variables edáficas relacionadas con la estructura, contenido de agua y concentración de nutrientes en el suelo. En el caso de la orientación, se calcularon dos nuevas variables: la orientación norte-sur (=coseno × orientación; valores positivos indican orientación sur y valores negativos orientación norte), y la orientación este-oeste (=seno × orientación; valores positivos representan una orientación hacia el oeste, y valores negativos una orientación hacia el este). Este método es una modificación de las estimaciones de Cushman y Wallin (2002).
Para caracterizar la estructura del suelo se midió la profundidad, el grado de compactación y la densidad aparente. En los tres casos, para la recolección de las muestras o la toma del dato dentro de cada parcela, se realizaron transectas, de 20m de longitud, distribuidas a una distancia mínima de tres metros entre sí. La profundidad del suelo (cm) se midió en 30 puntos por parcela utilizando una varilla de acero, la cual fue introducida en el perfil del suelo hasta alcanzar la roca madre (Kolb, Alpert, Enters, & Holzapfel, 2002; MacDou- gall, Boucher, Turkington, & Bradfield, 2006). En este caso, se dispusieron tres transectas a lo largo de las cuales, se realizaron 10 mediciones de profundidad de suelo cada dos metros de distancia entre ellas. La compactación (kg/cm2) se midió con un penetrómetro (E-280 Pocket Penetrometer) que registra la dureza de la capa superficial del suelo. Para ello, a lo largo de cuatro transectas, se tomaron 10 mediciones cada dos metros, y en total se realizaron 40 lecturas por parcela. En el caso de la medición de la densidad aparente del suelo (g/cm3), se recolectaron seis muestras de los primeros 10cm del perfil con un cilindro de acero de 10×5cm. En este caso, a lo largo de dos transectas, se recolectaron tres muestras de suelo cada cuatro metros, aproximadamente. Las muestras fueron secadas al aire y luego pasadas por un tamiz de 2mm. Posteriormente, fueron secadas en un horno a 60°C durante tres días hasta peso constante y se procedió a calcular la densidad aparente (Throop, Archer, Monger, & Waltman, 2012). Finalmente, las muestras fueron enviadas al Laboratorio de Edafología de la Facultad de Ciencias Agropecuarias (Universidad Nacional de Córdoba, Argentina), para su caracterización físico-química en cuanto a: textura (Day, 1986), pH (Thomas, 1996), conductividad eléctrica, materia orgánica según la técnica de Walkley & Black (Nelson & Sommers,1996) y nitrógeno total según la técnica de Kjeldahl (Bremner, 1996).
La humedad del suelo se estimó a partir del contenido volumétrico de humedad (%). En cada una de las parcelas, se realizaron dos transectas de 20m, en las cuales, cada dos metros se tomaron 10 mediciones. El equipo utilizado para la medición de humedad fue un Moisture Probe Meter (MPB-160-B del ICT International Pty Ltd), con el cual se tomaron 20 registros por parcela, en total, de los primeros 10cm del perfil del suelo, en los meses de noviembre y diciembre de 2010 y marzo de 2011. En el caso de la humedad, profundidad, densidad aparente y compactación del suelo se calculó una media por parcela.
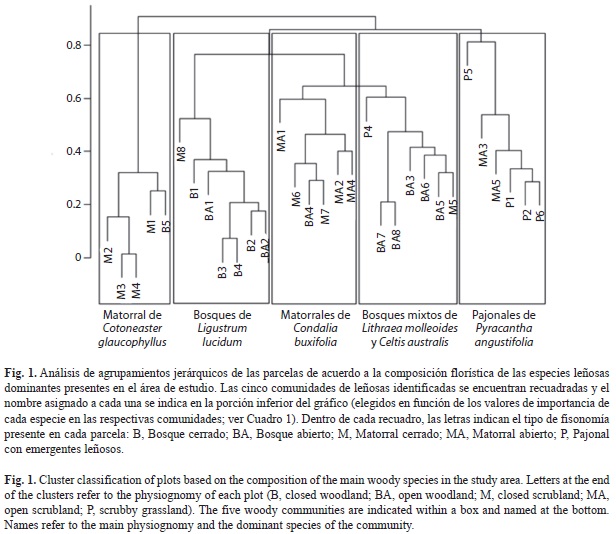
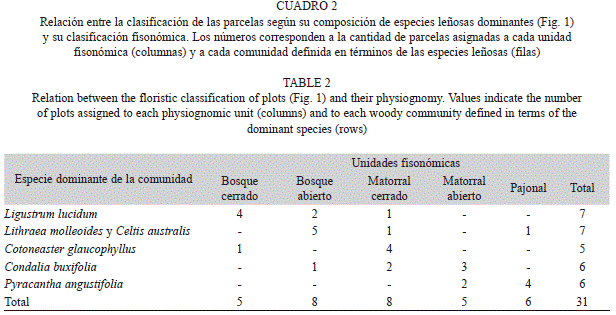
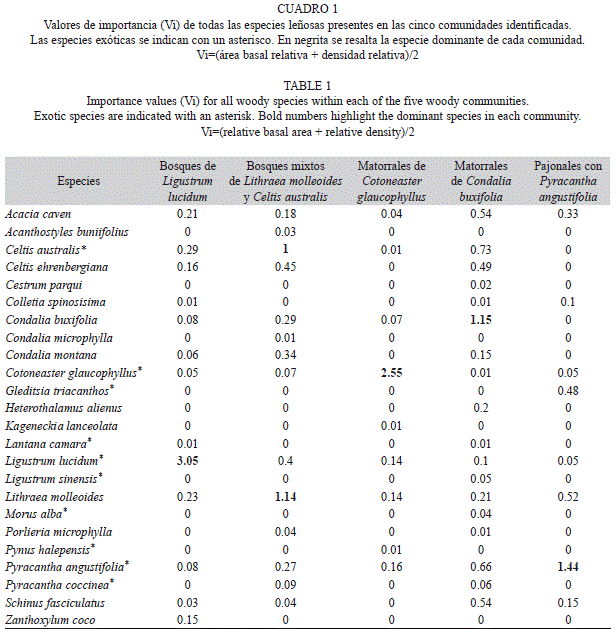
Para cada especie leñosa registrada en el relevamiento, se calculó su valor de importancia (Vi) por parcela. Este índice se obtuvo como una media entre el área basal relativa y la densidad relativa de cada una de las especies (Chen, Hsieh, Jiang, Hsieh, & Sun, 1997; Bledsoe & Shear, 2000). Específicamente el Vi es un indicador del grado de dominancia en cada parcela de cada una de las especies, el cual oscila entre cero y uno. Los valores más cercanos a uno indican que la especie es dominante. A partir de una matriz con los Vi de las especies leñosas por parcela, se realizó un análisis jerárquico de agrupamientos con la finalidad de identificar las principales comunidades de leñosas del área de estudio. Las especies con una única presencia, no se tuvieron en cuenta en el análisis. Para ello se empleó el método de agrupamiento promedio (UPGMA) con el índice de disimilitud o distancia de Bray Curtis. Una vez identificadas las distintas comunidades, se las caracterizó en cuanto a su riqueza de especies leñosas (riqueza total, de nativas y de exóticas) y se identificó la especie leñosa dominante de cada grupo, la cual le dio el nombre a la comunidad. Las especies dominantes se identificaron calculando los Vi dentro de cada comunidad, a partir de la suma de los Vi obtenidos para todas las especies presentes en las parcelas que conformaron cada una de las comunidades. En este caso el Vi, para cada especie, puede ser mayor a uno y, mientras más alto sea su valor, mayor será su grado de dominancia en esa comunidad (Chen et al., 1997; Bledsoe & Shear, 2000). Además, para evaluar si las comunidades leñosas identificadas en el análisis anterior exhibían una fisonomía identificable en particular, se realizó una tabla de contingencia entre las distintas comunidades leñosas y los cinco tipos de fisonomías. El nombre que se le atribuyó a cada comunidad responde, en consecuencia, a la especie y la fisonomía dominantes de cada una (Fig. 1, Cuadro 2).
Por último, se evaluó si las comunidades leñosas identificadas difieren en cuanto a las variables topográficas y edáficas registradas. Debido a que los datos presentaron homogeneidad de varianza, distribución normal e independencia de los errores, se analizaron a través de ANOVA. Además, se utilizó el test HSD Tukey como prueba a posteriori. Todos los análisis estadísticos fueron realiza- dos en R (R Development Core Team, 2012). Para el análisis de clusters se utilizó el paquete vegan (versión 2.0-10) y el paquete stats (versión 3.1.0) para los análisis de la varianza.
Resultados
Identificación y descripción de las comunidades de especies leñosas: En las 31 parcelas censadas se registró un total de 885 individuos pertenecientes a 24 especies (13 familias y 19 géneros) de plantas leñosas, de las cuales 14 son nativas y 10 exóticas (Apéndice). Las familias con más representantes fueron Rhamnaceae y Rosaceae, con cuatro especies cada una.
A partir del análisis jerárquico de agrupamientos, se identificaron cinco comunidades (Fig. 1) que difirieron principalmente en cuanto a la identidad de la especie dominante (Cuadro 1) y a los tipos fisonómicos que con- formaban (Cuadro 2). Algunas comunidades pueden adquirir más de una fisonomía (por ej., la comunidad dominada por Ligustrum lucidum corresponde generalmente a un bosque, pero esporádicamente puede aparecer como un matorral) y viceversa (por ej., tanto bosques abiertos como matorrales abiertos pueden ser parte de la comunidad dominada por Cotoneas- ter glaucophyllus) (Fig. 1, Cuadro 2). A continuación se describen las cinco comunidades de especies leñosas en cuanto al tipo de fisonomía que presentaron, su riqueza de leñosas (riqueza total, de nativas y de exóticas) y la identidad de las especies dominantes y subordinadas más importantes:
Bosques de Ligustrum lucidum, comunidad conformada principalmente por fisonomías de bosque cerrado y, en menor medida, de bosque abierto y matorral cerrado (Fig. 1, Cuadro 2). En esta comunidad se observó una riqueza media de 5.57 especies leñosas, 2.71 exóticas y 2.86 nativas por parcela. La especie exótica L. lucidum es la que presentó el mayor Vi observado no sólo en esta comunidad sino en toda el área de estudio (Cuadro 1). Le siguieron en orden de importancia Celtis australis, Lithraea molleoides y Acacia caven.
Bosques mixtos de Lithraea molleoides y Celtis australis, fisonómicamente conformada por bosques abiertos, aunque se observaron parcelas con coberturas propias de un mato- rral cerrado y pajonal con algunos emergentes arbóreos (Fig. 1, Cuadro 2). En esta comunidad se observó una media de 7.14 especies leñosas y una riqueza media de 2.42 exóticas y 4.71 nativas por parcela. A los altos Vi de la especie nativa, L. molleoides, y la exótica C. australis, le siguieron en importancia las especies Celtis ehrenbergiana, L. lucidum y Condalia montana (Cuadro 1).
Matorrales de Cotoneaster glaucophyllus, estuvo conformada por matorrales cerra- dos, excepto por una parcela con fisonomía de un bosque cerrado (Fig. 1, Cuadro 2). En esta comunidad se observó una riqueza media de 3.2 especies leñosas por parcela y una riqueza media de 2.2 exóticas y 1 nativa. La dominan- cia del arbusto exótico C. glaucophyllus fue seguida en importancia por Pyracantha angus- tifolia, L. molleoides y L. lucidum (Cuadro 1).
Matorrales de Condalia buxifolia, estuvo conformada principalmente por matorrales abiertos, aunque, también adquirió fisonomías propias de un matorral cerrado y de un bosque abierto (Fig. 1; Cuadro 2). Esta comunidad presentó la mayor riqueza de especies, con un total de 8.66 leñosas por parcela, y una riqueza media de 3.33 especies exóticas y 5.33 nativas por parcela. Al arbusto nativo dominante de esta comunidad, C. buxifolia, le siguieron en orden de importancia C. australis, P. angustifolia, A. caven y Schinus fasciculatus (Cuadro 1).
Pajonales con Pyracantha angustifolia, fisonómicamente conformada por pajonales con arbustos aislados y matorrales abiertos (Fig. 1, Cuadro 2). Esta comunidad exhibió una riqueza media de 3.5 especies leñosas por parcela y una riqueza media de 1.83 exóticas y 1.67 nativas. Al arbusto exótico dominante, P. angustifolia, le siguieron en orden de importancia, L. molleoides, Gleditsia triacanthos y A. caven (Cuadro 1).
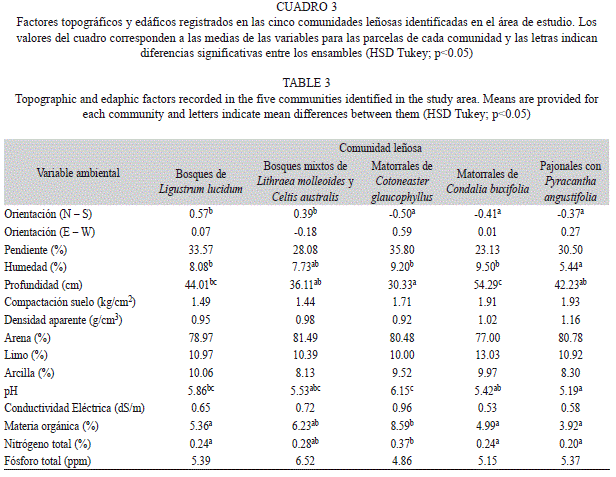
Patrones de distribución de las comunidades leñosas y variables ambientales: Las cinco comunidades leñosas identificadas se asociaron diferencialmente a la orientación N-S de las laderas (F4,26 31=4.35, p≤0.01), y al contenido de humedad (F4,26 31=2.87, p≤0.05), la concentración de nitrógeno (F4,26 31=3.49, p≤0.05) y materia orgánica (F4,26 31=3.51, p≤0.05), el pH (F4,26 31=2.79, p≤0.05) y la profundidad del suelo (F4,26 31=4.53, p≤0.01; Cuadro 3). Los bosques de Ligustrum lucidum y los bosques mixtos de Lithraea molleoides y Celtis australis se asociaron a la orientación sur, mientras las restantes comunidades lo hicieron a la orientación norte (Cuadro 3). Entre las comunidades asociadas a la orien- tación norte, los matorrales de Cotoneaster glaucophyllus se ubicaron sobre suelos menos profundos y con mayor contenido de materia orgánica, nitrógeno y más alto pH (Cuadro 3). Los matorrales de Condalia buxifolia se distribuyeron sobre suelos significativamente más profundos. Finalmente, los pajonales con Pyracantha angustifolia, a pesar de compartir la orientación con las dos comunidades de matorrales, fueron las únicas parcelas asociadas a suelos significativamente más ácidos y con el menor contenido de humedad.
Discusión
El arreglo espacial de las especies leñosas de las Sierras Chicas de Córdoba, junto a la composición florística y la fisonomía de los ensambles, permitieron identificar cinco comunidades principales, algunas de ellas dominadas por especies exóticas. Además, los resultados de este trabajo también indican que la presencia y abundancia de las principales invasoras del Chaco Serrano (Giorgis et al., 2011a) se asocian, a escala local, a condiciones topográficas y edáficas particulares, que difieren parcialmente de aquellas donde se presentan las comunidades dominadas por nativas. Las características ambientales a las cuales estuvieron asociadas las comunidades dominadas por nativas (matorrales y bosques de Condalia buxifolia y Lithraea molleoides), fueron en su mayoría compartidas con alguna comunidad dominada por exóticas, lo que plantearía un escenario posible de avance de las exóticas sobre estos remanentes nativos.
En concordancia con resultados previos disponibles para el área de estudio (Gavier- Pizarro et al., 2010; Giorgis et al., 2011a), y con respecto a las predicciones sobre el efecto de la proximidad a urbanizaciones, nuestros resultados reportan un alto grado de invasión por leñosas en las Sierras Chicas de Córdoba. Cerca del 50% de la flora leñosa registrada correspondió a exóticas, llegando a dominar en casi todas las comunidades identificadas y, si no lo hicieron, fueron las especies subordinadas más importantes en cada comunidad. Esto último es consistente con el patrón de avance de Ligustrum lucidum documentado en las últimas décadas a escala regional (Hoyos et al., 2010; Gavier-Pizarro et al., 2012), pero destaca, además, la importancia de otras leñosas invasoras como Cotoneaster glaucophyllus, Pyracantha angustifolia y Celtis australis. Las comunidades dominadas por especies exóticas se caracterizaron por una baja riqueza de leñosas (1.83 a 5.57 especies por parcela), mientras que aquellas dominadas por especies nativas exhibieron los mayores valores de riqueza (7.14 a 8.6 especies por parcela). Esto aportaría nuevas evidencias a la tendencia general de desplazamiento de las especies nativas y empobrecimiento florístico propio de comunidades invadidas descritas a otras escalas y en otras regiones del mundo (McKinney & Lockwood, 1999; Hejda, Pyšek, & Jarošík, 2009).
La distribución de las especies de plantas y de las comunidades vegetales ha sido tradicionalmente asociada, dentro de una misma región climática, a factores topográficos y edáficos (Woodward, 1987). Nuestros resultados también permiten destacar el efecto de los factores topográficos y edáficos a escala local sobre los patrones de dominancia de las especies exóticas y nativas y, además, la segregación espacial de las comunidades de leñosas en las Sierras Chicas de Córdoba. Las parcelas con comunidades de bosques (tanto las dominadas por la invasora L. lucidum como las codominadas por la nativa L. molleoides y la exótica C. australis), se presentaron preferentemente en laderas con orientación sur. Las laderas orientadas al norte presentaron alternativamente matorrales o pajonales dominados por nativas (C. buxifolia) o por exóticas (C. glaucophyllus o P. angustifolia). La presencia de bosques en laderas expuestas al sur y de matorrales en laderas con orientación al norte, puede deberse a las diferencias ambientales entre ambas situaciones (principalmente mayor insolación y menor humedad en laderas norte). No obstante, no se puede descartar que esta asociación se relacione a la historia del uso del suelo y los regímenes de disturbio en la región (Giorgis, 2011). Por ejemplo, se ha observado que laderas con orientación norte sufren una mayor frecuencia de incendios que laderas sur, por lo que la regeneración de especies arbóreas, tanto nativas como exóticas, se vería retardada (Giorgis, Cingolani, Gurvich, Tecco, & Cabido, 2013). En cuanto a las comunidades leñosas asociadas a la ladera norte, vale destacar las diferencias en variables edáficas entre los sitios ocupados por matorrales dominados por la exótica C. glaucophyllus y los pajonales dominados por el arbusto, también exótico, P. angustifolia. Los primeros se presentan sobre suelos significativamente menos ácidos que los segundos, con mayor contenido de humedad y concentración de nutrientes y de materia orgánica; las parcelas con comunidades dominadas por la nativa C. buxifolia presentan condiciones intermedias. La segregación espacial observada en las distintas comunidades en relación a la topografía, y la asociación de estas a variables edáficas particulares, podrían responder a dos mecanismos no excluyentes operando a escala local: por un lado, la presencia (o ausencia) de una especie invasora en un sitio sería el resulta- do de un filtro ambiental inicial (por ej., variables topográficas particulares estrechamente vinculadas con aspectos microclimáticos y/o disturbio). Por otro lado, si una vez establecida, una especie exótica se torna dominante en el sitio, su impacto sobre procesos ecosistémicos como la descomposición y el ciclo del agua, modificarían características edáficas locales (por ej., contenido de nutrientes, pH, humedad, entre otras). En consecuencia, sería oportuno realizar estudios complementarios evaluando si las diferencias en la calidad edáfica entre las comunidades leñosas corresponden a una condición preexistente o representan, más bien, una consecuencia del establecimiento de la especie invasora que, en este último caso, estaría actuando como un ingeniero ecosistémico (Crooks, 2002).
Estudios como este, en los cuales se analice la relación de las comunidades florísticas con aspectos topográficos y edáficos locales, abarcando la heterogeneidad ambiental propia de los sistemas montañosos, podría arrojar indicios de la dinámica de la vegetación del sistema, particularmente en áreas expuestas a una alta presión de propágulos exóticos (por ej., vegetación natural próxima a regiones urbanizadas; Gavier-Pizarro et al., 2010). De acuerdo a los resultados reportados en este tra- bajo, la dinámica de la vegetación a escala local de parche estaría siendo afectada seriamente por las leñosas invasoras, las cuales estarían desplazando a las especies nativas en nuestra área de estudio. La aparente segregación ambiental observada, no sólo sugiere que las especies invasoras tienen una capacidad potencial para colonizar casi todos los ambientes en las Sierras Chicas de Córdoba, variando la identidad de la invasora, sino que, en algunos casos, al establecerse estarían conformando comunidades dominadas por una única especie leñosa. Este fenómeno se pone de manifiesto al observar que de las cinco comunidades de leñosas descritas, cuatro están dominadas o codominadas por una exótica y, además, se asociaron a condiciones ambientales particulares, conformando tres fisonomías distintas (por ej., fisonomías boscosas dominadas por L. lucidum, matorrales cerrados dominados por C. glaucophyllus y pajonales por P. angustifo- lia). A su vez, las comunidades dominadas por nativas (por ej., C. buxifolia y L. molleoides), no sólo se encontraron invadidas en distinto grado (Cuadro 1), sino que se asociaron a condiciones ambientales similares a alguna de las comunidades dominadas por especies exóticas (Cuadro 3). En conjunto, los patrones descriptos, aunque circunscriptos a un área del Chaco Serrano con un avanzado grado de invasión, son compatibles con un escenario de expansión de las leñosas exóticas sobre los remanentes de bosques y matorrales nativos. En particular, se observó la presencia de L. lucidum en todas las comunidades leñosas (Cuadro 1) y en casi todas las parcelas (datos no mostrados), ya sea como individuos aislados o formando parches de bosques mono específicos. La expansión de L. lucidum hacia los matorrales y pajonales podría, además, verse facilitada por las leño- sas que dominan dichas fisonomías. Estudios previos en la región confirman que los arbustos de P. angustifolia y, en menor medida, los de C. buxifolia, facilitan el reclutamiento y supervivencia de L. lucidum bajo sus copas (Tecco et al., 2006; 2007). Estas evidencias de nodricismo sugieren que, al menos los pajonales y matorrales de P. angustifolia, pero probablemente también aquellos dominados por C. buxifolia y C. glaucophyllus podrían transformarse en bosques de L. lucidum. En conjunto, estos antecedentes, junto a resultados previos que muestran tendencias similares a escala de paisaje (Lichstein, Grau, & Aragón, 2004; Hoyos et al., 2010; Gavier-Pizarro et al., 2012), sugieren para el mediano y largo plazo un progresivo avance y dominancia de L. lucidum sobre todas las comunidades leñosas descritas, tanto las dominadas por leñosas nativas como por otras exóticas.
Dada la creciente problemática ambiental que supone el reemplazo de remanentes de bosque serrano por comunidades dominadas por leñosas invasoras, es importante identificar áreas prioritarias a proteger y restaurar, centrándose en determinados sectores, fisonomías y comunidades florísticas, con el objetivo de garantizar determinados servicios ecosistémicos (por ej., protección de cuencas hídricas). A partir de los patrones descritos en este estudio, sería conveniente que un programa de erradicación de especies exóticas se focalizara en laderas sur, donde L. lucidum tendría, al menos en la actualidad, su mayor desarrollo. De esta manera, al controlar su reclutamiento en estos ambientes, se facilitaría la recuperación de bosques de L. molleoides, al tiempo que se disminuiría la presión de propágulos de L. lucidum desde esos bosques hacia otras áreas que le son menos favorables, como los pajonales y matorrales. Además, la planificación de una restauración del sistema serrano debe tener en cuenta el legado edáfico que conlleva la dominancia de una exótica, aún luego de erradicarla local- mente (Marchante, Kjøller, Struwe, & Freitas, 2009; Grman & Suding, 2010). En este sentido, los impactos ecosistémicos de las invasiones sobre la biota del suelo, el ciclado de nutrientes y otros procesos asociados, son difíciles de predecir (Mack & D’Antonio, 2003; Ehrenfeld, 2003; 2004; Levine et al., 2003; Wolfe & Klironomos, 2005; Strayer, Eviner, Jeschke, & Pace, 2006; Furey et al., 2014), pudiendo comprometer la recolonización de las especies nativas y requerir actividades de enmienda por parte de los restauradores (Marchante et al., 2009; Grman & Suding, 2010).
Es interesante destacar la presencia y dominancia de la exótica C. australis. Ante- cedentes previos en la región han reportado su presencia ocasionalmente (Giorgis et al., 2011a), pero nunca como un elemento dominante de las comunidades serranas. En nuestra área de estudio, ésta fue la única especie invasora que compartió dominancia con leñosas típicas del bosque nativo como L. molleoides. Sería importante estudiar si este comporta- miento se mantiene en el tiempo, o si C. australis eventualmente excluye competitivamente a las especies nativas y logra formar comu- nidades mono específicas. Estas evidencias alertan sobre el potencial invasor de la especie, y podrían estar indicando que C. australis se encontraría en una primera fase de invasión (sensu Mack et al., 2000), lo cual minimiza los costos y aumenta las probabilidades de éxito de un programa de erradicación (Hulme, 2006).
Nuestros resultados indican que detrás del gran aumento en la superficie invadida en la región (Hoyos et al., 2010; Gavier-Pizarro et al., 2012), existe una compleja dinámica en la que interactúan numerosas especies en un ambiente heterogéneo. La capacidad de algunas invasoras leñosas del Bosque Chaqueño Serrano de constituir comunidades dominadas casi por una sola especie, su aparente asociación a condiciones edáficas particulares y su solapamiento ambiental con las leñosas nativas, constituyen un claro indicio de la necesidad de profundizar en el estudio de aspectos relacionados con la dinámica a largo plazo de los patrones observados y su impacto ecosistémico.
Agradecimientos
Agradecemos a Kevin Begg por permitirnos realizar este estudio en su estancia y a tres revisores anónimos que con sus sugerencias contribuyeron a incrementar la calidad del manuscrito. Este trabajo forma parte de la Tesis Doctoral de SZ. A las instituciones que apoya- ron este estudio: la Secretaria de Ciencia y Técnica de la Universidad Nacional de Córdoba, FONCYT (PICT N 903, PICT 953), CONICET (PIP 112-200801-01458, beca doctoral a SZ) y PIO Biodiversidad MINCyT-Córdoba.
Referencias
Begon, M., Harper, J. L., & Townsend, C. R. (1999). Ecología: individuos, poblaciones y comunidades. Barcelona, España: Omega. [ Links ]
Bledsoe, B. P. & Shear, H. T. (2000). Vegetation along hydrological and edaphic gradients in a North Caro- lina coastal plain creek bottom and implications for restoration. Wetlands, 20, 126-147. [ Links ]
Bremner, J. M. (1996). Nitrogen - Total. In D. L. Sparks, A. L. Page, P. A. Helmke, R. H. Loeppert, P. N. Soltanpour, M. A. Tabatabai, C. T. Johnston, & M. E. Sumner (Eds.), Methods of Soil Analysis. Part 3 – Chemical Methods (pp. 961-1010). Madison, Wisconsin: SSSA, CSSA. [ Links ]
Brown, A., Martínez Ortiz, U., Acerbi, M., & Corcuera, J. (Eds.). (2006). La Situación Ambiental Argentina 2005. Buenos Aires, Argentina: Fundación Vida Silvestre Argentina. [ Links ]
Cabido, M. R. & Zak, M. (1999). Vegetación del norte de Córdoba. Córdoba, Argentina: SAGyRR. [ Links ]
Cabrera, A. L. (1976). Regiones fitogeográficas argentinas. In W. F. Kugler (Ed.), Enciclopedia Argentina de Agricultura y Jardinería (pp. 1-85). Buenos Aires, Argentina: ACME. [ Links ]
Chen, Z. S., Hsieh, C. F., Jiang, F. Y., Hsieh, T. H., & Sun, I. F. (1997). Relations of soil properties to topography and vegetation in a subtropical rain forest in southern Taiwan. Plant Ecology, 132, 229-241. [ Links ]
Cingolani, A. M., Cabido, M. R., Renison, D., & Solís Neffa, V. (2003). Combined effects of environment and grazing on vegetation structure in Argentine granite grasslands. Journal of Vegetation Science, 14, 223-232. [ Links ]
Cingolani, A. M., Gurvich, D. E., Zeballos, S. R., & Renison, D. (2010). Sin ecosistemas saludables no hay agua segura. El caso de Córdoba. Revista Única, 111, 48-52. [ Links ]
Clark, D. B., Palmer, M. W., & Clark, D. A. (1999). Eda- phic factors and the landscape-scale distribution of tropical rain forest trees. Ecology, 80, 2662-2675. [ Links ]
Colautti, R. I., Grigorovich, I. A., & MacIsaac, H. J. (2006). Propagule pressure: A null model for biolo- gical invasions. Biological Invasions, 8, 1023-1037. [ Links ]
Crooks, J. A. (2002). Characterizing ecosystem-level consequences of biological invasions: The role of ecosys- tem engineers. Oikos, 97, 153-166. [ Links ]
Cushman, S. A. & Wallin, D. O. (2002). Separating the effects of environmental, spatial and disturbance factors on forest community structure in the Russian Far East. Forest Ecology and Management, 168, 201-215. [ Links ]
Davis, M. A., Grime, J. P., & Thompson, K. (2000). Fluctuating resources in plant communities, a gene- ral theory of invasibility. Journal of Ecology, 88, 528-534. [ Links ]
Day, P. R. (1986). Particle fractionation and particle-size analysis. In A. Klute (Ed.), Methods of Soil Analysis. Part I. Madison, Wisconsin: American Society of agronomy, Soil Science Society of America. [ Links ]
de Fina, A. L. (1992). Aptitud agroclimática de la República Argentina. Buenos Aires, Argentina: Academia Nacional de Agronomía y Veterinaria. [ Links ]
Ehrenfeld, J. G. (2003). Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems, 6, 503-523. [ Links ]
Ehrenfeld, J. G. (2004). The expression of multiple functions in urban forested wetlands. Wetlands, 24, 719-733. [ Links ]
Foti, T. & Blaney, M. (1994). A Classification System for the Natural Vegetation of Arkansas. Proceedings Arkansas Academy of Science, 48, 50-53. [ Links ]
Furey, C., Tecco, P. A., Perez-Harguindeguy, N., Giorgis, M. A., & Grossi, M. (2014). The importance of native and exotic plant identity and dominance on decomposition patterns in mountain woodlands of central Argentina. Acta Oecologica, 54, 13-20. [ Links ]
Gavier-Pizarro, G. I. & Bucher, E. H. (2004). Deforestación de las Sierras Chicas de Córdoba (Argentina) en el período 1970-1997. Córdoba, Argentina: Academia Nacional de Ciencias. [ Links ]
Gavier-Pizarro, G. I., Radeloff, V. C., Stewart, S. I., Huebner, C. D., & Keuler, N. S. (2010). Rural housing is related to plant invasions in forests of southern Wisconsin, USA. Landscape Ecology, 25, 1505-1518. [ Links ]
Gavier-Pizarro, G. I., Kuemmerle, T., Hoyos, L. E., Stewart, S. I., Huebner, C. D., Keuler, N. S., & Rade- loff, V. C. (2012). Monitoring the invasion of an exotic tree Ligustrum lucidum from 1983 to 2006 with Landsat TM/ETM+ satellite data and Support Vector Machines in Córdoba, Argentina. Remote Sensing of Environment, 122, 134-145. [ Links ]
Giantomasi, A., Tecco, P. A., Funes, G., Gurvich, D. E., & Cabido, M. (2008). Canopy effects of the invasive shrub Pyracantha angustifolia on seed bank compo- sition, richness and density in a montane shrubland (Córdoba, Argentina). Austral Ecology, 33, 68-77. [ Links ]
Giorgis, M. A. (2011). Caracterización florística y estructural del Bosque Chaqueño Serrano (Córdoba) en relación a gradientes ambientales y de uso (Tesis de Doctorado). Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Córdo- ba, Argentina. [ Links ]
Giorgis, M. A., Cingolani, A. M., Chiarini, F., Chiapella, J., Barboza, G., Espinar, L. A., Morero, R., Gurvich, D. E., Tecco, P. A., Subils, R., & Cabido, M. (2011a). Composición florística del Bosque Chaqueño serrano de la provincia de Córdoba, Argentina. Kurtziana, 36, 9-43. [ Links ]
Giorgis, M. A., Tecco, P. A., Cingolani, A. M., Renison, D., Marcora P., & Paiaro, V. (2011b). Factors associated with woody alien species distribution in a newly invaded mountain system of central Argentina. Biological Invasions, 13, 1423-1434. [ Links ]
Giorgis, M. A., Cingolani, A. M., Gurvich, D. E., Tecco P. A., & Cabido, M. (2013). ¿Cuán estrecha es la relación entre la fisonomía y la composición florística a lo largo de gradientes ambientales en los “ecosiste- mas inciertos”? Boletín de la Sociedad Argentina de Botánica, suplemento 48. [ Links ]
Grman, E. & Suding, K. N. (2010). Within-Year Soil Legacies Contribute to Strong Priority Effects of Exotics on Native California Grassland Communities. Restoration Ecology, 18, 664-670. [ Links ]
Gurvich, D. E., Tecco, P. A., & Díaz, S. (2005). Plant invasions in undisturbed ecosystems: The triggering attribute approach. Journal of Vegetation Science, 16, 723-728. [ Links ]
Hejda, M., Pyšek, P., & Jarošík, V. (2009). Impact of invasive plants on the species richness, diversity and composition of invaded communities. Journal of Ecology, 97, 393-403. [ Links ]
Hoyos, L. E., Gavier-Pizarro, G. I., Kuemmerle, T., Bucher, E. H., Radeloff, V. C., & Tecco, P. A. (2010). Invasion of glossy privet (Ligustrum lucidum) and native forest loss in the Sierras Chicas of Córdoba, Argentina. Biological invasions, 12, 3261-3275. [ Links ]
Hulme, P. E. (2006). Beyond control: Wider implications for the management of biological invasions. Journal of Applied Ecology, 43, 835-847. [ Links ]
Jobbágy, E. G., Acosta, A. M., & Nosetto, M. D. (2013). Rendimiento hídrico en cuencas primarias bajo pastizales y plantaciones de pino de las sierras de Córdoba (Argentina). Ecología Austral, 23, 87-96. [ Links ]
Kakembo, V., Rowntree, K., & Palmer, A. R. (2007). Topographic controls on the invasion of Pteronia incana (Blue bush) onto hillslopes in Ngqushwa (formerly Peddie) district, Eastern Cape, South Africa. Catena, 70, 185-199. [ Links ]
Kolb, A., Alpert, P., Enters, D., & Holzapfel, C. (2002). Patterns of Invasion within a Grassland Community. Journal of Ecology, 90, 871-881. [ Links ]
Levine, J. M., Vila, M., Antonio, C. M., Dukes, J. S., Grigulis, K., & Lavorel, S. (2003). Mechanisms underlying the impacts of exotic plant invasions. Proceedings of the Royal Society of London, 270, 775-781. [ Links ]
Lichstein, J. W., Grau, H. R., & Aragón, R. (2004). Recruitment limitation in secondary forests domina- ted by an exotic tree. Journal of Vegetation Science, 15, 721-728. [ Links ]
MacDougall, A. S., Boucher, J., Turkington R., & Bradfield, G. E. (2006). Patterns of plant invasion along an environmental stress gradient. Journal of Vegetation Science, 17, 47-56. [ Links ]
Mack, R. N., Simberloff, D., Mark Lonsdale, W., Evans, H., Clout, M., & Bazzaz, F. A. (2000). Biotic inva- sions: causes, epidemiology, global consequences, and control. Ecological Applications, 10, 689-710. [ Links ]
Mack, M. C. & D’Antonio, C. M. (2003). Exotic grasses alter controls over soil nitrogen dynamics in Hawaiian woodland. Ecological Applications, 13, 154-166. [ Links ]
Marchante, E., Kjøller, A., Struwe, S., & Freitas, H. (2009). Soil recovery after removal of the N2-fixing invasive Acacia longifolia: Consequences for ecosystem restoration. Biological Invasions, 11, 813-823. [ Links ]
McKinney, M. L. & Lockwood, J. L. (1999). Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends in Ecology & Evolution, 14, 450-453. [ Links ]
Nelson, D. W. & Sommers, L. E. (1996). Total Carbon, Organic carbon, and Organic Matter. In D. L. Sparks, A. L. Page, P. A. Helmke, R. H. Loeppert, P. N. Soltanpour, M. A. Tabatabai, C. T. Johnston, & M. E. Sumner (Eds.), Methods of Soil Analysis. Part 3 – Chemical Methods (pp. 961-1010). Madison, Wisconsin: ASA, SSSA, CSSA. [ Links ]
Nichols, G. E. (1923). A Working Basis for the Ecological Classification of Plant Communities. Ecology, 4, 154-179. [ Links ]
Organización de las Naciones Unidas para la Educación, la Ciencia y la Cultura [UNESCO]. (1973). Clasificación internacional y cartografía de la vegetación (pp. 1-112). [ Links ]
Pauchard, A., Kueffer, C., Dietz, H., Daehler, C. C., Alexander, J., Edwards, P. J., Arévalo, J. R., Cavieres, L. A., Guisan, A., Haider, S., Jakobs, G., McDougall, K., Millar, C. I., Naylor, B. J., Parks, C. G., Rew, L. J., & Seipel, T. (2009). Ain’t no mountain high enough: plant invasions reaching new elevations. Frontier Ecology Environment, 7, 479-486. [ Links ]
R Development Core Team. (2012). R: A Language and environment for statistical computing. Vienna, Austria: R. Foundation for Statistical Computing. Recuperado de http://www.R-roject.org, Vienna, Austria. [ Links ]
Schofield, C. J. & Bucher, E. H. (1986). Industrial Con- tributions to Desertification in South America. Tree, 1, 78-80. [ Links ]
Simberloff, D. (2009). The Role of Propagule Pressure in Biological Invasions. Annual Review of Ecology Evolution and Systematics, 40, 81-102. [ Links ]
Strayer, D. L., Eviner, V. T., Jeschke, J. M., & Pace, M. L. (2006). Understanding the long-term effects of species invasions. Trends in Ecology & Evolution, 21, 645-651. [ Links ]
Tecco, P. A., Gurvich, D. E., Díaz, S., Pérez-Harguindeguy, N., & Cabido, M. (2006). Positive interaction between invasive plants: The influence of Pyracantha angustifolia on the recruitment of native and exotic woody species. Austral Ecology, 31, 293-300. [ Links ]
Tecco, P. A., Diaz, S., Gurvich, D. E., Perez-Harguindeguy, N., Cabido, M., & Bertone, G. A. (2007). Facilitation and interference underlying the association between the woody invaders Pyracantha angustifolia and Ligustrum lucidum. Applied Vegetation Science, 10, 211-218. [ Links ]
Tecco, P. A., Urcelay, C., Díaz, S., Cabido, M., & Pérez- Harguindeguy, N. (2013). Contrasting functional trait syndromes underlay woody alien success in the same ecosystem. Austral Ecology, 38, 443-451. [ Links ]
Thomas, G. W. (1996). Soil pH and soil acidity. In D. L. Sparks, A. L. Page, P. A. Helmke, R. H. Loeppert, P. N. Soltanpour, M. A. Tabatabai, C. T. Johnston, & M. E. Sumner (Eds.), Methods of Soil analysis. Part 3 – Chemical Methods (Capítulo 16). Madison, Wisconsin: ASA, SSSA, CSSA. [ Links ]
Throop, H. L., Archer, S. R., Monger, H. C., & Waltman, S. (2012). When bulk density methods matter: implications for estimating soil organic carbon pools in rocky soils. Journal of Arid Environments, 77, 66-71. [ Links ]
Whittaker, R. H. (1970). Communities and ecosystems. New York: McMillan. [ Links ]
Wolfe, B. E. & Klironomos, J. N. (2005). Breaking new ground: soil communities and exotic plant invasion. Bioscience, 55, 477-487. [ Links ]
Woodward, F. I. (1987). Climate and plant distribution. Cambridge: Cambridge University Press. [ Links ]
Zak, M. R., Cabido, M., & Hodgson, J. G. (2004). Do subtropical seasonal forest in the Gran Chaco, Argentina, have a future? Biological Conservation, 120, 589-598. [ Links ]
Zuloaga, F. O., Morrone, O., & Belgrano, M. J. (2008). Catalogue of the vascular plants of the southern cone (Argentina, southern Brazil, Chile, Paraguay and Uruguay). Missouri: Missouri Botanical Garden Press. [ Links ]
Bledsoe, B. P. & Shear, H. T. (2000). Vegetation along hydrological and edaphic gradients in a North Caro- lina coastal plain creek bottom and implications for restoration. Wetlands, 20, 126-147. [ Links ]
Bremner, J. M. (1996). Nitrogen - Total. In D. L. Sparks, A. L. Page, P. A. Helmke, R. H. Loeppert, P. N. Soltanpour, M. A. Tabatabai, C. T. Johnston, & M. E. Sumner (Eds.), Methods of Soil Analysis. Part 3 – Chemical Methods (pp. 961-1010). Madison, Wisconsin: SSSA, CSSA. [ Links ]
Brown, A., Martínez Ortiz, U., Acerbi, M., & Corcuera, J. (Eds.). (2006). La Situación Ambiental Argentina 2005. Buenos Aires, Argentina: Fundación Vida Silvestre Argentina. [ Links ]
Cabido, M. R. & Zak, M. (1999). Vegetación del norte de Córdoba. Córdoba, Argentina: SAGyRR. [ Links ]
Cabrera, A. L. (1976). Regiones fitogeográficas argentinas. In W. F. Kugler (Ed.), Enciclopedia Argentina de Agricultura y Jardinería (pp. 1-85). Buenos Aires, Argentina: ACME. [ Links ]
Chen, Z. S., Hsieh, C. F., Jiang, F. Y., Hsieh, T. H., & Sun, I. F. (1997). Relations of soil properties to topography and vegetation in a subtropical rain forest in southern Taiwan. Plant Ecology, 132, 229-241. [ Links ]
Cingolani, A. M., Cabido, M. R., Renison, D., & Solís Neffa, V. (2003). Combined effects of environment and grazing on vegetation structure in Argentine granite grasslands. Journal of Vegetation Science, 14, 223-232. [ Links ]
Cingolani, A. M., Gurvich, D. E., Zeballos, S. R., & Renison, D. (2010). Sin ecosistemas saludables no hay agua segura. El caso de Córdoba. Revista Única, 111, 48-52. [ Links ]
Clark, D. B., Palmer, M. W., & Clark, D. A. (1999). Eda- phic factors and the landscape-scale distribution of tropical rain forest trees. Ecology, 80, 2662-2675. [ Links ]
Colautti, R. I., Grigorovich, I. A., & MacIsaac, H. J. (2006). Propagule pressure: A null model for biolo- gical invasions. Biological Invasions, 8, 1023-1037. [ Links ]
Crooks, J. A. (2002). Characterizing ecosystem-level consequences of biological invasions: The role of ecosys- tem engineers. Oikos, 97, 153-166. [ Links ]
Cushman, S. A. & Wallin, D. O. (2002). Separating the effects of environmental, spatial and disturbance factors on forest community structure in the Russian Far East. Forest Ecology and Management, 168, 201-215. [ Links ]
Davis, M. A., Grime, J. P., & Thompson, K. (2000). Fluctuating resources in plant communities, a gene- ral theory of invasibility. Journal of Ecology, 88, 528-534. [ Links ]
Day, P. R. (1986). Particle fractionation and particle-size analysis. In A. Klute (Ed.), Methods of Soil Analysis. Part I. Madison, Wisconsin: American Society of agronomy, Soil Science Society of America. [ Links ]
de Fina, A. L. (1992). Aptitud agroclimática de la República Argentina. Buenos Aires, Argentina: Academia Nacional de Agronomía y Veterinaria. [ Links ]
Ehrenfeld, J. G. (2003). Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems, 6, 503-523. [ Links ]
Ehrenfeld, J. G. (2004). The expression of multiple functions in urban forested wetlands. Wetlands, 24, 719-733. [ Links ]
Foti, T. & Blaney, M. (1994). A Classification System for the Natural Vegetation of Arkansas. Proceedings Arkansas Academy of Science, 48, 50-53. [ Links ]
Furey, C., Tecco, P. A., Perez-Harguindeguy, N., Giorgis, M. A., & Grossi, M. (2014). The importance of native and exotic plant identity and dominance on decomposition patterns in mountain woodlands of central Argentina. Acta Oecologica, 54, 13-20. [ Links ]
Gavier-Pizarro, G. I. & Bucher, E. H. (2004). Deforestación de las Sierras Chicas de Córdoba (Argentina) en el período 1970-1997. Córdoba, Argentina: Academia Nacional de Ciencias. [ Links ]
Gavier-Pizarro, G. I., Radeloff, V. C., Stewart, S. I., Huebner, C. D., & Keuler, N. S. (2010). Rural housing is related to plant invasions in forests of southern Wisconsin, USA. Landscape Ecology, 25, 1505-1518. [ Links ]
Gavier-Pizarro, G. I., Kuemmerle, T., Hoyos, L. E., Stewart, S. I., Huebner, C. D., Keuler, N. S., & Rade- loff, V. C. (2012). Monitoring the invasion of an exotic tree Ligustrum lucidum from 1983 to 2006 with Landsat TM/ETM+ satellite data and Support Vector Machines in Córdoba, Argentina. Remote Sensing of Environment, 122, 134-145. [ Links ]
Giantomasi, A., Tecco, P. A., Funes, G., Gurvich, D. E., & Cabido, M. (2008). Canopy effects of the invasive shrub Pyracantha angustifolia on seed bank compo- sition, richness and density in a montane shrubland (Córdoba, Argentina). Austral Ecology, 33, 68-77. [ Links ]
Giorgis, M. A. (2011). Caracterización florística y estructural del Bosque Chaqueño Serrano (Córdoba) en relación a gradientes ambientales y de uso (Tesis de Doctorado). Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Córdo- ba, Argentina. [ Links ]
Giorgis, M. A., Cingolani, A. M., Chiarini, F., Chiapella, J., Barboza, G., Espinar, L. A., Morero, R., Gurvich, D. E., Tecco, P. A., Subils, R., & Cabido, M. (2011a). Composición florística del Bosque Chaqueño serrano de la provincia de Córdoba, Argentina. Kurtziana, 36, 9-43. [ Links ]
Giorgis, M. A., Tecco, P. A., Cingolani, A. M., Renison, D., Marcora P., & Paiaro, V. (2011b). Factors associated with woody alien species distribution in a newly invaded mountain system of central Argentina. Biological Invasions, 13, 1423-1434. [ Links ]
Giorgis, M. A., Cingolani, A. M., Gurvich, D. E., Tecco P. A., & Cabido, M. (2013). ¿Cuán estrecha es la relación entre la fisonomía y la composición florística a lo largo de gradientes ambientales en los “ecosiste- mas inciertos”? Boletín de la Sociedad Argentina de Botánica, suplemento 48. [ Links ]
Grman, E. & Suding, K. N. (2010). Within-Year Soil Legacies Contribute to Strong Priority Effects of Exotics on Native California Grassland Communities. Restoration Ecology, 18, 664-670. [ Links ]
Gurvich, D. E., Tecco, P. A., & Díaz, S. (2005). Plant invasions in undisturbed ecosystems: The triggering attribute approach. Journal of Vegetation Science, 16, 723-728. [ Links ]
Hejda, M., Pyšek, P., & Jarošík, V. (2009). Impact of invasive plants on the species richness, diversity and composition of invaded communities. Journal of Ecology, 97, 393-403. [ Links ]
Hoyos, L. E., Gavier-Pizarro, G. I., Kuemmerle, T., Bucher, E. H., Radeloff, V. C., & Tecco, P. A. (2010). Invasion of glossy privet (Ligustrum lucidum) and native forest loss in the Sierras Chicas of Córdoba, Argentina. Biological invasions, 12, 3261-3275. [ Links ]
Hulme, P. E. (2006). Beyond control: Wider implications for the management of biological invasions. Journal of Applied Ecology, 43, 835-847. [ Links ]
Jobbágy, E. G., Acosta, A. M., & Nosetto, M. D. (2013). Rendimiento hídrico en cuencas primarias bajo pastizales y plantaciones de pino de las sierras de Córdoba (Argentina). Ecología Austral, 23, 87-96. [ Links ]
Kakembo, V., Rowntree, K., & Palmer, A. R. (2007). Topographic controls on the invasion of Pteronia incana (Blue bush) onto hillslopes in Ngqushwa (formerly Peddie) district, Eastern Cape, South Africa. Catena, 70, 185-199. [ Links ]
Kolb, A., Alpert, P., Enters, D., & Holzapfel, C. (2002). Patterns of Invasion within a Grassland Community. Journal of Ecology, 90, 871-881. [ Links ]
Levine, J. M., Vila, M., Antonio, C. M., Dukes, J. S., Grigulis, K., & Lavorel, S. (2003). Mechanisms underlying the impacts of exotic plant invasions. Proceedings of the Royal Society of London, 270, 775-781. [ Links ]
Lichstein, J. W., Grau, H. R., & Aragón, R. (2004). Recruitment limitation in secondary forests domina- ted by an exotic tree. Journal of Vegetation Science, 15, 721-728. [ Links ]
MacDougall, A. S., Boucher, J., Turkington R., & Bradfield, G. E. (2006). Patterns of plant invasion along an environmental stress gradient. Journal of Vegetation Science, 17, 47-56. [ Links ]
Mack, R. N., Simberloff, D., Mark Lonsdale, W., Evans, H., Clout, M., & Bazzaz, F. A. (2000). Biotic inva- sions: causes, epidemiology, global consequences, and control. Ecological Applications, 10, 689-710. [ Links ]
Mack, M. C. & D’Antonio, C. M. (2003). Exotic grasses alter controls over soil nitrogen dynamics in Hawaiian woodland. Ecological Applications, 13, 154-166. [ Links ]
Marchante, E., Kjøller, A., Struwe, S., & Freitas, H. (2009). Soil recovery after removal of the N2-fixing invasive Acacia longifolia: Consequences for ecosystem restoration. Biological Invasions, 11, 813-823. [ Links ]
McKinney, M. L. & Lockwood, J. L. (1999). Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends in Ecology & Evolution, 14, 450-453. [ Links ]
Nelson, D. W. & Sommers, L. E. (1996). Total Carbon, Organic carbon, and Organic Matter. In D. L. Sparks, A. L. Page, P. A. Helmke, R. H. Loeppert, P. N. Soltanpour, M. A. Tabatabai, C. T. Johnston, & M. E. Sumner (Eds.), Methods of Soil Analysis. Part 3 – Chemical Methods (pp. 961-1010). Madison, Wisconsin: ASA, SSSA, CSSA. [ Links ]
Nichols, G. E. (1923). A Working Basis for the Ecological Classification of Plant Communities. Ecology, 4, 154-179. [ Links ]
Organización de las Naciones Unidas para la Educación, la Ciencia y la Cultura [UNESCO]. (1973). Clasificación internacional y cartografía de la vegetación (pp. 1-112). [ Links ]
Pauchard, A., Kueffer, C., Dietz, H., Daehler, C. C., Alexander, J., Edwards, P. J., Arévalo, J. R., Cavieres, L. A., Guisan, A., Haider, S., Jakobs, G., McDougall, K., Millar, C. I., Naylor, B. J., Parks, C. G., Rew, L. J., & Seipel, T. (2009). Ain’t no mountain high enough: plant invasions reaching new elevations. Frontier Ecology Environment, 7, 479-486. [ Links ]
R Development Core Team. (2012). R: A Language and environment for statistical computing. Vienna, Austria: R. Foundation for Statistical Computing. Recuperado de http://www.R-roject.org, Vienna, Austria. [ Links ]
Schofield, C. J. & Bucher, E. H. (1986). Industrial Con- tributions to Desertification in South America. Tree, 1, 78-80. [ Links ]
Simberloff, D. (2009). The Role of Propagule Pressure in Biological Invasions. Annual Review of Ecology Evolution and Systematics, 40, 81-102. [ Links ]
Strayer, D. L., Eviner, V. T., Jeschke, J. M., & Pace, M. L. (2006). Understanding the long-term effects of species invasions. Trends in Ecology & Evolution, 21, 645-651. [ Links ]
Tecco, P. A., Gurvich, D. E., Díaz, S., Pérez-Harguindeguy, N., & Cabido, M. (2006). Positive interaction between invasive plants: The influence of Pyracantha angustifolia on the recruitment of native and exotic woody species. Austral Ecology, 31, 293-300. [ Links ]
Tecco, P. A., Diaz, S., Gurvich, D. E., Perez-Harguindeguy, N., Cabido, M., & Bertone, G. A. (2007). Facilitation and interference underlying the association between the woody invaders Pyracantha angustifolia and Ligustrum lucidum. Applied Vegetation Science, 10, 211-218. [ Links ]
Tecco, P. A., Urcelay, C., Díaz, S., Cabido, M., & Pérez- Harguindeguy, N. (2013). Contrasting functional trait syndromes underlay woody alien success in the same ecosystem. Austral Ecology, 38, 443-451. [ Links ]
Thomas, G. W. (1996). Soil pH and soil acidity. In D. L. Sparks, A. L. Page, P. A. Helmke, R. H. Loeppert, P. N. Soltanpour, M. A. Tabatabai, C. T. Johnston, & M. E. Sumner (Eds.), Methods of Soil analysis. Part 3 – Chemical Methods (Capítulo 16). Madison, Wisconsin: ASA, SSSA, CSSA. [ Links ]
Throop, H. L., Archer, S. R., Monger, H. C., & Waltman, S. (2012). When bulk density methods matter: implications for estimating soil organic carbon pools in rocky soils. Journal of Arid Environments, 77, 66-71. [ Links ]
Whittaker, R. H. (1970). Communities and ecosystems. New York: McMillan. [ Links ]
Wolfe, B. E. & Klironomos, J. N. (2005). Breaking new ground: soil communities and exotic plant invasion. Bioscience, 55, 477-487. [ Links ]
Woodward, F. I. (1987). Climate and plant distribution. Cambridge: Cambridge University Press. [ Links ]
Zak, M. R., Cabido, M., & Hodgson, J. G. (2004). Do subtropical seasonal forest in the Gran Chaco, Argentina, have a future? Biological Conservation, 120, 589-598. [ Links ]
Zuloaga, F. O., Morrone, O., & Belgrano, M. J. (2008). Catalogue of the vascular plants of the southern cone (Argentina, southern Brazil, Chile, Paraguay and Uruguay). Missouri: Missouri Botanical Garden Press. [ Links ]
1. Instituto Multidisciplinario de Biología Vegetal (CONICET – UNC), Av. Vélez Sársfield 1611, CC 495, X5000HVA Córdoba, Argentina; sebazeba@hotmail.com
2. Cátedra de Biogeografía, Departamento de Diversidad Biológica y Ecología, Facultad de Ciencias Exactas Físicas y Naturales, Universidad Nacional de Córdoba, Av. Vélez Sársfield 299, X5000JJC, Córdoba, Argentina; ptecco@efn.uncor.edu, mcabido@imbiv.unc.edu.ar, dgurvich@com.uncor.edu
Recibido 21-I-2014. Corregido 18-V-2014. Aceptado 19-VI-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}