










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.4 San José Oct./Dec. 2013
Declining densities and reproductive activities of the queen conch Strombus gigas (Mesogastropoda: Strombidae) in Banco Chinchorro, Eastern Caribbean, Mexico
Disminución de la densidad y actividades reproductivas de la concha reina Strombus gigas (Mesogastropoda: Strombidae) en el Banco Chinchorro, Caribe Oriental
Disminución de la densidad y actividades reproductivas de la concha reina Strombus gigas (Mesogastropoda: Strombidae) en el Banco Chinchorro, Caribe Oriental
*Dirección para correspondencia:
Abstract:
Queen conch is a gastropod inhabiting the Caribbean Sea, it represents the second largest fishery after the spiny lobster, but it has been extensively captured in the area. In order to know its population status in Chinchorro Bank, we determined conch density changes and its effects on reproductive activities, between July and November 2009. For this, data on conch density, morphology and reproductive activities were obtained from 15 sites within three fishing zones, and compared with previously collected data (1990, 1992, 1994, and 1997). Data showed that adult density decreased with time, from 10 700ind./ha in 1990, to 198ind./h in 2009. Neither egg masses nor spawns were found and mating was only observed once in July 2009. In July, adult (lip>4mm) density in the Southern zone was 23ind./ha whereas in the Northern zone and Central zone densities were 15 and 9ind./ha respectively. In November, density was somewhat higher: Southern zone 96ind./ha; Central zone 39ind./ha and Northern zone had 38ind./ha. In July, mean shell length was 170.80±46.28mm, with a higher median abundance at 180-189mm. In November, higher frequency was 187.63±45.14mm, maximum at 210-219mm interval. For the last 10 years period, mean adult conch densities have diminished in each zone, which might be the main cause of decreased reproductive activities of the conch at Banco Chinchorro. It is therefore an immediate need to analyse the management plan for this species in this Reserve and perhaps to promote a re-population of queen conch and culture activities. Rev. Biol. Trop. 61 (4): 1671-1679. Epub 2013 December 01.
Key words: Allee effect, distance sampling, mollusk, reproduction.
Resumen
El caracol Rosado es un gasterópodo que habita en el Caribe, y representa la segunda pesquería después de la langosta espinosa, sin embargo, el caracol ha sido pescado a niveles de casi extinción. Para conocer el estatus de la población se analizaron los cambios de densidad del caracol rosado en Banco Chinchorro y su efecto en las actividades reproductivas entre julio y noviembre 2009. Se recolectaron datos de densidad y morfología en tres zonas de pesca en 15 localidades; y se compararon con datos recolectados previamente en los mismos sitios (1990, 1992, 1994, y 1997). Los datos mostraron que la densidad de adultos ha decrecido con el tiempo de 10 700 (ind./ha) en 1990 a 198 (ind./ha). Las actividades reproductivas o productos se registraron, no se encontraron ni masas de huevo ni desoves, y únicamente se observó una copulación en julio. Considerando únicamente a los adultos (labio>4mm), en julio 2009, la densidad fue de 23ind./ha, mientras que en la zona norte y centro las densidades fueron 15 and 9 (ind./ha). En noviembre 2009, la densidad fue ligeramente más alta: zona sur 96 (ind./ha); zona Centro 39 (ind./ha) y zona Norte 38 (ind./ha). En julio (2009) la longitud de concha promedio fue de 170.80±46.28mm, con una mayor abundancia en las tallas 180-189mm. En noviembre (2009) la frecuencia más alta ocurrió en las talla promedio 187.63±45.14mm, con un intervalo de 210-219mm de longitud de concha. En los últimos 10 años, la densidad de los caracoles adultos ha disminuido en cada una de las zonas de pesca, lo cual podría ser la causa de la disminución de las actividades reproductivas del caracol rosado en Banco Chinchorro.
Palabras clave: Allee effect, distance sampling, moluscos, reproduction.
The Queen conch (Strombus gigas L. 1758), a gastropod inhabiting in the Caribbean Sea, has been extensively captured up to levels of near extinction (Appeldoorn, Dennis & Monterrosa-Lopez, 1987; Berg & Olsen, 1989) as it represents the second largest fishery after the spiny lobster (Appeldoorn, 1994). World Queen conch landings have been calculated around 6 000 metric tons with a value of 60 million US dollars (Chakalall & Cochrane, 1997). Populations of S. gigas have been affected almost in all countries where they are exploited at a commercial scale (Brownell & Stevely, 1981; Davis & Hesse, 1983). This is even more evident in countries where the fishery has been open to SCUBA diving (Stoner & Schwarte, 1994). The exhaustion of stocks has forced, at least temporarily, closure of fishing of S. gigas in Bermuda, Florida, Cuba, Bonaire, and the Virgin Islands (Stoner & Schwarte, 1994). Since 1992 S. gigas is considered a threatened species by CITES (Appeldoorn, 1994).
Unfortunately, studies of reproductive activities are scarce for S. gigas and other gastropods. In Chile, Avendaño, Cantillánez, Olivares & Oliva (1997, 1998) studied reproductive activities of Thais chocolata and found that these gastropods group together for reproduction in a range from 29 to 376 individuals in shallow waters. In the Caribbean, the first studies on conch reproductive activities were realized in the Turk and Caicos Islands (Davis & Hesse, 1983) and Puerto Rico (Egan, 1985; Appeldoorn, 1993). In The Bahamas, Stoner & Ray (2000) described the reproductive activities of S. gigas and found a relation with population density. Their results indicate a sigmoid relationship between density of adults and their reproductive activities, while sites with smaller densities 56 (ind./ha.) showed no reproductive activity and reproduction reached an asymptotic level near 200 mature (ind./ha.). In 2003, Tewfik and colleagues studied the distribution and abundance of S. gigas and S. costatus at Bocas del Toro, Panama; their results indicated a low adult density and the authors point out the possibility of an Allee effect in conch population, as a result of unsustainable fishing activities. Gascoin & Lipcius (2004) showed a direct relationship between the reproductive index (RI) and the density at the site of origin of conchs, linking it thereby to the Allee effect. Recently, Stoner et al. (2012) showed the impact of population change on the reproductive activities of Queen conch in The Bahamas.
For the Yucatan Peninsula we only found two studies dealing with Queen conch reproductive activities. One study was carried out in the Alacranes reef by Pérez-Pérez & Aldana-Aranda (2003) who showed that reproductive activity of S. gigas occurs in the period from January to October. Aldana-Aranda, Baqueiro-Cárdenas, Martinez Morales & Ochoa Báez (2003) studied the reproductive behaviour of Queen conch using the gonadic development in Banco Chinchorro, and showed that during the five months of sampling, gametogenesis and mature organisms were present, whereas spawning occurred from June to August, with the highest peak in July. The evaluation of the reproductive activities of organisms under exploitation is extremely important, as extraction without control could limit the reproductive capacity, due to a reduction of the overall population size, or could influence the adult reproductive capacity (known as Allee effect) (Avendaño et al., 1998). The main objective of this research is to determine the density, distribution and size structure of the Queen conch population in a reef lagoon, as well as to analyse density changes over time and to relate these parameters to reproductive activities.
Materials and Methods
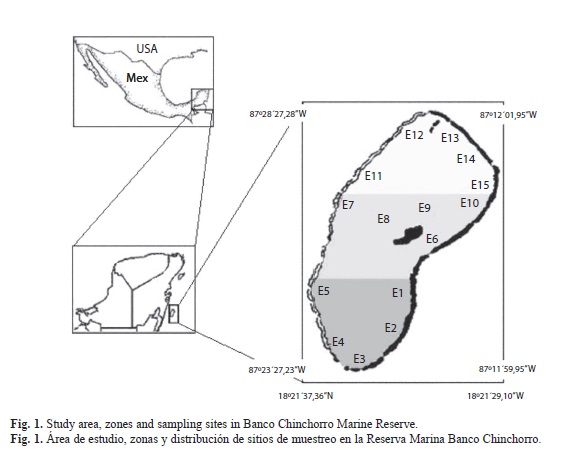
Study area: The Banco Chinchorro Marine Biosphere Reserve (18º47’, 18º14’N - 87º14’, 87º27’ W) is a reef complex located on the South-Eastern coast of Quintana Roo. The Reserve has a surface of 144 360ha and includes reef formations, a reef lagoon, and three keys: Lobos, Centro and Norte, and is located adjacent to oceanic waters (INE-SEMARNAP, 2000).
Sampling was carried out inside the reef lagoon during July and November, 2009 in three distinct zones: North (17 245ha), Central (21 110ha) and South (15 461ha), and at 15 sampling sites (E1, to E15), traditionally considered as fishing zones by local fishermen of S. gigas. Results were compared with data collected previously at the same sampling sites (1990, 1992, 1994, and 1997) (Fig. 1). At each station, triplicate transects of 100m x 3m were set using distance sampling. Distance sampling consists of using visual observation to evaluate population density and size. The perpendicular distance of an organism along the transect was registered and this was used to calculate the detection function which gives the probability of an object or organisms, at a given distance, being observed in transect (Buckland, Anderson, Burnham & Laake, 1993).
To relate the frequency of reproductive events to the physical variables of the water in each area, the following variables were measured: temperature, oxygen concentration and salinity, according to De Jesús-Navarrete (1999) using an oxygen meter YSI model 550A with a precision of 0.01°C (temperature) and 0.1mg/L (oxygen). Salinity was measured using a portable refractometer Extech model RF20 with a precision of 0.1 using the Practical Salinity Scale.
To determine the relationship between water parameters and density, a correlation analysis (Pearson, 0.05) was undertaken. The software Distance Sampling 5.0 was used to calculate conch population densities in each station, and to estimate the overall number of conchs in each zone. A bi-factorial ANOVA was used to determine statistical significance of differences between population densities, sampling months, and zones (North, Central and South). A multifactorial ANOVA was used to show differences between population density, zones and years using Statgraphics 6.0.
Results
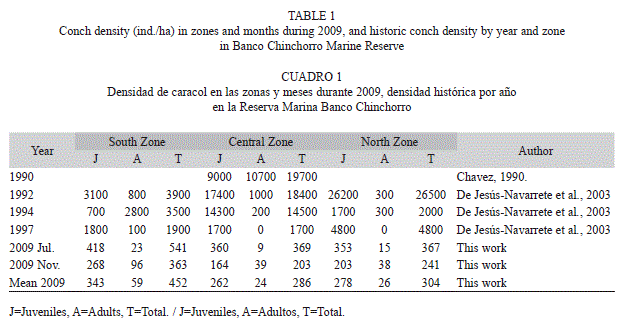
Total conch density: A total area of 48 600m2 was examined (81 transects). During July, conch densities at South, Central, and North zones were 541, 369 and 367ind./ha, respectively. In November, the South zone had the highest density 363ind./ha; in the Central and North zones the densities were 203 and 241ind./ha respectively. Considering only adult specimens (lip>4mm), in July we observed a density of 23ind./ha, in the South zone, followed by the North Zone with 15ind./ha. The Central Zone had the lowest density of 9ind./ha. During November, adult numbers were slightly higher; the South zone exhibited the highest density in the period (96ind./ha), and the Central and North zones had 39 and 38ind./ha, respectively. Comparing annual changes in conch densities, there was a trend towards low population densities overall in adult organisms (Table 1). When total conch density was analysed, no significant differences between months and zones (p=0.74, F=0.11) were found. However, when considering only the adult fraction, a significant statistical difference was shown between adult density in each season (p=0.0149). Comparing density, years and age classes using multifactorial ANOVA showed significant differences in density between years, and age class (p<0.05). A LSD test however, showed no differences between 2009 and 1997.
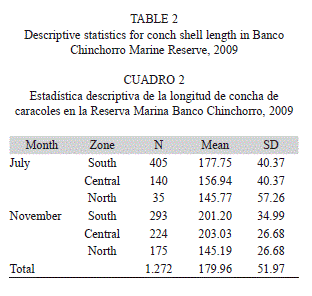
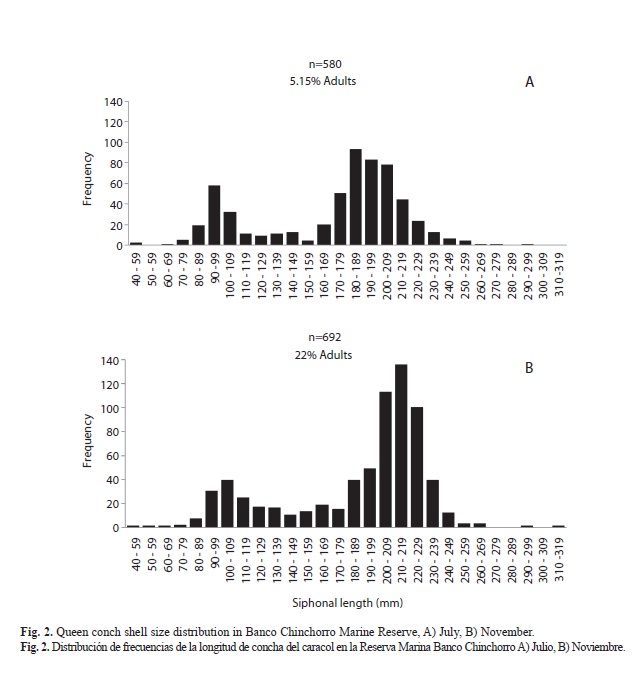
Size distribution: Shell length was measured for a total of 1 272 conchs in the two study periods. Shell sizes varied from 41mm to 314mm with a mean length of 179.96±51.97mm (Table 2). In July, 580 conchs were measured, with a mean length of 170.80±46.28mm. In November, 692 conchs were measured, indicating a small increase in mean shell length (187.63±45.15mm) (Table 2). Considering the zones, the mean lengths obtained were 187.60±39.89, 185.30±41.27 and 145.29±52.74mm for the Southern, Central and North zones, respectively (Table 2). In July, conch sizes varied from 41mm to 297mm, with a mean size of 170.80±46.28mm, the more frequent size being 180-189mm (93 conchs) (Fig. 2A), of which only 5.51% were mature. In November, 692 conchs were measured, of which 21.93% were mature. Shell length varied from 57mm to 314mm with a mean size of 187.63±45.14mm, the highest frequency being 210-219mm (136 conchs) (Fig. 2B).
Comparing conch density throughout the past 20 years, a decrease at all sites within the Chinchorro Bank was evident. Density variation was found to be three orders of magnitude lower than conch densities in 1990 (Table 1).
Physical parameters: Water temperature varied from 24.4 to 29.8°C during the sampling period, whereas oxygen varied from 6.08 to 6.5mg/L and salinity fluctuated from 35.0 to 38.5 UPS. No correlation between conch density and physical parameters was found (Pearson correlation, r2<0.50).
Discussion
Various authors have mentioned that conch distribution and reproduction depend on several factors, mainly temperature and dissolved oxygen in the water column, as well as sediment type and microalgal biomass at the sea bottom; additionally, the presence of conspecifics has an important effect on reproductive activities (Appeldoorn, 1988; Davis & Hesse, 1983). Furthermore, there are variations during the year, related to seasonal changes. During this research we only collected data during two months corresponding to the dry and rainy seasons, but we could prove that a low population density and adult abundance influences the likelihood of reproductive encounters.
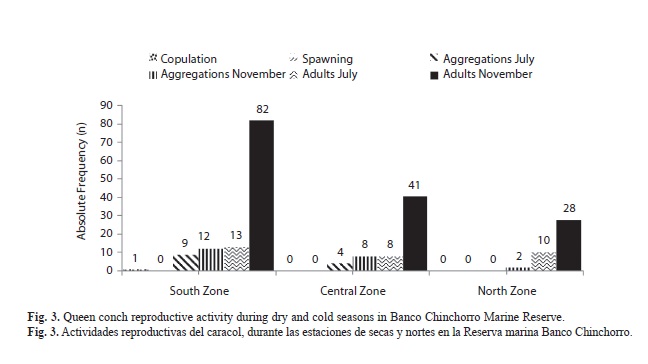
The basic premise in sustainable fishing is to capture only population surpluses, thereby avoiding damage to the overall reproductive capacity. When overexploitation disrupts this delicate balance, fishing activities cause alterations in the stock as they diminish total biomass, change mean conch size, alter the age of sexual maturity, and modify sex proportions and genetic structure (De Jesús-Navarrete, Medina-Quej & Oliva-Rivera, 2003). The evaluation of changes in density is of vital importance to determine if fishing activities are causing changes in the adult population, which could affect reproductive activities and thereby producing an Allee effect. Changes in population density at Banco Chinchorro are evident; populations had diminished almost 30% in 2009 compared with the same sites densities in 1990. The densities found during the research activities for this article can only confirm a decrease in population density and mean siphonal length of conchs. Additionally, we found that a decreased density of adult means fewer individuals for reproduction and consequently a decrease in the probability of adult encounters, or Allee effect (Berryman, 1999). Contrary to year-round reproduction of S. gigas at Banco Chinchorro, as reported (Cruz, 1986; Corral & Ogawa, 1987), we only observed a single copula, which could be the effect of a reduction in adult density.
In The Bahamas, Stoner & Ray (2000) reported that in populations with densities smaller than 56 and 48ind./ha, no copulation and spawning have been observed. Our results indicated that densities of mature conch in Banco Chinchorro are below densities reported by Stoner & Ray (2000), considering that in July the adult mean density in Banco Chinchorro was 15.5ind./ha and in November 57.65ind./ha, which might explain the absence of reproductive activities. This apparent difference between months might be explained by local conch movements or it could be a product of conch recruitment in the reef lagoon. This cannot be determined based on the data resulting from our research, however we must emphasise the presence of a high percentage of juveniles in the samples and this could represent recruitment of conch in the reef lagoon.
In a previous evaluation, De Jesús-Navarrete (1999) found egg mass variation abundance from 0.015 (egg masses/m2) in the North zone in October, up to 0.03egg masses/m2 in the Central zone in May. This contrasts with the absence of egg masses in this study, and apparently this decrease in reproductive activity is related to a low adult conch density, and should be considered carefully in the conch management plan. Our observations in the field also differed from those mentioned by Aldana-Aranda, et al. (2003) who reported June as the month with the highest peak in reproductive activity in Banco Chinchorro, revealed by histological techniques. In Bocas del Toro, Panama, Tewfik & Guzmán (2003) found a very low adult conch density 0.30 (ind./ha) and observed no reproductive activities, indicating the presence of an Allee effect as a consequence of a negative per capita growth rate in conch population. Tewfik, Guzmán & Jácome, (1998) reported a total density of 14.6 (ind./ha) with 58% adults, and they observed only one pair of conchs copulating, a female spawning and only one egg mass was registered. The authors also point out that low conch density is a consequence of over-exploitation. In Banco Chinchorro, even with higher adult conch densities than in Cayos Cochinos (Tewfik & Guzmán, 2003), we also observed a lack of reproductive activities.
In theory, conch are caught one year after the lip is formed, but differences exist between the time when conch reach the legal longitude (200mm) and sexual maturity (lip of 4mm width), which implies that the 90% of the population that is exploited corresponds to juveniles, with clear repercussions for the overall population (Berg & Olsen, 1989; Stoner et al., 2012). This difference could be demonstrated by our research in Banco Chinchorro, in which the mean shell length found was 170.80mm in July and 187.63mm in November, which is smaller than the effective legal size (200mm); thus an artificial selection induced by conch fishing may be caused. Another threat to conch population is illegal fishing, which has been estimated as similar to the legal catch, (8t/year) (De Jesús-Navarrete unpublished data); illegal fishing contributes to conch density decrease in Banco Chinchorro. In conclusion, densities of mature S. gigas were extremely low, and if this tendency continues then a negative effect on commercial fishing and an impact on conch biology, mainly on reproductive activity, will be observed in the short term. It is therefore necessary to analyse the management plan for this species in Banco Chinchorro immediately and perhaps to initiate a re-population of queen conch with cultured organisms.
Acknowledgments
We greatly appreciate the support of Banco Chinchorro Marine Reserve staff in the field data collection. This project was funded by CONANP-SEMARNAP. The comments of Frank Ocaña, Birgit Schmook, Gerald Islebe and two anonymous reviewers substantially improved the manuscript.
References
Aldana-Aranda, D., Baqueiro-Cárdenas, E., Martinez Morales, I. & Ochoa Báez I. (2003). Gonad Behavior During Peak Reproduction Period of Strombus gigas from Banco Chinchorro. Bulletin of Marine Science, 73 1, 241-248Links ]">.
Appeldoorn, R. S, Dennis, G. D. & Monterrosa-Lopez, O. (1987). Review of shared demersal resources of Puerto Rico and the Lesser Antilles region, p. 36-57. In R. Mahon (Ed.), Report and Proceedings of the expert consultation of shared fishery resources of the Lesser Antilles region. FAO Fish. Rep. 383. [ Links ]
Appeldoorn, R. S. (1988). Ontogenetic Changes in Natural Mortality rate of Queen Conch. Strombus gigas, (Mollusca: Mesogastropoda). Bulletin of Marine Science, 42(2), 159-165. [ Links ]
Appeldoorn, R. S. 1993. Reproduction, spawning potential ratio and larval abundance of queen conch off La Parguera, Puerto Rico. Manuscript, Report to Caribbean Fishery Management Council, San Juan. Puerto Rico. 20 pp. 1993. [ Links ]
Appeldoorn, R. S. (1994). Queen Conch management and research: status, needs and priorities. In R. S. Appeldoorn & B. Rodriguez (Eds.), Queen conch Biology, Fisheries and Mariculture (pp. 301-320). Fundación Cientifica Los Roques. [ Links ]
Avendaño, M., Cantillánez, M., Olivares, A. & Oliva, M. (1997). Conducta reproductiva de Thais chocolata (Duclos, 1832) (Gastropoda: Thaididae), en La Rinconada, Antofagasta-Chile: Causal de vulnerabilidad a la pesca. Revista de Biología Marina y Oceanografía, 3(2)2, 177-187. [ Links ]
Avendaño, M., Cantillánez, M., Olivares, A. & Oliva, M. (1998). Indicadores de agregación reproductiva de Thais chocolata (Duclos, 1832) (Gastropoda, Thaididae) en Caleta Punta Arenas (21°38’S-70°09’W). Investigaciones Marinas Valparaíso, 26, 15-20. [ Links ]
Ávila-Poveda, O. & Baqueiro-Cárdenas, E. (2006). Size at sexual maturity in the queen conch Strombus gigas from Colombia. Boletín de Investigaciones Marinas y Costeras, 35, 223-233. [ Links ]
Berg, Jr. & Olsen, O. (1989). Conservation and Management of Queen Conch (Strombus gigas L.) Fisheries in the Caribbean. In J. F. Caddy, (Ed.), Marine invertebrate fisheries: Their assessment andmanagement (p. 421-442). New York: John Wiley. [ Links ]
Berryman, A. A. (1999). Principles of population dynamics and their application, United Kingdom: Stanley Thornes, Cheltenham. [ Links ]
Brownell, W. N. & Stevely, J. M. (1981). The biology, fisheries, and management of the queen conch, Strombus gigas. Marine Fisheries Review, 4, 1-12. [ Links ]
Buckland, S. T., Anderson, D. R., Burnham, K. P, & Laake, J. J. (1993). Distance Sampling: Estimating Abundance of Biological Populations. England: Oxford University Press. [ Links ]
Chakalall, R. & Cochrane, K. (1997). The queen conch fishery in the Caribbean: An approach to responsible fisheries management. Proceedings of the Gulf and Caribbean Fisheries Institute 49, 531-554. [ Links ]
Chavez, E. A. (1990). An assessment of queen conch Strombus gigas stock of Chinchorro bank, Mexico. Proceedings of the Gulf and Caribbean Fisheries Institute 43, 1-23. [ Links ]
Corral, J. L. & Ogawa, J. (1987). Cultivo masivo de larvas de caracol Strombus gigas en estanques de concreto. Proceedings of the Gulf and Caribbean Fisheries Institute, 38, 345-351. [ Links ]
Cruz, R. C. (1986). Avances en la experimentación de producción masiva de caracol en Quintana Roo, México. Proceedings of the Gulf and Caribbean Fisheries Institute 37, 12-20. [ Links ]
Davis, M. & Hesse, R. C. (1983). Third world level conch mariculture in the Turks and Caicos Islands. Proceedings of the Gulf and Caribbean Fisheries Institute 35, 73-82. [ Links ]
De Jesús-Navarrete, A. (1999). Abundancia de larvas velígeras del caracol rosado Strombus gigas y su relación con las áreas de pesca en el Sur de Quintana Roo, México. (Informe técnico final. Proyecto 0420P-N). Consejo Nacional de Ciencia y Tecnología. 213 pp. [ Links ]
De Jesús-Navarrete, A., Medina-Quej, A. & Oliva-Rivera, J. J. (2003). Changes in the Queen Conch (Strombus Gigas L.) population structure at Banco Chinchorro, Quintana Roo, México, 1990-1997. Bulletin of Marine Science, 73(1), 219-229. [ Links ]
Egan, D. B. (1985). Aspects of the reproductive biology of Strombus gigas. (Master Thesis). University of Columbia Británica, Vancouver, Canada. [ Links ]
Gascoigne, J. & Lipcius, R. N. (2004). Conserving populations at low abundance: delayed functional maturity and Allee effects in reproductive behaviour of the queen conch Strombus gigas. Marine Ecology Progress Series, 284, 185-194. [ Links ]
INE-SEMARNAP. 2000. Programa de Manejo de la Biosfera Banco Chinchorro, México. Comisión Nacional de Áreas Naturales Protegidas. 193 pp. [ Links ]
Pérez-Pérez, M. & Aldana-Aranda, D. (2003). Actividad reproductiva de Strombus gigas (Mesogasteropoda: Strombidae) en diferentes hábitats del Arrecife Alacranes, Yucatán. Revista de Biología Tropical, 5 (Supl. 4), 119-126. [ Links ]
Stoner, A. W. & Schwarte, K. C. (1994). Queen conch, Strombus gigas, reproductive stocks in the central Bahamas: distribution and probable sources. Fishery Bulletin, 92, 171-179. [ Links ]
Stoner, A. W. & Ray, M. (2000). Evidence for Allee effects in an over-harvested marine gastropod: density-dependent copulation and egg production. Marine Ecology Progress Series, 202, 297-302. [ Links ]
Stoner, A. W., Davis M. & Booker C. 2012. Negative consequences of Allee Effect are compounded by fishing pressure: comparison of queen conch reproduction in fishing grounds and a marine protected area. Bulletin of Marine Science, 88, 89-104. [ Links ]
Tewfik, A., Guzmán, H., & Jácome, G. (1998). Assessment of the queen conch, Strombus gigas (Gastropoda: Strombidae) population in Cayo Cochinos, Honduras. Revista de Biología Tropical, 46, 137-150Links ]">.
Tewfik, A. & Guzmán, H. (2003). Shallow-water distribution and population characteristics of Strombus gigas and S. costatus (Gastropoda: Strombidae) in Bocas del Toro, Panama. Journal of Shellfish Research, 22 3, 789-794. [ Links ]
*Correspondencia a:
1Alberto De Jesús-Navarrete. Departamento de Aprovechamiento y Manejo de Recursos Acuáticos, El Colegio de la Frontera Sur, Unidad Chetumal, A.P. 424 Chetumal Quintana Roo, México.
2Adriana Valencia-Hernández. Licenciatura en Biología, Instituto Tecnológico de Chetumal, Av. Insurgentes 330 esquina Av. Andrés Quintana Roo, Chetumal, Quintana Roo, México; anavarre@ecosur.mx, diana_625@hotmail.com
Received 22-I-2013. Corrected 06-V-2013. Accepted 06-VI-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}