










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.4 San José Oct./Dec. 2013
Population biology and diet of Pomadasys corvinaeformis (Perciformes: Pomadasyidae) in Caraguatatuba Bay, Southeastern Brazil
Biología de poblaciones y dieta de Pomadasys corvinaeformis (Perciformes: Pomadasyidae) en Bahía Caraguatatuba, sureste de Brasil
Biología de poblaciones y dieta de Pomadasys corvinaeformis (Perciformes: Pomadasyidae) en Bahía Caraguatatuba, sureste de Brasil
Márcia Regina Denadai1*, Flávia Borges Santos2*, Eduardo Bessa3, Wellington Silva Fernandez4*, Luana Lorca5* & Alexander Turra4
*Dirección para correspondencia:
Abstract
Pomadasys corvinaeformis inhabits sandy and rocky bottoms in coastal waters, and is common in trawl samples taken from beaches. The species is very abundant on the Brazilian coast, and is of high economic and ecological importance. This study examined the spatio-temporal distribution, population biology and diet of P. corvinaeformis in Southeastern Brazil. Samples were taken by trawling monthly from August 2003 to October 2004, in two previously selected areas. The Northern area is more exposed to wave activity and is influenced by a river, functioning as a small estuary. In contrast, the Southern area is relatively sheltered from wave energy and influenced to a lesser degree by smaller rivers. The length of the specimens was measured, and the sex and gonadal stage were macroscopically identified. The abundance of this species was compared between areas and among months. The diet was analyzed seasonally by the frequency of occurrence, the percent volume, and the index of alimentary importance. P. corvinaeformis occurred in unequal proportions in the two study areas (86% in the Northern area and 14% in the Southern area) and was found most abundant in May 2004, followed by June 2004. The proportion of mature and in-maturation individuals increased gradually from autumn to summer. Nine major groups of food items were recorded in the diet of P. corvinaeformis, and crustaceans comprised five of the categories: unidentified crustacean fragments, zoea larvae, amphipods, copepods and shrimps. In both, fish stomach and intestine, crustacean fragments were the most frequent item. The second most frequent items were shrimp in the stomach, and amphipods in the intestine (mainly represented by their tubes). These results demonstrate that P. corvinaeformis can be considered a carnivore, with a preference on benthic organisms.
Key words: Corcoroca, feeding, Pomadasys corvinaeformis, roughneck grunt, spatio-temporal distribution, São Paulo.
Resumen
Pomadasys corvinaeformis, el pargo blanco, habita fondos rocosos y arenosos de zonas costeras y es común en arrastres de playas. La especie es muy abundante en la costa brasileña, teniendo una alta importancia ecológica y económica. Este estudio examina la distribución espacio-temporal, biología poblacional y la dieta de P. corvinaeformis en el sudeste de Brasil, a través de muestras mensuales tomadas desde agosto 2003 hasta octubre 2004, mediante arrastres en dos áreas previamente seleccionadas. El área norte es más expuesta a la acción de las olas y está bajo la influencia de un río, funcionando como un pequeño estuario. En contraste, el área sur es relativamente protegida de las olas e influenciada en menor grado por pequeños riachuelos. El largo, sexo y estadio gonadal de los organismos fue determinado y la abundancia de la especie comparada entre áreas y entre meses. La dieta a su vez, se estudió estacionalmente por medio de la frecuencia de ocurrencia, volumen porcentual y el índice de importancia alimentaria. Los resultados mostraron que P. corvinaeformis ocurrió de forma diferencial entre las dos áreas (86% en la norte y 14% en la sur) y además fue más abundante en mayo 2004, seguido de junio 2004. La proporción de organismos maduros/inmaduros creció gradualmente desde el otoño hasta el verano. Relacionado con la dieta, fueron registrados nueve grupos mayores de ítems alimentarios, estando los crustáceos presentes en cinco de esas categorías: fragmentos de crustáceos no identificados, larvas zoeas, anfípodos, copépodos y camarones. Tanto en el estómago como en el intestino de los peces, los fragmentos de crustáceos fueron el ítem más abundante, mientras que el segundo ítem en importancia fueron camarones en el estómago y anfípodos en el intestino (mayormente representados por su tubo). Los resultados indican que P. corvinaeformis puede ser considerado carnívoro, con clara preferencia por organismos bentónicos.
Palabras clave: alimentación, distribución espacio-temporal, pargo blanco, Pomadasys corvinaeformis, roughneck grunt, São Paulo.
The Roughneck grunt Pomadasys corvinaeformis occurs in the Western Atlantic from Mexico to Southern Brazil (Smith, 1997). It inhabits sandy and rocky bottoms of coastal waters (Courtenay & Sahlman, 1978), is common in trawls taken from beaches (Menezes & Figueiredo, 1980) and is found to a depth of 50m (Cervigón et al. 1992). The species is also found in low-salinity waters (Cervigón, 1993). These grunts may reach a maximum size of 25cm, since Menezes & Figueiredo (1980) recorded an individual with 24.8cm. They feed on crustaceans and small fish and have commercial value.
The species is common on the Brazilian coast (Menezes & Figueiredo, 1980), and is one of the most numerous species in the Southern coast ichthyofauna of Paraná in Southern Brazil (Chaves, 1998). Pomadasys corvinaeformis showed wide seasonal variations in abundance and age distribution in Ceará, Northeastern Brazil (Costa, Santos, Espínola & Monteiro-Neto, 1995). In Caraguatatuba Bay, which is strongly influenced by the local rivers and therefore has estuarine characteristics, P. corvinaeformis was numerous only in the autumn.
Although P. corvinaeformis is an abundant species on the Brazilian coast, the relatively few studies on its biology have examined the biometry (Costa et al. 1995), population structure (Chaves, 1998) and parasites (Cavalcanti, Chellappa, Pavanelli & Takemoto, 2006); as well as its cogeneric species P. commersonnii (Childs et al. 2008), P. incisus (Pajuelo, Lorenzo, Gregoire & Domínguez-Seoane, 2003) and P. kakaan (Al-Husaini et al. 2002).
P. corvinaeformis is relatively uncommon in Caraguatatuba Bay, although it has high economic and ecological importance in other Brazilian regions. The ichthyofauna of Caraguatatuba Bay has been little studied. This study is part of a wide-ranging pioneer research project on the megaepifauna of this bay, and the project is expected to provide highly important data to contribute to the development of future management plans for the region. Caraguatatuba Bay and its neighborhood have been impacted by socio-economic development including the construction of natural-gas pipelines and a gas treatment facility, plus the expansion of the port and oil terminal of São Sebastião. Since December 2004 (after the collections of this study ended), the state decree No. 49 215, which treats the Economic Ecological Zone (ZEE) of São Paulo’s Northern Shore, prohibited trawling in bay areas. Therefore, studies focusing on the ecology of species such as P. corvinaeformis provide important baseline data for evaluation of the possible impacts to the region.
Material and methods
Samples were taken monthly from August 2003 through October 2004 in two areas of Caraguatatuba Bay (23o37’S - 45o26’W and 23o44’S - 45o24’W; Fig. 1). Three sampling stations were selected in each area, South and North, among 200 possibilities, i.e., the beach length of 2 000m divided into 10m intervals. The position of each station was stored in a GPS at MLW (mean low water) and then the starting point for the trawls, 800m perpendicularly outward from the beach and beginning at MLW, was determined. At each station a 800m trawl was performed, from 800 to 1 600m perpendicular to the beach. This interval is equivalent to a water depth of 1 to 4m. The trawls were performed with a fishing boat, class G2M, 11m long with a 22HP engine. The trawling speed was about 1knot. The trawls were performed using two otter trawls with 2.0cm mesh, mouth aperture 1.6m in height and 6.0m in length, and bag depth 3.5m.
The fish were then removed from the net and immediately preserved in a 10% formalin solution in order to halt the enzyme action, and to preserve the digestive tube contents (Uieda & Castro, 1999). The samples were identified and stored in plastic containers. All specimens were identified to species in the laboratory and then transferred to 70% ethanol.
All individuals of P. corvinaeformis collected were measured for standard length (SL), which is the distance between the anterior end of the head and the base of the caudal fin (beginning of caudal fin rays; Figueiredo & Menezes, 1978).
A total of 160 individuals of P. corvinaeformis were sorted from the 346 obtained in the samples for the diet analysis, using a random-digits table. An abdominal ventro-sagittal incision was made from the anal aperture to the pelvic fin insertions. The digestive tubes and the gonads were removed. The gonads were analyzed, sexed and classified according to the maturation stages defined by Vazzoler (1996).
The digestive-tube length (distance between the beginning of the esophagus and the end of the rectum; DTL) was measured in order to establish the DTL/SL ratio (digestive tube length/standard length) and evaluate a possible relationship to the diet of P. corvinaeformis (Uieda & Castro, 1999). The digestive tubes were then preserved in 70% ethanol until analysis of diet contents.
The digestive tubes were divided into two portions, the first constituted by the pharynx, esophagus and stomach, and the second by the intestine and rectum, due to the different degrees of digestion of the contents in each portion. Finally, the contents of each digestive tube were identified. The volume of each item was measured according to methodologies from Bemvenute (1990) and Petti (1997).
The mean number of individuals (±SE) of P. corvinaeformis was calculated for the months and areas (South and North). The numbers in the two areas was also summed together. A two-way Anova was performed to test the differences in the spatial (South and North areas) and temporal (months) distributions. A histogram illustrating the relative frequency distribution of the size classes (standard length) of this species was constructed for the total population. A chi-square test was performed to determine if the sex ratio was equivalent between males and females with α=0.05.
The diet of P. corvinaeformis was analyzed seasonally by the frequency of occurrence (F%), percent volume (V%) and alimentary importance index (AIi). The alimentary importance index was calculated by a modification of the method used by Kawakami & Vazzoler (1980), based on the frequency of occurrence (Fi%) and on the percent volume (Vi%) of each item.
Results
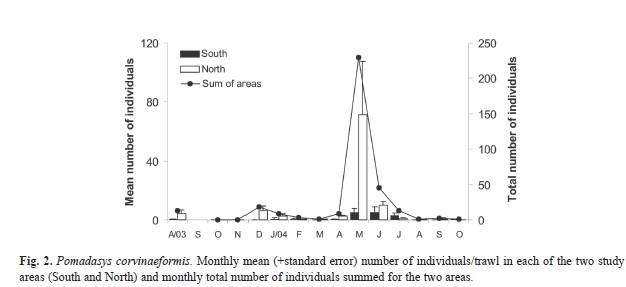
Pomadasys corvinaeformis was relatively uncommon in Caraguatatuba Bay during the study period; 346 individuals were obtained in the six monthly trawls made during the 15 months of this study. Of these, 86% were in the North area and 14% in the South area (Anova: F=4.993, df=1, p=0.029). The highest numbers were caught in May and June 2004, when 80% of all individuals were caught (Anova: F=4.099, df=13, p<0.001) (Fig. 2).
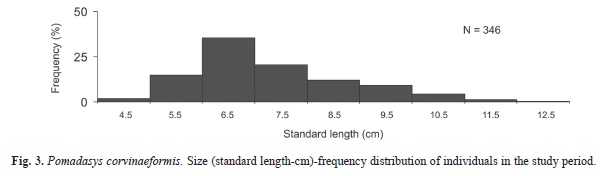
The distribution of the individuals among months seemed to be independent of the capture areas (South and North) (Anova: F=3.086, df=13, p=0.002). The lengths of the fish ranged from 4.5 to 12.5cm (Fig. 3), with a mean value of 7.34±1.44.
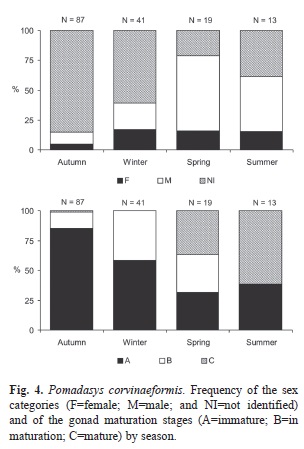
Most of the fish (68%) were immature and could not be sexed. Therefore the sex ratio was provided for only a few individuals by season (16, 13, 15 and 8, respectively for the autumn, winter, spring and summer), with a predominance of males (χ2=200, df=1, p<0.001) (Fig. 4). The proportion of mature and in-maturation fish gradually increased from autumn to summer (Fig. 4). In the autumn, 85% of the 87 fish analyzed were immature; in the winter 58.6% of the 41 individuals were immature and 41.4% in maturation; in the spring there were six immature individuals, six in maturation and seven already mature; and in the summer, five individuals were immature and eight were mature, with no individual in maturation recorded.
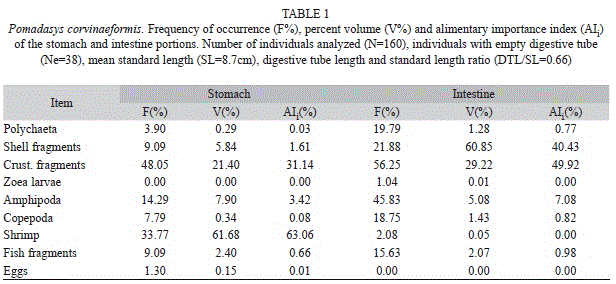
Nine food items were recorded, as major groups, in the diet (Table 1). Crustaceans comprised five of the categories: unidentified crustacean fragments, zoea larvae, amphipods (entire or tubes), copepods and shrimp. The polychaetes were represented by their fragments and tubes, mollusks by their shells, fish by their fragments and scales; and the producers of unidentified eggs. Of the 160 analyzed individuals, 38 had empty digestive tubes. The DTL/SL was calculated as 0.66.
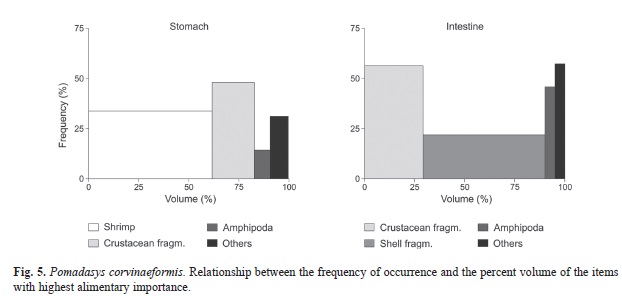
Comparing the frequency of occurrence of the items between stomach and intestine (Table 1), some particularities are observed. In both the portions, crustacean fragments were the most frequent item. The second item in frequency in the stomach was shrimp, and in the intestine, amphipods (mainly represented by their tubes). Polychaetes, shell fragments, copepods and fish fragments were more frequent in the intestine. Zoea larvae occurred exclusively in the intestine, and eggs exclusively in the stomach. With respect to the volume of the main items ingested (Table 1), shrimp, an easily digestible item, were important in the stomach contents, while in the intestine the presence of shell fragments, which persist after the mollusks soft parts are digested, indicate that this item is also important in the diet of P. corvinaeformis. The alimentary indexes, shown in figure 5, by the relationship between frequency of occurrence and percent volume, show that the most voluminous items, both in the stomach (shrimp) and in the intestine (shell fragments), were not the most frequent, although shrimp was the most important item recorded in the stomach contents (AIi=63.06), while crustacean fragments were the most important item in the intestinal contents (AIi=49.92). Crustacean fragments were the most frequent item in both portions of the digestive tube.
Unidentified organic matter was present in 28.7% of the stomachs with contents, totaling 7.84% of the volume found in this portion of the gut and in 48.7% of the intestines with contents, totaling 16.41% by volume of the total gut contents.
Discussion
Pomadasys corvinaeformis is a common species on the Brazilian coast (Menezes & Figueiredo, 1980). It is one of the five most abundant species taken in traditional fisheries in Fortaleza, Ceará (Costa et al. 1995) and also one of the most abundant in the ichthyofauna of Guaratuba Bay, Paraná (Chaves, 1998). Pomadasys corvinaeformis in Caraguatatuba Bay occurred mainly at the end of autumn (May to June 2004). Costa et al. (1995) correlated the differences in the capture of this species to the periods of heavier and lighter rainfall. These authors observed a higher density of this fish in the bimester following the higher mean rainfall, and associated this with the intake of nutrients from continental drainages, which increases the availability of food to the juveniles and larvae. The coast of São Paulo state, however, does not have well defined rainy and dry seasons, although rainfall is heavier in the summer. Thus, as previously recorded by Chaves (1998) for a population of P. corvinaeformis on the coast of Paraná, the abundance pattern observed in Caraguatatuba Bay is the inverse of that observed in Fortaleza. As observed by Chaves (1998), the occurrence of the population in Caraguatatuba Bay, as well as the one in Guaratuba Bay, in bays rather than off an open-sea beach, as in Fortaleza, may be a cue that rainfall and salinity are not the only factors that affect abundance patterns for the species.
The data for the gonad maturation stages in Caraguatatuba Bay, although only a few individuals were analyzed, showed an increase in the proportion of sexually mature individuals in the summer, the period when this species probably spawns. Chaves (1998) determined that Guaratuba Bay is not the spawning site for P. corvinaeformis, and suggested that the low numbers of individuals in this bay during the summer, with their return at the end of this season, coincides with the spawning period for the species in the open sea. The same was observed by Costa, Santos, Espínola & Monteiro-Neto (1995) for this species in Fortaleza, that is, a reduction in the number of individuals during the rainy period (November to February; summer) with displacement of the peak of frequencies for the larger size classes. This reduction was then followed in March-April (end of summer and beginning of autumn) by the entrance of juveniles into the population, i.e., juvenile recruitment. Probably the same argument explains the lower abundance of this fish in the warmer months of the year in Caraguatatuba Bay, as well as the dominance of small individuals, because the larger individuals may reach 25cm (Menezes & Figueiredo, 1980). Therefore, Caraguatatuba Bay is likely used for shelter and growth by the juveniles of P. corvinaeformis, which probably only reach capture size (5.5 to 7.5cm) in the autumn, as also observed by Costa et al. (1995).
The diet of P. corvinaeformis in Caraguatatuba Bay was based mainly on crustaceans (represented by unidentified fragments, shrimp and amphipods) and mollusks (represented by shell fragments). The separation of the stomach and intestine portions of the gut and the use of the data from the former portion made it possible to identify marked differences in the species’ diet. In the stomach contents, shrimp were highly important, consumed by 33.8% of the individuals and totaling 61.7% of the volume of ingested food. For the intestine, mollusk shells were important, being consumed by 21.9% of the individuals and totaling 60.9% of the ingested food. Therefore, if only the stomach contents were analyzed, little importance would be given to mollusks in the diet of this fish. The shells are preserved in the intestinal contents after the soft parts are completely digested. The crustacean fragments and the amphipod and polychaete tubes were other items that remained in the intestinal contents, although they were also recorded in the stomach.
Similarly to our data obtained in Caraguatatuba Bay, Costa et al. (1995) found, in order of importance, the following items in the diet of P. corvinaeformis in Fortaleza: crustaceans, polychaetes, fish and algae. No algae were found in the individuals examined in Caraguatatuba Bay. Likewise, mollusks were not obtained in the study in Fortaleza, demonstrating some differences in the diet of fish from the two regions. Among the crustaceans, mysids, copepods and amphipods were the three most important groups in the diet of this fish at Fortaleza (Costa et al. 1995), evidencing a tendency to predate on vagile microcrustaceans. Costa et al. (1995) concluded that P. corvinaeformis possesses a varied feeding spectrum, and feeds throughout the water column. The DTL/SL ratio calculated for the fish in Caraguatatuba Bay (0.66) suggests a carnivorous habit. Konchina (1977) stated that most members of the family Pomadasyidae are zoophagous, and Menezes & Figueiredo (1980) referred to P. corvinaeformis as feeding on crustaceans and small fish.
Costa et al. (1995) also noted the presence of large amounts of organic matter and unidentified crustacean remains in the diet of P. corvinaeformis, and related this to the advanced degree of digestion of the items. The presence of sand in the stomach contents, according to these authors, may be due to the voluntary ingestion for use as abrasives in digestion, or involuntary, occurring as the net is pulled through the surf zone to the beach. In Caraguatatuba Bay, unidentified organic matter was relatively frequent (28.7% of the stomachs and 48.7% of the intestines) in the individuals analyzed, and was more voluminous in the intestine (16.4%) than in the stomach (7.84%). This information demonstrates that the observation by Costa et al. (1995) of advanced digestion should be accepted, because the volume of unidentified organic matter is larger in the intestine. However, in view of the benthophagous habit of the species, involuntary ingestion of deposited organic matter is also possible. No significant amount of sand was found in the digestive tract of the fish analyzed in Caraguatatuba Bay, which reinforces the hypothesis suggested by Costa et al. (1995) of involuntary ingestion of sand by the fish during beach trawling, because the collections in Caraguatatuba Bay involved a different fishing method.
The results presented above demonstrates that Pomadasys corvinaeformis is rare in Caraguatatuba Bay, and is occasionally found in high abundance, as in May-June 2004. Pomadasys corvinaeformis can be considered a carnivore, showing a preference to consume benthic organisms such as shrimp, amphipods and mollusks.
Acknowledgments
We thank the State of São Paulo Research Foundation (FAPESP), which financed this study; and the scholarship in the program Jovem Pesquisador em Centro Emergente awarded to Márcia Regina Denadai (Proc. Fapesp No. 06/57575-1). We thank also Janet W. Reid, who revised the English text.
References
Al-Husaini, M., Al-Baz, A., Al-Ayoub, S., Safar, S., Al-Wazan Z., & Al-Jazzaf, S. (2002). Age, growth, mortality, and yield-per-recruit for nagroor, Pomadasys kakaan, in Kuwait’s waters. Fisheries Research, 59, 101-115. [ Links ]
Bemvenute, M. A. (1990). Hábitos alimentares de peixes-rei (Atherinidae) na região estuarina da Lagoa dos Patos. Rev. Atlântica, 12, 79-102. [ Links ]
Cavalcanti, E. T. S., Chellappa, S., Pavanelli, G. C., & Takemoto, R. M. (2006). Presença de Lernanthropus rathbuni (Copepoda: Lernanthropidae) no coró, Pomadasys corvinaeformis (Osteichthyes: Haemulidae) em águas costeiras do Rio Grande do Norte. Arquivos de Ciências do Mar, 39, 134-137. [ Links ]
Cervigón, F. (Ed.). (1993). Los peces marinos de Venezuela. Vol. 2. Caracas: Fundación Científica Los Roques. [ Links ]
Cervigón, F., Cipriani, R., Fischer, W., Garibaldi, L., Hendrickx, M., Lemus, A. J., Márquez, R., Poutiers, J. M., Robaina G., & Rodriguez, B. (Eds.). (1992). Fichas FAO de identificación de especies para los fines de la pesca. Guía de campo de las especies comerciales marinas y de aguas salobres de la costa septentrional de Sur América. Rome: FAO. [ Links ]
Chaves, P. T. C. (1998). Estrutura populacional de Pomadasys corvinaeformis (Steindachner) (Teleostei, Haemulldae) na Baía de Guaratuba, Paraná, Brasil. Revista Brasileira de Zoologia, 15, 203-209. [ Links ]
Childs, A. R., Booth, A. J., Cowley, P. D., Potts, W. M., Næsje, T. F., Thorstad, E. B., & Økland, F. (2008). Home range of an estuarine-dependent fish species Pomadasys commersonnii in a South African estuary. Fisheries Management and Ecology, 15, 441-448. [ Links ]
Costa, P. S. R., Santos, M. A. M., Espínola M. F. A., & Monteiro-Neto, C. (1995). Biologia e biometria do coró, Pomadasys corvinaeformis (Steindachner) (Teleostei: Pomadasyidae), em Fortaleza, Ceará, Brasil. Arquivos de Ciências do Mar, 29, 20-27. [ Links ]
Courtenay, W. R., & Sahlman, H. F. (1978). Pomadasyidae. In W. Fischer (Ed.). FAO species identification sheets for fishery purposes. Western Central Atlantic (Fishing Area 31) (4th ed.). Rome: FAO. [ Links ]
Figueiredo, J. L., & Menezes, N. A. (Eds.). (1978). Manual de peixes marinhos do sudeste do Brasil. V. Teleostei (1). São Paulo: Museu de Zoologia da Universidade de São Paulo. [ Links ]
Kawakami, E., & Vazzoler, G. (1980). Método gráfico e estimativa de índice alimentar aplicado no estudo de alimentação de peixes. Boletim do Instituto Oceanográfico, 29, 205-207. [ Links ]
Konchina, Y. V. (1977). Some data on the biology of grunts (Family Pomadasyidae). Journal of Ichthyology, 17, 548-558. [ Links ]
Menezes, N. A., & Figueiredo, J. L. (Eds.). (1980). Manual de peixes marinhos do sudeste do Brasil. V. Teleostei (4). São Paulo: Museu de Zoologia da Universidade de São Paulo. [ Links ]
Pajuelo, J. G., Lorenzo, J. M., Gregoire, M., & Domínguez-Seoane, R. (2003). Life history of Pomadasys incisus (Osteichthyes: Haemulidae) in the Canarian Archipelago. Scientia Marina, 67, 241-248. [ Links ]
Petti, M. A. V. (1997). Papel dos crustáceos braquiúros na rede trófica da plataforma interna de Ubatuba, SP. Nerítica, 11, 123-137. [ Links ]
Smith, C. L. (Ed.). (1997). Field guide to tropical marine fishes of the Caribbean, the Gulf of Mexico, Florida, the Bahamas, and Bermuda. National Audubon Society. New York: Chanticleer Press Edition. [ Links ]
Uieda, V. S., & Castro, R. M. C. (1999). Coleta e fixação de peixes de riachos, In E. P. Caramaschi, R. Mazzoni, & P. R. Peres-Neto (Eds.). Ecologia de Peixes de Riachos. (pp. 01-22). Rio de Janeiro: Série Ecologia Brasiliensis. [ Links ]
Vazzoler, A. E. A. M. (Ed.). (1996). Biologia da Reprodução de peixes Teleósteos: Teoria e Prática. Maringá: Eduem. [ Links ]
*Correspondencia a:
Márcia Regina Denadai: Centro Universitário Módulo. 11660-903, Caraguatatuba, SP, Brasil; marciard@gmail.com
Flávia Borges SantosUniversidade Estadual do Sudoeste da Bahia, Departamento de Ciências Naturais. 45083-900, Vitória da Conquista, BA, Brasil; flaborgs@gmail.com
Eduardo Bessa: Instituto de Ciência Naturais e da Terra da Universidade do Estado de Mato Grosso. 78300-000, Tangara da Serra, MT, Brasil; edu_bessa@yahoo.com
Wellington Silva Fernandez: Instituto Oceanográfico da Universidade de São Paulo, Departamento de Oceanografia Biológica. 05508-120, São Paulo, SP, Brasil; fernandez@usp.br
Luana Lorca: Centro Universitário da Fundação de Ensino Octávio Bastos. Rua General Osório, 433, 13870-431, São João da Boa Vista, SP, Brasil; luana_lorca@gmail.com
Alexander Turra: Instituto Oceanográfico da Universidade de São Paulo, Departamento de Oceanografia Biológica. 05508-120, São Paulo, SP, Brasil; turra@usp.br
1. Centro Universitário Módulo. 11660-903, Caraguatatuba, SP, Brasil; marciard@gmail.com
2. Universidade Estadual do Sudoeste da Bahia, Departamento de Ciências Naturais. 45083-900, Vitória da Conquista, BA, Brasil; flaborgs@gmail.com
3. Instituto de Ciência Naturais e da Terra da Universidade do Estado de Mato Grosso. 78300-000, Tangara da Serra, MT, Brasil; edu_bessa@yahoo.com
4. Instituto Oceanográfico da Universidade de São Paulo, Departamento de Oceanografia Biológica. 05508-120, São Paulo, SP, Brasil; turra@usp.br, fernandez@usp.br
5. Centro Universitário da Fundação de Ensino Octávio Bastos. Rua General Osório, 433, 13870-431, São João da Boa Vista, SP, Brasil; luana_lorca@gmail.com
Received 18-IX-2012. Corrected 10-II-2013. Accepted 12-III-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}