










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.4 San José Oct./Dec. 2013
Biology of Kalipharynx sp. (Trematoda: Digenea) metacercariae in Biomphalaria (Gasteropoda: Planorbidae) from Northeastern Argentina
Biología de metacercarias Kalipharynx sp. (Trematoda: Digenea) en Biomphalaria (Gasteropoda: Planorbidae) del noreste Argentino
Biología de metacercarias Kalipharynx sp. (Trematoda: Digenea) en Biomphalaria (Gasteropoda: Planorbidae) del noreste Argentino
*Dirección para correspondencia:
Abstract
In Argentina, no ecological studies have been reported on the infection parameters of Kalipharynx sp. metacercariae in planorbid snails. To this end, the aims of this study were: (i) to provide information on the population biology of Kalipharynx sp. metacercariae in the planorbid snails Biomphalaria tenagophila and B. occidentalis through the study of prevalence and intensity of larval infection during a seasonal cycle; (ii) to evaluate the effects of host shell size on prevalence and infection intensity, (iii) to evaluate the effect of infection intensity on cyst size. Samples were taken between June 2010 and April 2011 (encompassing all seasons) from a subtropical permanent pond in Corrientes City, Corrientes, Argentina. A total of 362 metacercariae (n=262 and n=100; from B. tenagophila and B. occidentalis respectively) were collected from 616 snails (n=466 and n=150 from B. tenagophila and B. occidentalis respectively). The metacercarial cysts were found in the digestive gland, mantle cavity, intestine and ovotestis. B. tenagophila showed a range of infection from 1 to 60 cysts per snail (mean=4.5±SD=9.9), and cyst diameter ranging between 255 and 705μm (466.4±119); while, B. occidentalis showed a range of infection from 1 to 23 (5.5±5.6), and cyst diameter ranging between 310 to 900μm (554.5±150). Results obtained indicated that, although absent in autumn, metacercariae of Kalipharynx sp. were present most of the year in both species of Biomphalaria, showing high values in both warm-season. Furthermore, both the infection intensity and host shell size varied significantly between seasons, although no seasonal variation was observed with respect to metacercarial cyst size, suggesting the possibility of more than one peak of cercariae emergence during the year. The prevalence of infection was significantly and positively correlated with snail size in both host species (p<0.05). The smallest host size class harbouring a metacercarial infection was 7.1-8.0mm and 11.1-12.0mm in B. tenagophila and B. occidentalis, respectively. The mean intensity of infection was positively correlated with snail size, but this relation was significant only in B. tenagophila (p<0.05). However, non-significant negative correlations were observed for intensity of infection vs cyst size in both host species (p>0.05). The results of this study show a significant influence of host size on prevalence and infection intensity, and a tendency towards density-dependent reductions in the growth of cysts. This is the first study in Argentina analyzing the population biology of Kalipharynx sp. metacercariae.Rev. Biol. Trop. 61 (4): 1647-1656. Epub 2013 December 01.
Key words: larval trematodes, Kalipharynx, Biomphalaria, population, Argentina.
Resumen
En Argentina, no hay estudios ecológicos que hayan reportado los parámetros de infección de metacercarias Kalipharynx sp. en caracoles planorbídeos. Por lo que los objetivos de este estudio fueron: (i) proporcionar información sobre la biología de la población de metacercarias Kalipharynx sp. en los planorbídeos Biomphalaria tenagophila y B. occidentalis, a través del estudio de la prevalencia e intensidad de la infección por larvas durante un ciclo estacional, (ii) evaluar los efectos del tamaño del hospedador sobre la prevalencia y la intensidad de la infección (iii) evaluar el efecto de la intensidad de infección sobre el tamaño del quiste. Entre junio 2010 y abril 2011 se recolectaron en una laguna semipermanente de Corrientes, Argentina, un total de 616 caracoles (n=466 y n=150 de B. tenagophila y B. occidentalis, respectivamente) de los cuales se extrajeron 362 metacercarias (n=262 y n=100; de B. tenagophila y B. occidentalis, respectivamente). Se encontraron quistes en glándula digestiva, cavidad del manto, intestino y ovotestis. B. tenagophila y B. occidentalis estuvieron infectados con metacercarias la mayor parte del año, excepto en otoño, con valores altos en las estaciones cálidas (primavera-verano). La intensidad de infección y el tamaño del hospedador mostraron diferencias significativas entre las tres estaciones analizadas (invierno-verano), aunque no se observaron variaciones estacionales con respecto al tamaño de los quistes, sugiriendo la posibilidad de más de un pico de emergencia de las cercarias durante el año. La prevalencia se correlacionó positiva y significativamente con el tamaño del hospedador en ambas especies (p<0.05). La intensidad media de infección se correlacionó positivamente con el tamaño del hospedador, pero esta relación fue significativa sólo en B. tenagophila (p<0.05). Sin embargo, se observaron correlaciones negativas no significativas entre la intensidad de infección y el tamaño de los quistes en ambas especies (p>0.05). Los resultados obtenidos muestran una influencia significativa del tamaño del hospedador sobre la prevalencia y la intensidad de infección y una tendencia hacia reducciones denso-dependientes en el crecimiento de los quistes. Es el primer estudio que analiza la biología poblacional de metacercarias Kalipharynx sp. en caracoles planorbídeos de Argentina.
Palabras clave: larvas de tremátodo, Kalipharynx, Biomphalaria, población, Argentina.
Digenetic trematodes have a complex life cycle that may include three intermediate hosts and several developmental stages, e.g. miracidium, cercaria, metacercaria and mesocercaria (Esch & Fernandez, 1994; Esch, Barger & Fellis, 2002). In general, most of cycle occurs in the aquatic environment. The miracidia swim until their first molluscan intermediate host, and develop different larval stages (rediae, sporocysts), depending on the trematode species, until they reach the cercarial stage. The free-swimming cercariae escapes from the first intermediate host, and penetrate in the body of the compatible second intermediate host (aquatic invertebrates or aquatic vertebrates), where becomes a metacercaria that may, or may not, encyst. Freshwater snails of genus Biomphalaria serve as first and/or second intermediate host in the life cycle of some trematode superfamilies (Toledo & Fried, 2010). During this cycle, intramolluscan development is abiotic factors dependent, such as water temperature (Fried & Graczyk, 1997). Indeed, the development of cercariae in snails from mid-latitude climates, e.g. cold-temperature systems, shows a marked seasonality (Chubb, 1979). However, these conditions may vary among regions with subtropical climates, where the temperature is not a significant factor in determining strong seasonal patterns of trematode development (Esch & Fernandez, 1994; Hamann, 2004, 2006; Hamann, Kehr & González, 2010). In this sense, ecological studies regarding larval trematodes in planorbid snails from subtropical environments, have demonstrated that the short winter favours the development of larval trematodes the majority of the year (Ostrowski de Núñez, Hamann & Rumi, 1990, 1991).
Important factors in the transmission of larval trematodes, as the prevalence and intensity of infection, may be related to intermediate host size. Numerous field studies in marine and freshwater snails showed a positive correlation between prevalence of cercarial infection and snail size (see review Sorensen & Minchella, 2001; Graham, 2003). In addition, some studies showed an increase in the prevalence and mean intensity of metacercarial infection among larger snails (Morley, Lewis & Adam, 2004; Flores, Semenas & Veleizán, 2010). These relationships may be attributed to age-snails; older snails often are larger than young snails, within a given population, then larger-old snails may have been more time exposed to miracidia or cercariae (Minchella, Leathers, Brown & Mcnair, 1985; Graham 2003). On the other hand, parasites may interact within host, for example by reducing the growth of metacercariae when the intensity of infection increases in the intermediate host (Shandland & Goater, 2000; Brown, Lorgeril, Loly & Thomas, 2003; Saldanha, Leung, & Poulin, 2009).
In the present study, research was undertaken to test the following hypotheses: (1) the geographic location of the study area shows no marked seasonality in population of parasitic trematodes, (2) the level of larval infections are linked to the host size and size of metacercarial cysts. These premises were tested by analyzing the population biology of metacercariae of Kalipharynx Boeger & Thatcher, 1983 in the planorbid snails Biomphalaria tenagophila (D´Orbigny, 1835) and B. occidentalis Paraense, 1981. Both host species inhabits in routed and free floating plants forming part of benthos coast and pleuston from wetlands (Poi de Neiff & Neiff 2006; Poi de Neiff, Galassi & Franceschini, 2009). B. tenagophila has a wide distribution in South America reaching the Northeastern and Center of Argentina, whereas B. occidentalis has a much more limited distribution in South America and Argentina, in this last restricted to Corrientes province (Rumi, 1991; Rumi, Gutiérrez, Núñez & Darrigran, 2008). The genus Kalipharynx is considered as incertae sedis because it is not yet clear in which family it should be included (Pojmanska, Tkach & Gibson, 2008). Adult specimens of the type species of this genus (K. piramboae Boeger & Thatcher, 1983) were found in Chaco province, Argentina (Lunaschi, 1994) parasitizing lungfishes Lepidosiren paradoxa Fitzinger, 1837. The life cycle of this parasite is not known, but cercariae and metacercariae are reported with similar morphological characteristics parasitizing snails of genus Biomphalaria in Corrientes province, Argentina (Ostrowski de Núñez, et al., 1990). In Northeastern Argentina, no ecological studies regarding the infection parameters of metacercariae and the host-parasite relationships in these planorbid snails, it also represents the first study analyzing the population biology of Kalipharynx sp. metacercariae. To this end, the aims of the study were: (1) to provide information about seasonal variation of the prevalence and intensity of metacercarial infection in both snail hosts; (2) to analyze the effects of host shell size on prevalence and the intensity of larval infection, and (3) to evaluate the influence of the intensity of parasite infection on the size of metacercarial cysts.
Materials and Methods
Study area: The study area was a semipermanent pond located approximately 3km from the city of Corrientes, Corrientes, Argentina (27°29’24.20” S - 58°45’00.84” W). It is oval-shaped pond with 291.8m long, 121.1m wide and 0.7m deep. The predominant aquatic vegetation at the littoral zone is composed of rooted (Ludwigia uruguensis, L. peploides, Ludwigia sp.) and floating (Pistia stratoides, Hydroystria laevigata, Salvinia biloba, Azolla filiculoides, Lemna minuta) hydrophytes, and several species of grasses (Panicum sp., Elionurus muticus). The mean annual temperature is 23°C, with a mean temperature in the warmer month (January) of 26-27°C and a mean temperature in the coldest month (July) of 13-16°C (Carnevali 1994). This area is surrounded by human activities, e.g. housing, hospital and parks. The deep of the littoral zone ranged between 45cm (June 2010) and 10cm (April 2011) during sampling period. The water temperature varied between 13.1°C (July) and 28.7°C (November). Table 1 shows values of temperature and mean precipitation by season.
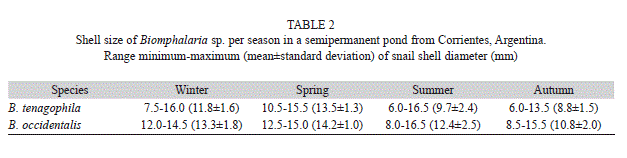
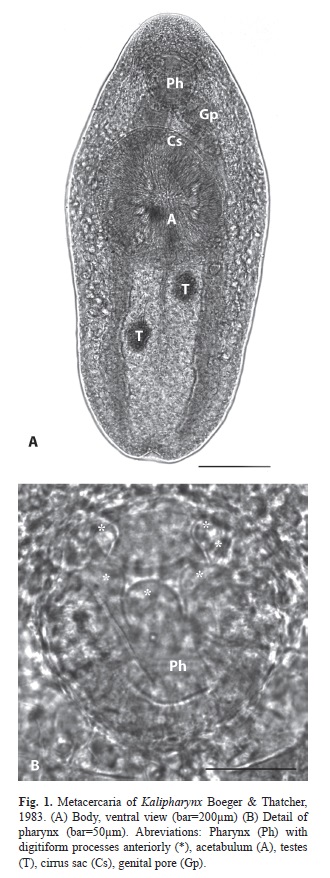
Sampling and laboratory procedure: Samples of B. tenagophila and B. occidentalis were collected between June 2010 and April 2011, encompassing all seasons. Sampling time was one hour and a half for each collecting trip. Snails were always collected by the same two people, along the shoreline of the pond to a maximum depth of 45cm, by using simple meshed of 25cm frame diameter (Thiengo 1995). In laboratory, all snails were measured by maximum shell diameter (mm) with Vernier calipers 0.1mm of precision. Table 2 shows the range of host shell size (mean±SD) of both host species for each sampling season. After that measurements were taken, all snails were inspected and metacercariae encysted were isolated under dissecting microscope from the host. The cysts were manually counted and were measured (diameter) in Carl Zeiss Jena microscope using an ocular micrometer (µm). For each snail infected, the mean cyst diamater of all metacercariae harboured was calculated. After metacercariae were removed from cysts with the use of preparation needles and specimens were observed in vivo mounted in 0.6% saline solution, these were killed in hot distilled water and preserved in 70% ethyl alcohol (Hamann & González, 2009). Metacercariae were stained with alcoholic hydrochloric carmine, dehydrated in a graded ethyl alcohol series, cleared in creosote and mounted in Canada balsam. Systematic determination of metacercariae was carried out following Boeger & Thatcher (1983), based on descriptions about genus Kalipharynx Boeger & Thatcher, 1983. The characteristics considered for systematic determination were pharynx large, strongly muscled and with digitiform processes anteriorly, acetabulum large pre-equatorial, testes diagonal post-acetabular and post-ovarian, cirrus-sac long and genital pore ventro-lateral between suckers (Fig. 1A, B). Specimens of metacercariae were deposited in the Helminthological Collection of Centro de Ecología Aplicada del Litoral (CECOAL), Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Corrientes, Province, Argentina (accession numbers CECOAL 1008180323, CECOAL 1008180325).
Prevalence and mean intensity of infection were calculated according to Bush, Lafferty, Lotz, and Shostak (1997). Comparison of two proportions (Z test) was used to test for differences in prevalence of larval infection between seasons. The intensity of infection and its relation with biotic and abiotic factors were analysed with a multivariate analyses of variance (MANOVA). Data of intensity of infection, host size and cyst size were log10 (x+1) transformed to satisfy the conditions for normality (Sokal & Rohlf, 1995). When the MANOVA Wilks’ Lambda was significant (<0.05), an univariate test (ANOVA) was carried out for each dependent variable; the probabilities were calculated according to Bonferroni procedure, because it provides great control over Type I error (Krebs, 1999). The data of autumn was not considered in MANOVA, because no infection was observed. Spearman´s rank test (rs) was used to examine the relationships between prevalence, intensity of infection and snail size, and between mean intensity of infection and cyst size. For correlations with prevalence, snail sizes were grouped into shell-diameter classes. P values <0.05 were considered significant. The software used was Xlstat 7.5 (Addinsoft, 2004) and SYSTAT 7.0 (SPSS, 1997).
Results
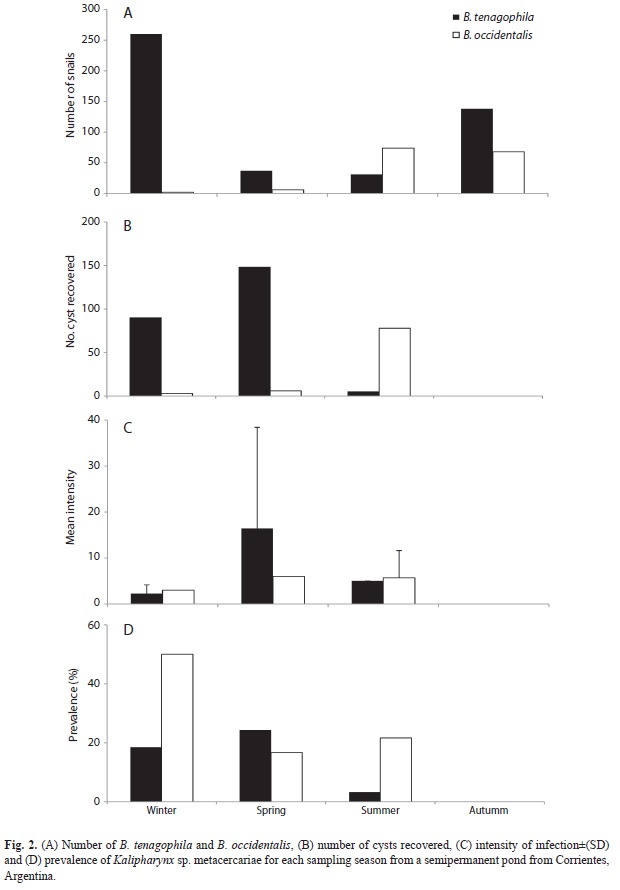
A total of 362 metacercariae (n=262 and n=100, for B. tenagophila and B. occidentalis, respectively) were collected from 616 snails (n=466 and n=150 for B. tenagophila and B. occidentalis, respectively) between June (2010) and April (2011). The metacercarial cysts were found in the digestive gland (7.9% B. tenagophila; 1.0% B. occidentalis), mantle cavity (42.2%; 71.0%), intestine (43.1%; 26.0%) and ovotestis (2.7%; 2.0%). The shell size of snails collected ranged from 6.0 to 16.5mm (mean=10.9±SD=2.2) in B. tenagophila and from 8.0 to 16.5mm (11.8±2.4) in B. occidentalis (Table 2).
Prevalence and mean intensity vs. season: B. tenagophila showed the highest prevalence of infection in spring (Z=2.4, df=1, p<0.05), whereas B. occidentalis showed no significant difference between the prevalence of spring (16.6%) and the prevalence of summer (21.6%) (Z=-0.3, df=1, p>0.05) (Fig. 2D). In this analysis only spring and summer were considered, because of uneven season sample sizes of snails (Fig. 2A). We performed MANOVA by using the season as factor, and infection intensity, cyst size and host shell size as the dependent variables. These results showed that the season had a significant effect on these variables (MANOVA Wilk Lambda=0.8; F3,62=5.1; p=0.003). Therefore, an a posteriori univariate test (ANOVA) was performed (using probabilities according to Bonferroni criteria: α=0.016). These data showed significant differences in the infection intensity (F1,64=7.8; p=0.007) and host shell size (F1,64=8.5; p=0.005) between seasons (Fig. 2C). Cyst size did not show significant seasonal difference (F1,64=2.6; p=0.112).
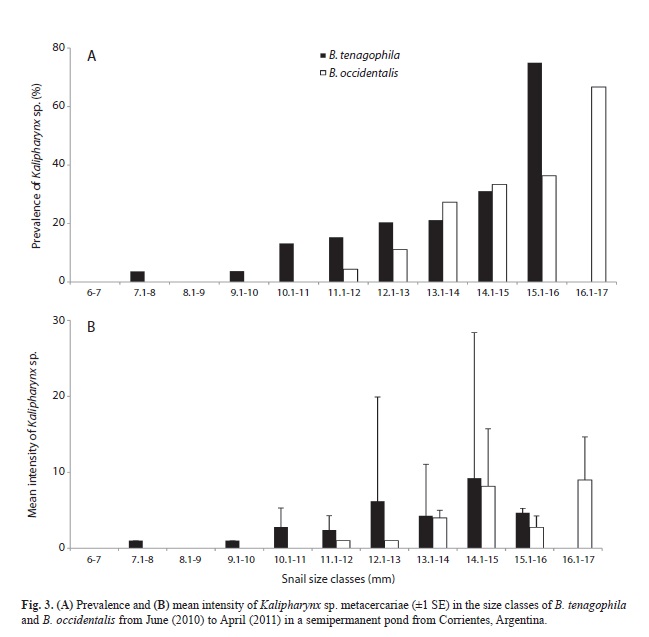
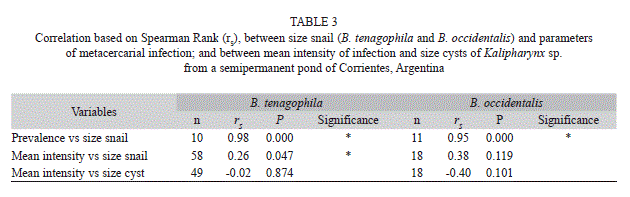
Prevalence and intensity of infection vs size host: The overall prevalence of infection was 12.4% (n=58) and 12.0% (n=18) in B. tenagophila and B. occidentalis, respectively. The smallest size class of B. tenagophila harbouring a metacercarial infection was 7.1-8.0mm and in B. occidentalis was 11.1-12.0mm (Fig. 3A). The maximum prevalence of infection for each species was recorded in snails larger than 15mm, e.g., for B. tenagophila at 15.1-16.0mm size class and B. occidentalis at 16.1-17.0mm size class, with a peak in prevalence of 75.0% and 66.6%, respectively (Fig. 3A). The prevalence of infection was significantly and positively correlated with snail size in both host species (Table 3).
The number of metacercariae encysted ranged from 1 to 60 (mean=4.5±SD=9.9) in B. tenagophila and from 1 to 23 (5.5±5.6) in B. occidentalis. The highest mean intensity of infection occurring in the size range 14.1-15.0mm in B. tenagophila and in the size range 16.1-17.0mm in B. occidentalis (Fig. 3B). The shell size of B. tenagophila was significantly and positively correlated with intensity of infection, whereas the shell size of B. occidentalis showed a non-significant positive correlation with intensity of infection (Table 3).
Intensity of infection vs. size of metacercarial cysts: The cyst diameter ranged from 255 to 705μm (466.4±119.0) in B. tenagophila, and from 310 to 900μm (554.5±150.0) in B. occidentalis. Non-significant negative correlations were observed between intensity of infection and cyst size in both host species (Table 3).
Discussion
Several seasonal studies of trematodes (see review Esch & Fernandez, 1994) showed two peaks in parasite infection caused by the combination of snail mortality, temperature changes and ephemeral presence of allogenetic trematodes. In the present study, although absent in autumn, metacercariae of Kalipharynx sp. were present most of the year in both species of Biomphalaria, showing high values in both warm-season. The drastic drop of infection parameters in autumn, despite the optimal temperature conditions compared with those in other seasons, may be related to the decreased host average size in this season or the extensive host mortality (Esch & Fernandez, 1994). In addition, both infection intensity and host shell size varied significantly between three seasons analyzed (winter-summer), but size of metacercarial cyst did not show seasonal variation, this result suggests that there may be more than one emergence peak of cercariae since the temperature slight fluctuations between seasons may allow the formation and emergence of cercariae more than once in the year (Ostrowski de Núñez, et al., 1990, 1991).
Regarding host size, the prevalence of metacercarial infection increased with a corresponding increase in snail size. Similar results were observed for metacercariae of echinostomatids and strigeids in the freshwater snail B. peregrina (Flores et al., 2010). Also, we found that larger snails harbored more parasites (in terms of intensity of infection) than smaller snails, although this relation was significant only in B. tenagophila, which suggest that host size is an important factor in our system. Several studies (Campbell, 1973; Hamann & Kehr, 1999; Keeney, Walker, Khan, King & Poulin, 2009; Saldanha et al., 2009; Flores et al., 2010) showed that the intensity of metacercarial infection may be conditioned by the intermediate host body size, e.g. the larger size of the host provides more resources (i.e. space and/or availability nutrients) for the growth of the metacercariae. Additionally, Morley et al. (2004) has demonstrated an increase in the mean number of cysts per host with size host (Lymnea peregra) and time of exposure, that is, the larger and older snails harbouring more metacercariae.
On the other hand, we identified a trend to reduce the size of the cysts when the intensity of infection is high. Similarly, other studies have reported reductions in the growth of metacercariae when the intensity of infection increases in the intermediate host (Shandland & Goater, 2000; Brown et al., 2003). Indeed, in their study about intraspecific variation in body size of metacercariae in isopods, Saldanha et al. (2009) showed that the intensity of infection was the determining factor on the variation in size of the metacercariae, showing a weak correlation with host size.
In conclusion, although absent in the autumn, metacercariae of Kalipharynx sp. are present most of the year in both species of Biomphalaria. Also, no seasonal variation of cyst size suggests the possibility of more than one peak of cercariae emergence during the year. Lastly, we detected an increase of prevalence and intensity of metacercarial infection with increasing the snail size, which suggest that larger snails have been exposed to cercariae for a longer period, and a tendency towards density-dependent reductions in the growth of cysts, which could be due to the crowding effect. This is the first study in Argentina analyzing the population biology of Kalipharynx sp. metacercariae.
Acknowledgments
The English language was improved by Inglés Científico (www.inglescientifico.com.ar).
References
Addinsoft. 2004. Xlstat for Excel, version 7. 5. New York, USA: Addinsoft. [ Links ]
Boeger, W. A. & Thatcher, V. E. 1983. Kalipharynx piramboae gen. et sp. n. (Trematoda: Fellodistomidae) parasite do peixe pulmonado amazonico Lepidosiren paradoxa Fitzinger. Acta Amazonica 13: 171-175. [ Links ]
Brown, S. P., de Lorgeril, J., Loly, C. & Thomas, F. 2003. Field evidence for density-dependent effects in the trematode Microphallus papillorpbustus in its manipulated host, Gammarus insensibilis. Journal of Parasitology 89: 668-672. [ Links ]
Bush, A. J., Lafferty, K. D., Lotz, J. M. & Shostak, A. W. 1997. Parasitology meets ecology on its own terms: Margolis et al. revisted. Journal of Parasitology 83: 575-583. [ Links ]
Campbell, R. A. 1973. Influence of temperature, host, and host size on metacercarial development of Cotylurus flabelliformis (Trematoda: Strigeidae). Transactions of the American Microscopical Society 92: 662-665. [ Links ]
Carnevali, R. 1994. Fitogeografía de la provincia de Corrientes. Asunción, Paraguay: Ediciones Litocolor. [ Links ]
Chubb, J. C. 1979. Seasonal occurrence of helminths in freshwater fishes. Part II Trematoda. Advances in Parasitology 17: 141-296. [ Links ]
Esch, G. W. & Fernandez, J. C. 1994. Snail-trematode interactions and parasite community dynamics in aquatic systems: a review. The American Midland Naturalist 131: 209-237. [ Links ]
Esch, G. W., Barger, M. A. & Fellis, K. J. 2002. The transmission of digenetic trematodes: style, elegance, complexity. Integrative and Comparative Biology 42: 304-312. [ Links ]
Flores, V. R., Semenas, L. G. & Veleizán, A. A. 2010. Larval digenean community parasitizing the freshwater snail, Biomphalaria peregrina (Pulmonata: Planorbidae), from a temporary pond in Patagonia, Argentina. Journal of Parasitology 96: 652-656. [ Links ]
Fried, B. & Graczyk, T. K. 1997. Advances in trematode biology. New York, USA: CRC. [ Links ]
Graham, A. L. 2003. Effects of snail size and age on the prevalence and intensity of avian schistosome infection: relating laboratory to field studies. Journal of Parasitology 89: 458-463. [ Links ]
Hamann, M. I. & Kehr, A. I. 1999. Relaciones ecológicas entre metacercarias de Lophosicyadiplotomum sp. (Trematoda, Diplostomidae) y Lysapsus limellus Cope, 1862 (Anura: Pseudidae) en una población local del nordeste argentino. Facena 15: 39-46. [ Links ]
Hamann, M. I. 2004. Seasonal maduration of Catadiscus propinquus (Digenea: Diplodiscidae) in Lysapsus limellus (Anura: Pseudisae) from Argentinean subtropical permanent pond. Physis 59: 29-36. [ Links ]
Hamann, M. I. 2006. Seasonal maturation of Glypthelmins vitellinophilum (Trematoda: Digenea) in Lysapsus limellus (Anura: Pseudisae) from an Argentinian subtropical permantent pond. Physis 59: 29-36. [ Links ]
Hamann, M. I. & C. E. González. 2009. Larval digenetic trematodes in tadpoles of six amphibian species from Northeastern Argentina. Journal of Parasitology 95: 623-628. [ Links ]
Hamann, M. I., A. I. Kehr & González, C. E. 2010. Helminth community structure of Scinax nasicus (Anura: Hylidae) from a South American sobtropical area. Disease of Aquatic Organisms 93: 71-82. [ Links ]
Keeney, D. B., Walker, K. B., Khan, N., King, T. M. & Poulin, R. 2009. The influence of clonal diversity and intensity-dependence on trematode infections in an amphipod. Parasitology 136: 339-348. [ Links ]
Krebs, C. J. 1999. Ecological methodology. Menlo Park, California, USA: Addison Wesley Longman. [ Links ]
Lunaschi, L. I. 1994. Primer registro de Kalipharynx piramboae Boeger y Tatcher, 1983 (Trematoda: Plagiorchiidae) en Argentina. Neotrópica 40: 9-13. [ Links ]
Minchella, D. J., Leathers, B. K., Brown, K. M. & Mcnair, J. N. 1985. Host and parasite counter adaptations: an example from a freswater snail. American Naturalist 126: 843-854. [ Links ]
Morley, N. J., Lewis, J. W. & Adam, M. E. 2004. Metacercarial utilization of a naturally infected single species (Lymnea peregra) snail community by Echinoparyphium recurvatum. Journal of Helmintology 78: 51-56. [ Links ]
Ostrowski de Núñez, M., Hamann, M. I. & Rumi, A. 1990. Larval trematodes of Schistosoma mansoni transmiting snail: Biomphalaria spp. in northeastern Argentina. Acta Parasitologica Polonica 35: 85-96. [ Links ]
Ostrowski de Núñez, M., Hamann, M. I. & Rumi, A. 1991. Population dynamics of planorbid snail from a lenitic biotope in northeastern Argentina. Larval trematodes of Biomphalaria occidentalis and analysis of their prevalence and seasonality. Acta Parasitologica Polonica 36: 159-166. [ Links ]
Poi de Neiff, A. & Neiff, J. J. 2006. Riqueza de especies y similaridad de los invertebrados que viven en plantas flotantes de la planicie de inundación del río Paraná (Argentina). Interciencia 31: 221-225. [ Links ]
Poi de Neiff, A., Galassi, M. E. & Franceschini, M. C. 2009. Invertebrate assemblages associated with leaf litter in three floodplain wetlands of the Paraná river. Wetlands 29: 896-906. [ Links ]
Pojmanska, T., Tkach, V. V. & Gibson, G. I. 2008. Genera incertae sedis, genera inquirenda, nomina nuda, larval or collective names and recently erected genera, p. 735-755. In R. A. Bray, D. I. Gibson & A. Jones. Keys to the Trematoda, Volume 3. London, England: The Natural History Museum and CABI Publishing. [ Links ]
Rumi, A. 1991. La familia Planorbidae Rafinesque, 1815 en la República Argentina. [ Links ]
Rumi, A., Gutiérrez, D. E., Núñez, V. & Darrigran, G. A. 2008. Malacología latinoamericana. Moluscos de agua dulce de Argentina. Revista Biología Tropical 56: 77-111. [ Links ]
Saldanha, I., Leung, T. L. F. & Poulin, R. 2009. Causes of intraespecific variation in body size among trematode metacercariae. Journal of Helminthology 83: 289-293. [ Links ]
Shandland, G. J. & Goater, C. P. 2000. Development and intensity dependence of Ornithodiplostomum ptychocheilus metacercariae in fathead minnows (Pimephales promelas). Journal of Parasitology 86: 1056-1060. [ Links ]
Sokal, R. R. & Rohlf, F. J. 1995. Biometry. New York, USA: WH Freeman and Company. [ Links ]
Sorensen, R. E. & Minchella, D. J. 2001. Snail-trematode life history interactions: past trends and future directions. Parasitology 123: 3-18. [ Links ]
SPSS. 1997. SYSTAT 7.5 for Windows. SPSS Inc. Chicago, USA. [ Links ]
Thiengo S. 1995. Técnicas malacológicas, p. 255. In F. S. Barbosa. Tópicos em malacologia médica. Rio de Janeiro, Brasil: Fio Cruz. [ Links ]
Toledo, R. & B. Fried. 2010. Biomphalaria snails and larval trematodes. New York, USA: Springer. [ Links ]
*Correspondencia a:
1María Virginia Fernández: Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Centro de Ecología Aplicada del Litoral (CECOAL), Ruta Provincial Nº 5Km 2.5, W 3400 AMD, Corrientes, Argentina; mvirginiafernandez@yahoo.com.ar
1Monika Inés Hamann: Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Centro de Ecología Aplicada del Litoral (CECOAL), Ruta Provincial Nº 5Km 2.5, W 3400 AMD, Corrientes, Argentina; monika_hamann@yahoo.com
1Arturo Ignacio Kehr: Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Centro de Ecología Aplicada del Litoral (CECOAL), Ruta Provincial Nº 5Km 2.5, W 3400 AMD, Corrientes, Argentina; arturokehr@yahoo.com.ar
Received 28-IX-2012. Corrected 30-IV-2013. Accepted 28-V-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}