










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.4 San José Oct./Dec. 2013
Morphology and ultrastructure of the freshwater rotifer Brachionus bidentatus (Monogononta: Brachionidae) using scanning and transmission electron microscopy
Morfología y ultraestructura del rotífero de agua dulce Brachionus bidentatus (Monogononta: Brachionidae), utilizando microscopía electrónica de barrido y transmisión
Morfología y ultraestructura del rotífero de agua dulce Brachionus bidentatus (Monogononta: Brachionidae), utilizando microscopía electrónica de barrido y transmisión
Gerardo Guerrero-Jiménez1*, Guadalupe Zavala-Padilla2*, Marcelo Silva-Briano3* & Roberto Rico-Martínez1
*Dirección para correspondencia:
Abstract
The study of sexual reproductive behavior supported by ultrastructural evidence is important in rotifers to describe differences among potential cryptic species. In this research, the morphology of the rotifer Brachionus bidentatus is described at the ultrastructural level, using electronic microscopy, together with a brief description and discussion of its sexual reproductive behavior. The characteristics of the (a) male,(b) the female, (c) the sexual egg or cyst, (d) the partenogenic egg, (e) the no-fecundated sexual egg (male egg), and (f) the trophi, were described. Another part of this research is dedicated to the ultrastructure of the sex cells of the male rotifer B. bidentatus. Samples were obtained from La Punta pond in Cosio, Aguascalientes, Mexico (22°08’ N - 102°24’ W), and a culture was maintained in the laboratory. Fifty organisms, from different stages of the rotifer Brachionus bidentatus, were fixed in Formol at 4% and then prepared; besides, for the trophi, 25 female rotifer Brachionus bidentatus were prepared for observation in a JEOL 5900 LV scanning electronic microscope. In addition, for the observation of male sex cells, 500 males of Brachionus bidentatus were isolated, fixed and observed in a JEOL 1010 transmission microscope. Females of B. bidentatus in laboratory cultures had a lifespan of five days (mean±one SD=4.69±0.48; N=13), and produced 4.5+3.67 (N=6) parthenogenetic eggs during such lifespan. In the case of non-fertilized sexual eggs, they produced up to 18 eggs (mean±one SD=13±4.93; N=7). Sexual females produced a single cyst on average (mean±one SD=1±0; N=20). For the sexual cycle, the time of copulation between male and female ranged from 10 to 40 seconds (mean±one SD=17.33±10.55, N=7). The spermatozoa are composed of a celular body and a flagellum, the size of the body is of 300nm while the flagellum measures 1 700nm. The rods have a double membrane. Their mean length is almost 2.45µm±0.74, N=6; and their mean wide is 0.773µm±0.241, N=11. The evidence on the specific ultrastructural characteristics of the rotifer B. bidentatus is notorious, even more in the male and in the cyst cell. Regarding the ultrastructure of the spermatozoa and the rods, compared to other species they only differ in size, despite their structural resemblance. Our study of the ultraestructure of this species adds useful information that along with molecular data will help clarify the taxonomy of brachionid rotifers. Rev. Biol. Trop. 61 (4): 1737-1745. Epub 2013 December 01.
Key words: Rotifer, Brachionus bidentatus, ultrastructure, rods, spermatozoon.
Resumen
El estudio del comportamiento reproductivo sexual apoyado en evidencias ultraestructurales en rotíferos, es importante para describir diferencias entre especies potencialmente crípticas. En este trabajo se describe a nivel ultraestructural la morfología del rotífero Brachionus bidentatus, usando microscopía electrónica, junto con una breve descripción y discusión de su comportamiento sexual reproductivo. Se presentan las características del: (a) macho, (b) hembra, (c) huevo partenogenético, (d) huevo sexual no fecundado, (e) trofos. También se muestra un apartado sobre la ultra estructura de las células sexuales de rotíferos macho B. bidentatus. Para el estudio se utilizó una cepa proveniente del bordo La Punta (22°08’ N - 102°24’ W), ubicado en Cosió, estado de Aguascalientes, México y se cultivaron en el laboratorio. Para el procesamiento de las muestras se tomaron 50 organismos fijados en Formol al 4%, de los diferentes estadios del rotífero Brachionus bidentatus, mientras que para el trofos se tomaron 25 organismos hembra del rotífero Brachionis bidentatus y se prepararon para observarse en un microscopio electrónico de barrido JEOL 5900 LV, mientras que para las observaciones de las células sexuales del macho se aislaron 500 organismos machos del rotífero Brachionus bidentatus, se fijaron e incluyeron en resina epóxica (EPON) para su observación en un microscopio electrónico de transmisión JEOL 1010 operado a 80kv. Los análisis obtenidos de hembras de B. bidentatus en cultivos de laboratorio demuestran un ciclo de vida de cinco días (4.69±0.48; N=13), y una producción de 4.5±3.67 (N=6) huevos partenogenéticos. En el caso de los huevos no fertilizados, la hembra produce más de 18 huevos (13±4.93; N=7). Para los huevos sexuales solo se produce uno solo por hembra (1±0; N=20). En el ciclo sexual, los tiempos de copula entre el macho y la hembra están en el rango de 10 a 40 segundos (17.33±10.55, N=7). Los espermatozoides se componen de un cuerpo celular y un flagelo, el tamaño del cuerpo celular es de 300nm mientras que el flagelo mide 1 700nm. Los bastones presentan una doble membrana y su tamaño a lo largo va de 2.45µm±0.74; N=6 mientras que el ancho es de 0.773µm±0.241; N=11. La evidencia sobre las características específicas ultraestructurales del rotífero B. bidentatus son notorias, más aun en el macho y en el quiste. En cuanto a la ultraestructura de los espermatozoides y los bastones respecto a otras especies sólo difieren en su tamaño, a pesar de las similitudes estructurales las estructuras de los bastones se evidencian con mayor claridad y dan más evidencias sobre su funcionalidad. Nuestro estudio de la ultraesturctura de esta especie añade información útil que junto con un análisis molecular ayudarán a clarificar la taxonomía de rotíferos brachionidos.
Palabras clave: Rotifero, Brachionus bidentatus, ultraestructura, varillas, espermatozoide
The study of rotifer ultrastructure is still important for two reasons: (1) there is still a paucity of information regarding basic rotifer morphology, (2) detailed study of rotifer trophi using SEM is an useful tool to help differentiate cryptic species (Segers, Murugan & Dumont, 1993). In the past, the description of new species was based on light microscopy; nowadays, we have known from the work of molecular genetics that many species are really species complexes (Gilbert & Walsh, 2005; Wallace, Snell, Ricci & Nogrady, 2006). Brachionus bidentatus is a freshwater rotifer that has several polymorphic characteristics (Koste, 1978; Silva-Briano 2000). Recently, one of these variants, having two anterior wing-like projections, was described as a new species after experiments demonstrated that these projections were not the result of phenotypic plasticity (Silva-Briano, Galván-de la Rosa, Pérez-Legaspi & R. Rico-Martínez 2007). Rotifer ultrastructure has been studied since 1980 (Wallace et al., 2006), but unfortunately has been restricted to very few species. Perhaps the work by Clément & Wurdak (1991), is the most complete, and summarized findings in ultrastructure and anatomy. Their work includes transmission electron microscopy (TEM) and scanning electron microscope (SEM) photographs of the different structures found in eight species: Asplanchna brightwellii, Brachionus calyciflorus, B. plicatilis, B. sericus, Notommata copeus, Philodina roseola, Rhinoglena frontalis, and Trichocerca rattus. Recently, many studies have focused on the ultrastructure of the trophi, giving the importance of this organ in the phylum (Wallace et al., 2006).
In rotifers, two types of cells have been identified in the sperm vesicle: spermatozoa and rods (Clément & Wurdak, 1991). The study of the ultrastructure of the spermatozoa in rotifers has been limited to the genera Asplachna and Brachionus (Koehler, 1965; Clément, 1977; Clément & Wurdak, 1991). Melone & Ferraguti (1993) described with details the ultrastructure of the sperm in Brachionus plicatilis. The cellular body of the spermatozoon has several vesicles; the nucleus is partially condensed, and has several mitochondria and some cytoplasm. On the other hand, the rods are cells that lack flagellum and nucleus and have great secretory activity. They are composed of a dense homogeneous substance originated in the Golgi apparatus (Clément & Wurdak, 1991). The studies regarding rotifer ultrastructure in general and sperm cells in particular are quite limited. The goal of this work was to study the ultrastructure of female and male B. bidentatus, and the three types of eggs (parthenogenetic, sexual unfertilized and sexual fertilized) using SEM and TEM. We also analyzed the trophi and the spermatic cells, and compared our findings with those previously published.
Materials and methods
Maintenance of the Brachionus bidentatus culture: Specimens of B. bidentatus were collected from La Punta reservoir (22°08’N - 102°24’W) located in the municipality of Cosío in Aguascalientes State, Mexico. Rotifer cultures were maintained in petri dishes with EPA medium at pH=7.5 (U.S. Environmental Protection Agency, 1985), that were kept in a bioclimatic chamber with a photoperiod of 16:8L: D, at 25ºC. The cultures were fed 1x106cells/mL of the chlorophyte Nannochloris oculata grown in Bold´s Basal Medium (Nichols, 1973).
Egg production of Brachionus bidentatus females: Neonate females were isolated at birth and placed in separate wells of a 24-well polystyrene plate (Costar Co.). Females (N=13), were monitored every two hours from birth to death. The number and type of eggs produced by each female were recorded.
Preparation of samples for SEM: Fifty female specimens, 50 male specimens, and 50 cysts of the freshwater rotifer B. bidentatus previously fixed in 4% formaldehyde, were dehydrated in graded series of ethanol, then took them to critical point and mounted in an aluminum stub (1cm high and 1.2cm in diameter), and covered with gold. To study the trophi, 25 B. bidentatus females were prepared according to the protocol of Segers et al (1993) with slight modifications. Briefly, this protocol consisted in isolating 25 females in a Petri dish. Then adding a drop of sodium hipochloride, and wait until the lorica was dissolved. Then the females were washed three times with distilled water and mounted in a SEM cylinder. The specimens were observed in a JEOL 5900 LV scanning electronic microscope.
Preparation of male specimens for TEM: Males of B. bidentatus (N=500) were fixed in 2% Glutaraldehyde (GTA) and 4% Paraformaldehyde (PFA) with 0.16M phosphate buffer (PBS). Then, they were post-fixated with 1% osmium tetraoxide (OSO4). Later, the males were embedded in epoxic resin (EPON) and observed in a JEOL 1010 transmission microscope operated in 80kv.
Results
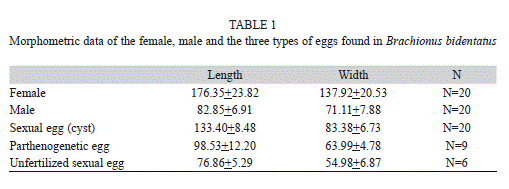
Life cycle of Brachionus bidentatus: In laboratory cultures females of B. bidentatus have a lifespan of five days (mean±one SD=4.69±0.48; N=13), and produce 4.5±3.67 (N=6) parthenogenetic eggs during such lifespan. Unfertilized sexual females produce up to 18 male eggs (mean±one SD=13±4.93; N=7). Once fertilized, sexual females produce a single diapausing embryo or cysts (mean±one SD=1±0; N=20). The length of time of copulation ranges from 10 to 40s (mean±one SD=17.33±10.55, N=7). The morphometric measurements of the adult females, males, and the three types of eggs present in B. bidentatus are shown in Table 1.
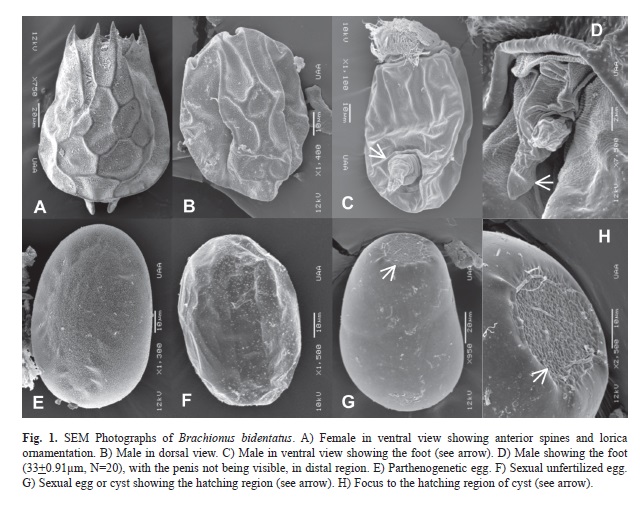
Males can reach up to 90µm in length (Table 1). However their loricas lack ornamentation and spines as is present in females (Fig. 1C). The male lorica, as in the female, is composed of the dorsal and ventral plates. The dorsal plate has a peculiar ridge. No trophi was observed in the male. Regarding the reproductive apparatus, and in contrast with other males described previously (even within the genus), where a foot and penis are clearly seen, in B. bidentatus we observed only the foot (Fig. 1D) whose mean length is 33µm (Table 1). Therefore, the penis must have been retracted or hidden by the techniques used in preparing the male specimens.
The parthenogenetic eggs are oval with approximately 100µm length (Table 1). They have a smooth surface and round contour (Fig. 1E). A female is able to produce seven to 10 eggs during her lifespan. Usually they are unable to carry more than three eggs at a time (N=5).The length of non-fertilized, sexual eggs (male eggs) is 80µm (Table 1); they possess a smooth surface, but show some borders and ridges (Fig. 1F). The fertilized sexual eggs (diapausing embryos) are oval with a very peculiar structure in their anterior part (Fig. 1G, see arrow); the surface of the egg is smooth with a small rugose surface from which neonates hatch. Inside of this hatching area, the rugose surfaces appear to be composed of small entangled membranes (Fig. 1H).
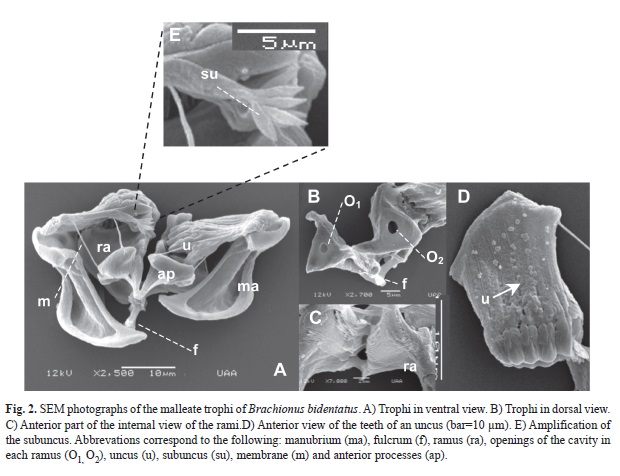
Ultrastructure of the trophi: The trophi of B. bidentatus is malleate with bilateral symmetry, and it is composed of manubria, fulcrum, rami and unci (Fig. 2). Each manubrium is joined to its respective unci (Fig. 2A and B), which provides support while processing food. The unci have five teeth disposed from the smallest to the largest teeth in dorsal view (Fig. 2D). On the other hand, a sixth segmentation is observed appearing to be a sixth tooth, but it is in fact, the projections of the subuncus, which shows a series of small teeth similar to the bristles of a toothbrush (Fig. 2E). The largest tooth is linked to the anterior process through a fine membrane. A third more robust membrane joins the anterior processes to the posterior part of each rami. The ramus is the structure where the teeth of each uncus collide. In the internal part of the uncus there are multiple rows of thin ridges that work in the grinding process (Fig. 2C). Finally, the fulcrum is a cylindrical prolongation that is joined to the rami (Fig. 2F).
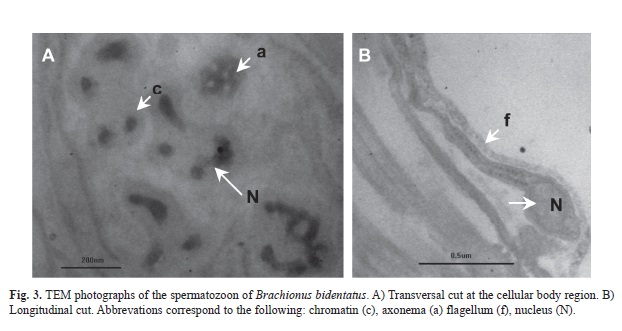
Ultrastructure of the sperm cells: The spermatozoon of B. bidentatus are composed of a body cell and a flagellum. The length of the cellular body is 300nm, while the flagellum is 1 700nm long (Fig. 3B). The nucleus of the body cell shows dense areas of chromatin and an axonema in the process of disintegration (Fig. 3A). Spermatozoan are located in specific regions of the rotifer forming a conglomerate without mixing with other cells and always surrounded by a large amount of rough endoplasmic reticulum and free ribosomes.
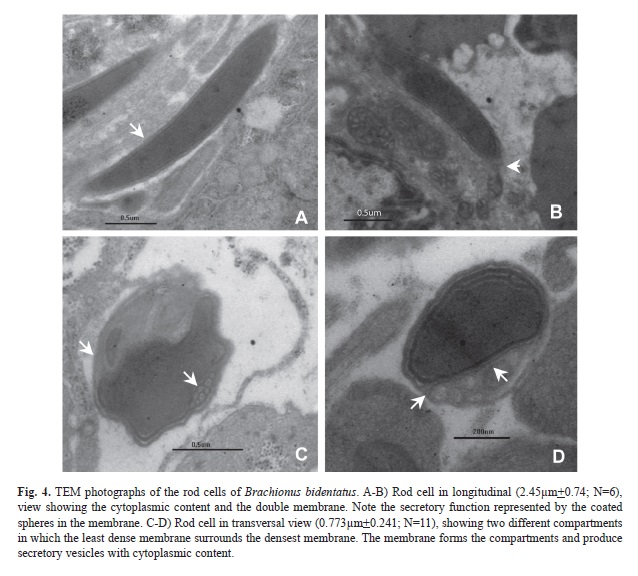
The rods have double membrane. Their length is almost 2.45μm and they are less than 1μm wide (Fig. 4). In a transversal view, they have two sections: one very electrodense, in which a membrane is divided in small vesicles separated by its own membrane that in turn is covered by a less dense membrane. The second section, which is the densest area of the rod, has small vesicles in which the cytoplasmic content can be seen. The least dense region of the rod also presents a vesicular arrangement, however these vesicles may occupy the whole area of the rod. Double membranes can be observed in Fig. 4. The least electrodense membrane covers the whole content of the rod including the densest membrane, which only surrounds the cytoplasmic content. In Fig. 4B and C, it is possible to observe that the space surrounded by the least dense membrane is totally occupied. In this case, the small vesicles with cytoplasmic content start to separate. These vesicles are shaped spherically and can be confused with the ultrastructure of the mitochondria. Once the vesicles are separated from this region, they become elongated and are transported to a cavity in which some flagella are found.
Discussion
The study of B. bidentatus ultrastructure and sexual features have shown its importance as it allows to determine subtle, yet important, differences in sexual reproductive behavior and structure that contribute to the general knowledge of this genus, the best studied in rotifers.
The cysts are not only part of the sexual cycle, but they are also capable of withstanding unfavorable conditions, thus being reservoirs of genetic diversity (Gómez & Carvalho, 2000). They are characterized by differences in morphology and a darker color with respect to the parthenogenetic eggs based on observations of light microscopy; however, many other differences are clear when they are observed with electronic microscopy (Pourriot & Snell, 1983). The morphology and ornamentation of the cyst observed at the electronic microscope differs in each species, and therefore can be used to clarify the relationship among cryptic species (Gilbert & Wurdak, 1978). The ornamentation of the B. bidentatus cyst showed several specific characteristics: there is a granular area in the anterior part of the cyst surrounded by a membrane envolving the content of the cyst.
Females of B. bidentatus are widely distributed throughout the State of Aguascalientes (Silva-Briano & Adabache Ortiz, 2000). The characteristics of the female are clearly described by Koste (1978). The length of the female lorica ranges between 153-578µm, which agrees with Stemberger´s range of 175-578µm (Stemberger, 1979), and reports of mean lorica length of 315µm (Rico-Martínez, 1999). The mating behavior of B. bidentatus was briefly described by De Ridder (1991), and Rico-Martínez (1999). The latest author reports a mean lorica width of 65µm, and a mean number of 4.58 granular cells for the male. However, in our work the male mean lorica length was 82.85µm, 71.11µm of mean lorica width, and a mean number of five granular cells. We found many similarities in the mating behavior of B. bidentatus with that of other Brachionus species. One difference is that the male is quite small (less than 100µm long). Many rotifers males lack mastax, or if present it is degenerated. The same is true for pharynx, stomach and gastric cells (Gilbert, 1993; Snell & Wallace, 2010). In the case of the male B. bidentatus, the mastax is absent, but we were not able to identify the stomach using TEM and SEM. When comparing the B. bidentatus male with that of B. araceliae included in the work of Silva-Briano et al. (2007), the differences in size and general morphology were clear.
Males of B. bidentatus show a region where the sexual cells are found (photographs not included), these cells are the spermatozoa and the rods. The number of sexual cells own by a male ranges from 30 to 200 (Wallace et al., 2006). In B. plicatilis the spermatozoa are 40 to 45µm long, and possess a cellular body and a flagellum (Melone & Ferraguti, 1993). These spermatozoa are larger than that of the spermatozoa of B. bidentatus which reaches only 2µm long. The sexual cigar-like appearance cells are known as rods. However, in B. bidentatus the rods are only 2µm long, much shorter than those of B. plicatilis, which are each 15µm long (Melone & Ferraguti, 1993). The secretory function of the rods has been described earlier (Clément & Wurdak 1991), and in the present study we confirmed this finding, where transversal cuts allowed the visualization of a dense region with cytoplasmic content originated in the Golgi Apparatus (Clément & Wurdak 1991). Clément & Wurdak (1991) have described a double membrane involving the rods of Brachionus sericus such membranes were also observed in B. bidentatus.
The observations of B. bidentatus ultrastructure presented here suggested that there are differences in general morphology, trophi, and sexual cells among members of the same genus. Even in the case of B. araceliae, which was previously described as a morphotype of B. bidentatus, the differences are striking. Therefore, if we are to achieve a deeper comprehension understanding of rotifer biology, additional studies of their ultrastructure is required.
Acknowledgments
The authors acknowledge the help of Maria de Lourdes SeguraValdez and Luis Felipe Jiménez Garcia, and also to Araceli Adabache-Ortiz for her help with the SEM images.
References
Clément, P. (1977). Ultrastructural research on rotifers. Hydrobiologia, 8, 270-297. [ Links ]
Clément, P. & Wurdak, E. S . (1991). Rotifera. In F. W. Harrison & E. E. Ruppert (Eds.). Microscopic Anatomy of Invertebrates, Vol. 4: Aschelmintes (219-297). New York: Wiley-Liss. [ Links ]
De Ridder, M. (1991). Rotifers from Algeria. Journal of African Zoology, 105, 734-783. [ Links ]
Environmental Protection Agency. (1985). Methods for measuring the acute toxicity of effluents to freshwater and marine organisms. In W. H. Peltier & C. I. Weber (Eds.). EPA-600/485-013. Washington, D.C., USA. Environmental Protection Agency. [ Links ]
Gilbert, J. J. (1993). Rotifera, Asexual Propagation and Reproductive Strategies. In K. G. Adiyodi & R. G. Adiyodi (Eds.). Reproductive Biology of Invertebrates, Volumen VI, part A (p. 231-263). New Delhi: Oxford and IBH Publishing Co. [ Links ]
Gilbert, J. J. & Wurdak, E. S. (1978). Species-specific morphology of resting eggs in the Rotifer Asplancha. Transactions of the American microscopical Society, 97, 330-339. [ Links ]
Gilbert, J. J., & Walsh, E. J. (2005).Brachionus calyciflorus is a species complex: Mating behavior and genetic differentiation among four geographically isolated strains. Hydrobiologia, 546, 257-265. [ Links ]
Gómez, A. & Carvalho, G. (2000). Sex, parthenogenesis and genetic structure of rotifers: microsatellite analysis of contemporary and resting egg bank populations. Molecular Ecology, 9, 203-214. [ Links ]
Koehler, J. (1965). An electron microscopic study of the dimorphic spermatozoa of Asplanchna (Rotifera). I. The adult testis. Zeitchrift für Zellforschung, 67, 57-76. [ Links ]
Koste, W. (1978). Rotatoria. Ein Bestimmungswerk, begründet von Max Voigt Überordning Monogononta. (Ed.). Gebrüder Borntraeger. Berlin. Stuttgart. 10. [ Links ]
Melone, G. & Ferraguti, M. (1993). The Spermatozoon of Brachionus plicatilis (Rotifera, Monogononta), with Some Notes on Sperm Ultraestructure in Rotifera. Acta Zoologica 75(2), 81-88. [ Links ]
Nichols, H.W. (1973). Growth media-freshwater. In J. R. Stein (Ed.). Handbook of physiological methods (p. 7-24). Cambridge MA: Cambridge University Press. [ Links ]
Pourriot, R. & Snell, T. W. (1983). Resting eggs in rotifers. Hydrobiologia, 114, 213-224. [ Links ]
Rico-Martínez, R. 1999. Final Report of the project Characterization of mating behavior in several freshwater species of rotifers. International Foundation for Science (IFS). Stokholm, Sweden. 39 p. [ Links ]
Segers, H. Murugan, G. & Dumont, H. (1993). On the taxonomy of the Brachionidae: description of Plationus n. gen. (Rotifera, Monogononta). Hydrobiologia, 268, 1- 8. [ Links ]
Silva-Briano, M. & Adabache-Ortiz, A. (2000). Brachionus species in Aguascalientes State, México. In M. Munawar (Ed.). Aquatic Ecosystems of México. (p. 203- 211). Burlington, Ontario, Canada. Ecovision World Monograph series. [ Links ]
Silva-Briano, M., R. Galván-de la Rosa, A. Pérez-Legaspi & R. Rico-Martínez. (2007). On the description of Brachionus araceliae sp.nov. A new species of freshwater rotifer from Mexico. Hidrobiológica, 17(2), 179-183. [ Links ]
Stemberger, R.S. (1979). A guide to rotifers of the Laurentian Great Lakes. EPA-600/4-79-021. Washington, D.C.: U.S. Environmental Protection Agency. [ Links ]
Snell, T.W. & Wallace, R.L. (2010). Chapter 8: Rotifera. In J.H. Thorp & A. Covich. (Eds.). Ecology and Classification of North American Freshwater Invertebrates (p. 173-206). New York: Academic Press. [ Links ]
Wallace, R.L., Snell T.W., Ricci. C. & Nogrady T. (2006). Rotifera Part 1: Biology, Ecology and Systematics. Ghent, Belgium: Kenobi Productions. [ Links ]
*Correspondencia a:
1Gerardo Guerrero-Jiménez. Universidad Autónoma de Aguascalientes, Laboratorio de Toxicología Acuática, Edificio 60. Depto. Química. Avenida Universidad 940, C.P. 20131, Aguascalientes, Ags., México
2Guadalupe Zavala-Padilla. Universidad Nacional Autónoma de México. Instituto de Biotecnología. Laboratorio de Microscopía.Av. Universidad #2001, Col. Chamilpa C.P. 62210 Cuernavaca Morelos. México; gzavala@ibt.unam.mx
3Marcelo Silva-Briano. Universidad Autónoma de Aguascalientes Laboratorio de Ecología. Edificio 202. Depto. Biología. Avenida Universidad 940, C.P. 20131, Aguascalientes, Ags., México; msilva@correo.uaa.mx
1Roberto Rico-Martínez. Universidad Autónoma de Aguascalientes, Laboratorio de Toxicología Acuática, Edificio 60. Depto. Química. Avenida Universidad 940, C.P. 20131, Aguascalientes, Ags., México; rrico@correo.uaa.mx
Received 22-I-2013. Corrected 20-IV-2013. Accepted 27-V-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}