










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.4 San José Oct./Dec. 2013
Dinámica poblacional de Donax denticulatus (Bivalvia: Donacidae) en playa Carenero, costa sur oriental de Cuba
Population dynamic of Donax denticulatus (Bivalvia: Donacidae) at Carenero Beach, Southeastern Cuba
Population dynamic of Donax denticulatus (Bivalvia: Donacidae) at Carenero Beach, Southeastern Cuba
*Dirección para correspondencia:
Abstract
Bivalve molluscs of Donax genus are a very important component of macro-invertebrate assemblages of sandy beaches, and some species are of commercial value in different countries. Although in Cuba Donax denticulatus is not a currently exploited species, the information concerning a stock assessment is a basic step for future use of this resource. With the aim to generate new data on this species structure, growth and secondary production, monthly samples of D. denticulatus were taken from a beach of the Southeastern coast of Cuba, from February to December 2008. Samples were taken from four stations located along the beach; three strata were marked (P1, P2 and P3) per station across the intertidal zone. Three replicated samples of sediments were taken from each stratum with a 0.025m2 PVC core and were posteriorly sieved with a 1mm mesh. Histograms of shell length were constructed based on 1mm intervals and growth parameters were estimated using ELEFAN I routine of FISAT. Mean density ranged from 146.67-855.55ind./m2 and no differences were found among months (ANOVA, p>0.05) but among strata (Scheffé, p<0.05). An association of recruits, young and adults abundances with the strata (X2, p<0.01) was found. Most of the recruits were found in the upper strata while young and adult individuals were abundant in the mid-lower strata. Size frequency histograms of this population showed recruitment between March-June. Growth parameters estimated by Von Bertalanffy were L∞=27.5mm and K=1.5/year; and the life spam was estimated in 1.5 years. Using the length-converted catch curve, the mortality rate (Z) was estimated in 4.97/year. Based in terms of Ash free dry mass (AFDM), mean biomass was estimated in 0.47g/m2.year and the somatic production in 12.4g/m2.year. The renewal rate (P/B) estimated for this population was 26.38, the highest among other Donax populations. High densities, fast growth and high somatic production indicate that this population can be exploited for fishing purposes for which some management measures are proposed.
Key words: Donax, population structure, growth, production, biomass, sandy beaches, Cuba.
Resumen
Con el objetivo de aportar información sobre la estructura poblacional, crecimiento y producción secundaria de una población de D. denticulatus, se tomaron muestras mensuales desde febrero a diciembre 2008, en una playa de la costa sur oriental de Cuba. Las muestras se tomaron con un cilindro de PVC de 0.025m2 en cuatro estaciones ubicadas a lo largo de la playa y en cada estación se delimitaron tres estratos (P1, P2 y P3) en la zona intermareal. No se encontró diferencias en la densidad entre los meses pero sí entre los estratos. Existe asociación en la abundancia de reclutas, jóvenes y adultos entre estratos. Existe un reclutamiento unimodal entre marzo y junio. Los parámetros de crecimiento fueron L∞=27.5mm y K=1.5/año. El tiempo de vida teórico fue de 1.5 años y la tasa de mortalidad (Z) se estimó en 4.97/año. En términos de peso seco libre de cenizas (PSLC) se estimó la biomasa media en 0.47g/m2.año y la producción somática en 12.4g/m2.año, con una tasa de renovación (P/B) de 26.38. La evaluación de la población indica que esta tiene potencial para ser utilizada como carnada o para el consumo.
Palabras clave: Donax, estructura poblacional, crecimiento, producción, biomasa, playas arenosas, Cuba.
Las playas arenosas se consideran como uno de los ecosistemas costeros más dinámicos. Los organismos que habitan las playas arenosas están regulados fundamentalmente por parámetros físicos como el oleaje, las mareas, la pendiente de la playa y el tamaño de los sedimentos (Defeo & McLachlan, 2005). Entre los invertebrados dominantes en este ambiente, se encuentran los bivalvos del género Donax que están distribuidos en todos los continentes (Ansell, 1983). En el Caribe se encuentran D. striatus Linné, 1767 y D. denticulatus Linné, 1758, las cuales comparten el mismo ámbito de distribución, llegando en ocasiones a encontrarse poblaciones de las dos especies en la misma playa (Wade, 1967).
Los primeros estudios sobre D. denticulatus fueron realizados por Wade (1967, 1968) en playas de Jamaica, aportando información sobre la ecología de la especie y su historia natural. Se han registrado datos sobre su densidad en playas de Puerto Rico (Sastre, 1984) y de las Antillas Menores: Nevis, San Vicente, Las Granadinas, Antigua y Trinidad (Wade, 1967). En playas de Venezuela se han realizado estudios sobre el crecimiento y la reproducción de esta especie (Vélez et al. 1985: García et al., 2003: Marcano et al., 2003). Esta especie habita exclusivamente en la zona intermareal de las playas arenosas y el éxito de su adaptación a este ambiente tan dinámico se atribuye a que es un activo migrador mareal capaz de coordinar rápidamente los movimientos para enterrarse en el sedimento (Trueman, 1971). Las poblaciones de D. denticulatus pueden variar en densidad, talla de los individuos y patrones de coloración entre diferentes playas (Wade, 1967). Esta especie presenta un estadio larval planctónico que puede durar entre tres o cuatro semanas antes de asentarse en el sedimento, teniendo altas tasa de mortalidad durante los primeros meses de vida. También existen diferencias en las tasas de crecimiento entre las poblaciones y esto puede estar relacionado con la disponibilidad de alimento (Wade, 1968).
En Cuba esta especie solo se ha reportado para la plataforma occidental (Espinosa, 2007), sin embargo también ha sido observada en playas de la región oriental del país. Hasta el momento solo existe un estudio de D. denticulatus en Cuba y solo ha tratado sobre los caracteres de su forma y tamaño (Ocaña & Fernández, 2011). El presente trabajo tiene como objetivo caracterizar la estructura poblacional, estimar los parámetros de crecimiento y la producción secundaria de D. denticulatus en playa Carenero en la costa sur oriental de Cuba.
Materiales y Métodos
Área de estudio: Playa Carenero se encuentra ubicada hacia el sureste del Golfo de Guacanayabo en los 20°05’07” N - 77°34’32” W en la plataforma sur oriental de Cuba. Presenta una longitud aproximada de 900m con un ancho promedio de 5m en la cara de la playa. Los sedimentos son mixtos compuestos por restos calcáreos de animales marinos y material terrígeno con una granulometría media. La playa presenta una baja pendiente, siendo muy somera con una profundidad media de 0.6m en los primeros 100m de distancia desde la línea de costa. En esta zona el régimen de marea es semidiurno con un rango que no sobrepasa 0.5m. Durante la época de lluvias (mayo-noviembre), la playa recibe la influencia del río Sevilla ubicado a 5km al norte de la playa y de la Ciénaga de Managuano, ubicada detrás de la playa.
Muestreo: Los muestreos se realizaron mensualmente desde febrero hasta diciembre de 2008 durante la marea baja. Se siguió un diseño de muestreo estratificado (Schoeman et al., 2003). Cuatro estaciones fijas separadas por 200m de distancia, fueron seleccionadas a lo largo de la playa. En cada estación se seleccionaron tres estratos (P1, P2 y P3) perpendiculares a la costa coincidiendo con el límite de marea baja, la parte media de la zona intermareal y el límite superior de la resaca, respectivamente. En cada estrato de cada estación, se extrajeron tres réplicas con un cilindro de PVC de 0.025m2 (diámetro=18cm) (Unidad de Muestreo, UM), introduciéndolo en el sustrato arenoso hasta una profundidad de 20cm para un total de 36 unidades de muestreo por mes. El sedimento retenido en el cilindro fue cribado en un tamiz de 1mm de abertura de malla. Los individuos de D. denticulatus se introdujeron en bolsas plásticas y se congelaron.
En el laboratorio, se determinó la longitud anteroposterior de cada individuo (L) con el empleo de un vernier digital, con una precisión de 0.01mm. Los individuos fueron agrupados en intervalos de clases de 1mm para elaborar los histogramas de distribución de tallas en cada mes. Los especímenes también se categorizaron en tres grupos: reclutas (L<5mm), jóvenes (5mm<L<15mm) y adultos (L>15mm), siguiendo los criterios de Wade (1968). A 117 individuos de todas las clases de tallas muestreadas, se les determinó el peso total con concha (PT, g) y luego se extrajo la masa visceral de cada uno, se colocó en papel secante y se determinó el peso de la carne húmeda (PCH, g) en una balanza con resolución de 1mg.
Crecimiento, longevidad y mortalidad: La información básica para la estimación del crecimiento fue obtenida de una serie de 11 histogramas de frecuencias de distribución de tallas. A través del empleo de la rutina ELEFAN I del paquete FISAT II (Gayanilo et al., 2005) se estimaron los parámetros de la ecuación de crecimiento de Von Bertalanffy (ECVB) expresada como: Lt=L∞[1–e(-K(t-to)]; donde Lt es la talla correspondiente a la edad t, L∞ es la longitud máxima teórica, K es el coeficiente de crecimiento y to representa la edad teórica a la longitud cero. Con la técnica de Gulland & Holt (1959) se obtuvieron estimados preliminares de L∞ y K. Usando los valores obtenidos por esta técnica, se empleó el análisis de superficie de respuesta del ELEFAN I para determinar la ECBV que mejor se ajusta a los datos de distribución de tallas. Se seleccionó el mayor valor de Rn como criterio de ajuste.
Para comparar los parámetros de crecimiento con otras especies de Donax se calculó el índice de rendimiento en crecimiento Φ’ (Pauly & Munro 1984): Φ’=2log(L∞)+log(K) y la tasa de crecimiento en el punto de inflexión (Pc) de la curva talla-peso (Pauly, 1979): Pc=log[K(L∞)3]
El tiempo de vida (tmax) teórico para la población de D. denticulatus en playa Carenero fue estimado mediante la inversa de la ecuación de crecimiento de Von Bertalanffy (Taylor, 1958): tmax=[lnL95%-ln(L∞-L95%)]/K; donde L95% representa el 95% de la longitud antero-posterior máxima registrada durante los muestreos.
Con los datos acumulados de los muestreos se elaboró la curva de captura, con el objetivo de hallar la tasa de mortalidad (Z) del modelo de extinción exponencial: Nt=Nt-1(e–Z*Δt); donde Nt es el número de individuos en el tiempo t y Z es el parámetro de curvatura de la ecuación.
Biomasa y producción: La relación talla-peso usando PT(g) y PCH(g), se determinó mediante análisis de regresión lineal a través de la ecuación: ln(PT, PCH)=ln(a)*ln(L)b; donde a y b son parámetros de la curva.
A partir de los datos de densidad, frecuencia de talla, los parámetros de la ECVB y las relaciones talla-peso, se estimó la producción de la población P (g/m2.año), considerando PT y PCH, a través del método de la tasa de crecimiento específico en peso (Crisp, 1984). De acuerdo a este método la expresión matemática para estimar la producción es: P=ΣNi*Mi*Gi (g/m2.año); donde Ni es la densidad media (ind./m2), Mi es el peso medio individual en la clase de talla i y Gi es la tasa de crecimiento específico: Gi=b*K*((L∞/Li)-1) [1/año]; donde b es el exponente de la relación talla-peso y Li es la longitud media de los individuos en la clase de talla i.
La biomasa media anual (B, g/m2.año) se calculó a través de B=ΣNi*Mi y la tasa de renovación anual (P/B) a partir de la producción anual y de la biomasa media anual obtenida.
Para que estos resultados sean comparables con otros estudios en poblaciones del género Donax, los datos de producción y tasa de renovación anual fueron convertidos a peso seco libre de ceniza (PSLC), usando el factor de conversión PSLC/PCH=5.8% (Ricciardi & Bourget, 1998).
Mensualmente se determinó la densidad (ind./m2, media±error estándar) extrapolando el número de individuos obtenidos en cada una de las 36 UM. Para comparar los valores de densidad entre los meses, así como entre los estratos, se empleó un ANOVA unifactorial realizando la transformación log(x+1) de los datos originales para cumplir los requisitos de homocedasticidad. Una vez transformados los datos se corroboró que la variable siguió una distribución normal a través de una prueba de Kolmogorov-Smirnov. Cuando existieron diferencias significativas, los resultados del ANOVA fueron seguidos por una prueba de Scheffé para realizar comparaciones a posteriori. Para evaluar si existe asociación entre las clases de tallas y los estratos, se aplicó una prueba Chi-cuadrado (X2). Las diferencias estadísticas fueron consideradas a un nivel de significación de 95% (α=0.05) (Zar, 1999).
Resultados
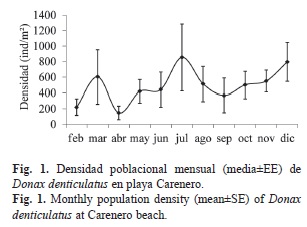
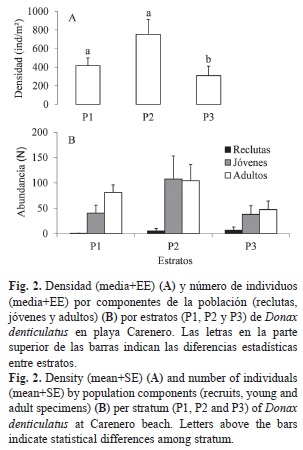
Estructura poblacional: La población de D. denticulatus en playa Carenero no presenta diferencias significativas en la densidad entre meses (ANOVA, p>0.05) (Fig. 1). La mayor abundancia se encontró en julio (855.55±430.04ind./m2), mientras que la menor fue en abril (146.67±87.75ind./m2). Al comparar la densidad entre estratos (Fig. 2A), se observa que en el estrato medio (P2) se alcanzan los mayores valores (753.33±160.0ind./m2) y los menores valores en el estrato superior (P3) (308.79±101.9ind./m2). Existen diferencias significativas de la densidad entre el estrato superior de la playa y los dos más bajos (P1 y P2) (Scheffé, p<0.05). Existe una asociación muy significativa (X2=206.8; g.l=4; p<0.01) entre la composición por tallas y la distribución de los individuos en los diferentes estratos de la playa. La mayor proporción de reclutas se encontró en P3, mientras que la mayor proporción de jóvenes y adultos fue en P2. En P1 se encontró una mayor proporción de adultos con respecto a los jóvenes y reclutas, estos últimos aparecieron con una abundancia muy baja (Fig. 2B).
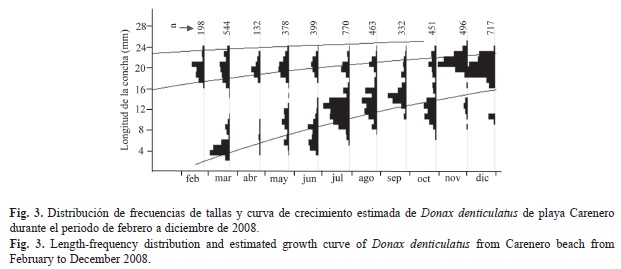
La distribución de frecuencia de tallas (Fig. 3) muestra que la población está compuesta por distintas cohortes. Al principio de los muestreos en febrero existían dos cohortes, una de las cuales desapareció en abril y la otra se mantuvo hasta junio. En marzo y junio aparecieron dos nuevas cohortes que fueron seguidas hasta el final de los muestreos. En octubre se observa una nueva cohorte que debe haber sido reclutada en septiembre pero en este mes no se registró en el muestreo.
Crecimiento, longevidad y mortalidad: Fueron recolectados un total de 4 880 individuos durante todo el periodo de estudio. La menor talla registrada fue de 3.66mm y la longitud máxima alcanzada fue de 25.35mm. Con el empleo del método de Gulland & Holt (1959) se estimaron valores de L∞=27.5mm y K=1.5/año, los cuales se usaron en la rutina ELEFAN I. A través de la rutina ELEFAN I se corroboró que los estimados preliminares presentaron un mayor valor de ajuste (Rn=0.15), lo cual brinda un estimado final de L∞=27.5mm y K=1.5/año. Estos estimados fueron seleccionados para elaborar el modelo de crecimiento de la población (Fig. 3).
Los índices de crecimiento Φ’ y Pc tuvieron valores de 3.05 y 4.49, respectivamente. El tiempo de vida (tmax) teórico fue de 1.5 años. La curva de captura (Fig. 4), permitió estimar la tasa de mortalidad total de esta población en 4.97/año, siendo la ecuación de regresión: ln(N/Δt)=15.82-4.97t (r2=0.93).
Biomasa y producción: La relación observada entre L y PT y entre L y PCH para esta población, quedó expresada respectivamente por las ecuaciones: ln(PT)=(-9.16)*ln(L)3.20 (r2=0.99) y ln(PCH)=(-11.46)*ln(L)3.41 (r2=0.93). Teniendo en cuenta el peso total de los individuos con la concha, la biomasa media alcanzó un valor de 43.0g/m2.año y la producción somática es de 1 081.8g/m2.año. Considerando el peso de la carne húmeda, la biomasa media alcanzó un valor de 8.2g/m2.año y la producción somática es de 213.8g/m2.año. En términos de peso seco libre de cenizas, según el factor de conversión PSLC/PCH=5.8%, la biomasa media alcanzó un valor de 0.47g/m2.año y la producción somática es de 12.4g/m2.año. Aplicando el factor de conversión de Ricciardi & Bourget (1998), la tasa de renovación (P/B) fue de 26.38.
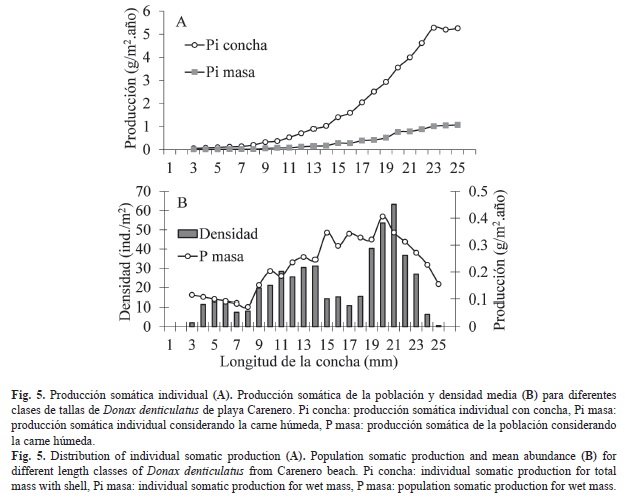
La producción individual (Pi) muestra que el valor más elevado con concha lo reportan los individuos de 23mm-25mm con valores superiores a 5.0g/m2.año. Los individuos de estas mismas clases de tallas también muestran la mayor producción en carne húmeda con valores superiores a 1.0g/m2.año (Fig. 5A). La distribución de la producción anual total en masa húmeda y de la densidad por clases de talla indica que la mayor densidad es de individuos de la talla de 21mm, mientras que la mayor producción fue aportada por individuos de 20mm de longitud (Fig. 5B).
Discusión
Estructura poblacional: La densidad no mostró diferencias significativas entre los meses, contrario a lo que sucede con muchas poblaciones de D. striatus y D. denticulatus en el Caribe donde se han observado variaciones cuando se comparan diferentes localidades e incluso dentro de una misma localidad en diferentes épocas del año (Sastre, 1984: McLachlan et al., 1996: Ocaña et al., 2010). Aunque estadísticamente no aparecen diferencias en la densidad, sí se observa un aumento de esta variable en marzo, julio y diciembre. Esto puede ser explicado por la aparición de nuevos reclutas y a errores asociados al muestreo. En marzo apareció una nueva cohorte que contribuyó con el 45% de los individuos colectados ese mes. Por otra parte, los perfiles de playa muestran condiciones variables que pueden resultar en drásticas contracciones y expansiones de la distribución transversal de la macrofauna. Esto implica que el uso de unidades de muestreo individuales para determinar la densidad no es un método adecuado para hacer estimados de abundancia. En especies intermareales, las agregaciones persisten en el tiempo pero la posición relativa de estas en el eje transversal de la playa puede variar de acuerdo a la susceptibilidad de cada especie a las variaciones en las condiciones ambientales (Brazeiro & Defeo, 1996). Las estrategias de muestreo basadas en gradientes ambientales seleccionando estratos fijos definidos a priori, no cubren toda la distribución transversal de las especies. Estos diseños deben ser evitados en estudios poblacionales en playas arenosas porque brindan un estimado sesgado de la abundancia, sobre todo en playas con pequeños rangos de marea (Defeo & Rueda, 2002).
Los valores de densidad encontrados en playa Carenero fueron superiores a los reportados por García et al. (2003) para playa Brava en Venezuela. En otra playa de Venezuela se reportaron valores de 212-2 435ind./m2 (Vélez et al., 1985). En Puerto Rico se encontró diferencias de la densidad de D. denticulatus entre playas (Sastre, 1984), al igual que en Jamaica (Wade 1967, 1968). Estos autores plantean que estas variaciones se relacionan con el tamaño del sedimento, la pendiente de la playa, el grado de exposición a las olas y el contenido de materia orgánica de la arena.
Los histogramas de composición de frecuencias de tallas muestran que en esta población el reclutamiento ocurre principalmente entre marzo-junio, coincidiendo con un periodo seco según datos aportados por la Estación Meteorológica de Cabo Cruz (ubicada a 30km al sur de playa Carenero). En esta especie otros autores han encontrado un reclutamiento continuo a lo largo del año (Wade, 1968; Vélez et al., 1985; Marcano et al. 2003), sin embargo en playa Carenero después del comienzo de las lluvias a partir de julio, solo se detectó una nueva cohorte (en octubre) pero con una baja abundancia de individuos. Es posible que la incidencia de las lluvias haya provocado una menor supervivencia de las larvas. En playas de Jamaica se ha documentado que la disminución de la salinidad provoca mayores mortalidades de las larvas (Wade, 1968). Sin embargo, los factores que determinan la extensión del reclutamiento en una escala temporal, son aún controversiales.
La población estudiada en playa Carenero muestra un patrón de distribución vertical donde la mayor densidad de individuos se encuentra en los dos primeros estratos, alcanzando el valor máximo en el estrato intermedio. Las poblaciones del género Donax manifiestan patrones de zonación correspondientes con las diferentes clases de tallas. Los juveniles tienden a asentarse en diferentes niveles antes de migrar hacia la zona de los adultos (McLachlan et al., 1996). En playa Carenero los adultos de D. denticulatus se ubican preferentemente en la parte baja de la zona intermareal y los reclutas se disponen en los estratos más altos. Este comportamiento puede estar asociado al tamaño del sedimento. En los muestreos fue evidente que durante el cribado del sedimento, la retención de partículas en el tamiz fue disminuyendo desde P1 hacia P3, siendo muy baja en el estrato superior donde se asientan la mayoría de los reclutas. Esta especie se entierra en el sedimento y al parecer la selección preferencial de los reclutas y jóvenes por los estratos donde el sedimento es más fino, es una forma de evitar ser barridos por el oleaje lo que posibilita que aprovechen más energía en procesos de desarrollo que de mantenimiento. Luego al ir creciendo, los individuos son capaces de migrar hacia la parte más baja de la zona intermareal donde son capaces de soportar el embate del oleaje debido a su mayor capacidad de enterramiento. Este patrón de zonación en la zona intermareal ha sido observado para esta especie en otras playas (Wade, 1967; Marcano et al., 2003) y para otras especies del género como D. striatus (Ocaña et al., 2010), D. faba (Alagarswami, 1966), D. trunculus y D. vittatus (Ansell & Lagardére, 1980; Zeichen et al., 2002) y en D. hanleyanus (Gil & Thomé, 2000).
Crecimiento, longevidad y mortalidad: La máxima longitud alcanzada en esta población es mayor que la reportada en playas de Jamaica por Wade (1967) y en una playa de Venezuela (Vélez et al., 1985), en ambos estudios los individuos no sobrepasaron los 24mm de longitud anteroposterior. En otras playas de Venezuela los individuos alcanzaron tallas por encima de los 30mm (García et al., 2003; Marcano et al., 2003). En playas de Puerto Rico los individuos alcanzaron tallas entre 20mm-30mm (Sastre, 1984), esta variación en el crecimiento fue atribuida a factores como las características morfodinámicas de las playas y al nivel de materia orgánica contenido en los sedimentos.
El valor de K estimado es mayor que el valor reportado por Vélez et al. (1985), similar al obtenido por García et al. (2003) y menor que el valor estimado por Marcano et al. (2003), todos en playas de Venezuela. La tasa de crecimiento y el índice de rendimiento en crecimiento, calculados para esta población, son relativamente altos. Herrmann et al. (2009) realizaron una compilación para comparar el crecimiento entre 29 especies de Donax en diferentes áreas climáticas. Según esos datos la población de D. denticulatus de playa Carenero, presenta una tasa de crecimiento similar a las poblaciones estudiadas en Venezuela. Los altos valores de la tasa de crecimiento también coinciden con los de otras especies localizadas en zonas templadas que presentan mayores tallas, lo cual puede explicarse por el hecho que los estimados de crecimiento basados sólo en datos de frecuencia de longitud pueden ser sesgados para las especies tropicales. Esto se debe a que las especies tropicales presentan un reclutamiento continuo y por tanto la composición de tallas puede no presentar modas definidas que permitan separar las distintas cohortes (Sparre & Venema, 1997). Este resultado también podría estar relacionado con errores del diseño de muestreo donde la selección a priori de estratos conllevó a que no se detectaran los individuos de una cohorte en algunos meses y en otros sí. En los histogramas de frecuencia de tallas se hace evidente este error al observar que la segunda cohorte presenta mayor número de individuos en noviembre y diciembre con respecto a los meses precedentes.
La mayoría de las especies de Donax presentan una longevidad de uno a dos años, mientras la mortalidad es a menudo alta, particularmente en las especies tropicales (Ansell, 1983). El valor tmax=1.5 años, estimado en este estudio, es similar a los valores de otras poblaciones de la misma especie o de su congénere D. striatus (Wade, 1968: Vélez et al., 1985: McLachlan et al., 1996: Rocha-Barreira et al., 2002: Marcano et al., 2003), que comparte el mismo ámbito de distribución. La tasa de mortalidad obtenida para D. denticulatus es superior a otras especies tropicales como D. incarnatus (Thippeswamy & Joseph, 1992), D. dentifer (Riascos & Urban, 2002) y D. obesulus (Aguirre & Mendo, 2008). Este hecho también pudiera estar asociado a errores en el muestreo como se explicó anteriormente. La población de playa Carenero no es objeto de explotación pesquera por lo que se asume que la mortalidad observada (Z) se corresponde con la mortalidad natural.
Biomasa y producción: La biomasa media alcanzó un valor de 0.47g/m2.año, lo que se corresponde con valores obtenidos para especies que habitan en las regiones tropicales y subtropicales (Herrmann et al., 2009). Sin embargo, la producción somática alcanzó un valor de 12.4g/m2.año, que es superior al resto de las especies de Donax que habitan en estas áreas climáticas. Estos valores pudieran estar sobre estimados debido a que inicialmente los individuos colectados fueron congelados y durante el proceso de secado en papel (ver materiales y métodos) pudo haber quedado retenido líquido contribuyendo a obtener mayores valores en el peso. Este error pudo haber sido amplificado al emplear el factor de Ricciardi & Bourget (1998) para convertir los valores a PSLC.
La población de D. denticulatus de playa Carenero presentó una tasa de renovación anual superior a la reportada para cualquier otra especie del mismo género. Independientemente de haber error asociado al muestreo y a las estimaciones de los parámetros como biomasa y producción somática, es de esperar un alto valor de P/B. Los altos valores de P/B son característicos de poblaciones compuestas por individuos de pequeñas tallas que crecen muy rápido y tienen poca longevidad. La alta producción puede favorecer que la población se recupere rápidamente de la alta tasa de mortalidad natural y por tanto sea un mecanismo compensatorio que permita mantener estable la abundancia de la población.
La población de D. denticulatus de playa Carenero tiene potencialidades para su uso como carnada o para el consumo humano, debido a un rápido crecimiento y su alta tasa de renovación anual. En este sentido se sugiere que en caso de explotarse, esta actividad se realice en los estratos más bajos de la playa donde se localizan los individuos más grandes; a fin de evitar el impacto sobre los individuos más pequeños que se localizan en la parte superior de la zona intermareal. Otro aspecto importante sería tener en cuenta que el reclutamiento ocurre entre marzo-junio, por tanto las capturas se deben realizar preferentemente en otra época del año. Cabe destacar que debe tenerse en cuenta también que la población tiene una alta tasa de mortalidad natural, por tanto es necesario llevar un control sobre el esfuerzo pesquero. Se recomienda que otros estudios que se realicen sobre especies de almejas de playas en el Caribe tengan en cuenta un diseño de muestreo con mayor número de réplicas de las unidades de muestreo ocupando todo el eje transversal de la playa a fin de evitar el sesgo en la estimación de la abundancia, del crecimiento, la producción somática y la mortalidad. También se recomienda realizar la determinación directa de PSCL y emplear esta medida en la relación talla-peso en lugar de PCH que metodológicamente introduce error en la estimación de la biomasa y la producción somática de la población.
Agradecimientos
Agradecemos el apoyo brindado por Vladimir Martínez, Carlos Ocano y Osbel Calaña durante los muestreos de campo. A Liliana Gómez que, desde un inicio nos apoyó en el laboratorio. Los comentarios de Alberto de Jesús Navarrete y cuatro revisores anónimos ayudaron a mejorar el manuscrito.
Referencias
Aguirre, A., & Mendo, J. 2008. Crecimiento y producción de Donax obesulus Reeve, 1854 (Bivalvia: Donacidae) en playa Sarapampa, Asia, Lima. Ecologia Aplicada, 7, 63-70. [ Links ]
Alagarswami, K. 1966. Studies on some aspects of biology of the wedge-clam Donax faba from Mandapam coast in the Gulf of Mannar. Journal of Marine Biology Association India, 8, 56-75. [ Links ]
Ansell, A. D. 1983. The biology of the genus Donax. In A. McLachlan., T. Erasmus & W. Junk (Eds.). Developments in hydrobiology, 19. Sandy beaches as ecosystems. (pp. 607-635). La Haya, HG: W. Junk. [ Links ]
Ansell, A. D., & Lagardére, F. 1980. Observations on the biology of Donax trunculus and Donax vittatus at Ile d’Oléron (French Atlantic coast). Marine Biology, 57, 287-300. [ Links ]
Brazeiro, A., & Defeo, O. 1996. Macroinfauna zonation in microtidal sandy beaches: is it possible to identify patterns in such variable environments? Estuarine, Coastal and Shelf Sciences, 42, 523-536. [ Links ]
Crisp, D. J. 1984. Energy flow measurements, p: 284-372. In N.A. Holme & A.D. McIntyre (Eds.). Methods for the study of marine benthos. IBP Handbook 16. Oxford, OX: Blackwell Scientific. [ Links ]
Defeo, O., & McLachlan, A. 2005. Patterns, processes and regulatory mechanisms in sandy beach macrofauna: a multi-scale analysis. Marine Ecology Progress Series, 295, 1-20. [ Links ]
Defeo, O., & Rueda, M. 2002. Spatial structure, sampling design and abundance estimates in sandy beach macroinfauna: some warnings and new perspectives. Marine Biology, 140, 1215-1225. [ Links ]
Espinosa, J. 2007. Moluscos–Filo MOLLUSCA. In R. Claro (Ed.). La Biodiversidad marina de Cuba. (CD-ROM) (p. 57-62), Instituto de Oceanología, Ministerio de Ciencia, Tecnología y Medio Ambiente, La Habana, Cuba. [ Links ]
García, N., Prieto, A., Alzola, R., & Lodeiros, C. 2003. Crecimiento y distribución de tallas de Donax denticulatus (Mollusca: Donacidae) en playa Brava, península de Araya, estado Sucre, Venezuela. Revista Cientifica FCV-LUZ, 13, 464-470. [ Links ]
Gayanilo, F. C. Jr., Sparre, P., & Pauly, D. 2005. FAO-ICLARM stock assessment tools II (FiSAT II). User’s guide, FAO, 8. Roma, Italia. [ Links ]
Gil, G. M., & Thomé, J. W. 2000. Morfometria da concha em Donax hanleyanus Philippi, 1847 (Mollusca, Bivalvia, Donacidae) e sua relação com as zonas de praia. Acta Biologica Leopoldensia, 22, 161-170. [ Links ]
Gulland, J. A., &. Holt, S. J. 1959. Estimation of growth parameters for data at inequal time intervals. Journal du Conseil / Conseil Permanent International pour l’Exploration de la Mer, 25, 47-49. [ Links ]
Herrmann, M., Carstensen, D., Fischer, S., Laudien, J., Penchaszadeh, P. E., & Arntz, W. E. 2009. Population structure, growth and production of the wedge clam Donax hanleyanus (Bivalvia: Donacidae) from northern Argentinean beaches. Journal of Shellfish Research, 28, 511-526. [ Links ]
Marcano, J. S., Prieto, A., Lárez, A., & Salazar, H. 2003. Crecimiento de Donax denticulatus (Linné 1758) (Bivalvia: Donacidae) en la ensenada La Guardia, isla de Margarita, Venezuela. Zootecnia Tropical, 21, 237-259. [ Links ]
McLachlan, A., Dugan, J. E., Defeo, O., Ansell, A. D., Hubbard, D. M., Jaramillo, E., & Penchaszadeh, P. E. 1996. Beach clam fisheries. Oceanography and Marine Biology: An* Annual Reviews, 34, 163-232. [ Links ]
Ocaña, F. A., & Fernández, A. 2011. Morfometría de la concha de Donax denticulatus y Donax striatus de dos playas de Cuba oriental. Revista de Ciencias Marinas y Costeras, 3, 67-75. [ Links ]
Ocaña, F. A., Fernández, A., Silva, A., González, P. A., & García, Y. 2010. Estructura poblacional de Donax striatus (Bivalvia, Donacidae) en playa Las Balsas, Gibara, Cuba. Revista de Ciencias Marinas y Costeras, 2, 27-38. [ Links ]
Pauly, D. 1979. Gill size and temperature as governing factors in fish growth: a generalization of von Bertalanffy’s growth formula. Berlin Institut für Meereskunde an der Universität Kiel 63, 156p. [ Links ]
Pauly, D., & Munro, J. L. 1984. Once more on the comparison of growth in fish and invertebrates. Fishbyte, 2, 21. [ Links ]
Riascos, J. M., & Urban, H. J. 2002. Dinámica poblacional de Donax dentifer (Veneroida: Donacidae) en Bahía Málaga, pacífico colombiano durante el fenómeno “El Niño” 1997/1998. Revista de Biologia Tropical, 50, 1113-1123. [ Links ]
Ricciardi, A., & Bourget, T. E. 1998. Weight-to-weight conversion factors for marine benthic macroinvertebrates. Marine Ecology Progress Series, 163, 245-251. [ Links ]
Rocha-Barreira, C. A., Batista, W. F., de Oliveira Monteiro, D., & Franklin-Junior, W. 2002. Aspectos da estrutura populacional de Donax striatus (Linnaeus, 1758) (Mollusca: Donacidae) na Praia do Futuro, Fortaleza-CE. Arquivos de Ciências do Mar, 35, 51-55. [ Links ]
Sastre, M. P. 1984. Relationships between environmental factors and Donax denticulatus populations in Puerto Rico. Estuarine, Coastal and Shelf Science, 19, 217-230. [ Links ]
Schoeman, D. S., Wheeler, M., & Wait, M. 2003. The relative accuracy of standard estimators for macrofaunal abundance and species richness derived from selected intertidal transect designs used to sample exposed sandy beaches. Estuarine, Coastal and Shelf Science, 58S, 5-16. [ Links ]
Sparre, P., & Venema, S. C. 1997. Introducción a la Evaluación de Recursos Pesqueros Tropicales. Parte 1. Manual. FAO Documento Técnico de Pesca 306/1 Rev. 2. Roma, Italia. [ Links ]
Taylor, C. C. 1958. Cod growth and temperature. Journal du Conseil / Conseil Permanent International pour l’Exploration de la Mer, 23, 366-370. [ Links ]
Thippeswamy, S., & Joseph, M. M. 1992. Allometry in the wedge clam, Donax incarnatus (Gmelin) from Panambur beach, Mangalore. Indian Journal of Marine Sciences, 21, 161-163. [ Links ]
Trueman, R. D. 1971. The control of burrowing and the migratory behavior of Donax denticulatus (Bivalvia: Tellinacea). Journal of Zoology, 165, 453-469. [ Links ]
Vélez, A., Venables, B. J., & Fitzpatrik, L. C. 1985. Growth production of the tropical beach clam Donax denticulatus (Tellinidae) in eastern Venezuela. Caribbean Journal of Science, 21, 63-73. [ Links ]
Wade, B. A. 1967. On taxonomy, morphology, and ecology of the beach clam, Donax striatus Linné. Bulletin of Marine Science, 17, 723-740. [ Links ]
Wade, B. A. 1968. Studies on the biology of the West Indian beach clam, Donax denticulatus L. 2. Life-history. Bulletin of Marine Science, 18, 876-901. [ Links ]
Zar, J. H. 1999. Biostatistical Analysis (4th. ed.). New Jersey, N.J.: Prentice-Hall. [ Links ]
Zeichen, M. M., Agnesi, S., Mariani, A., Maccaroni, A., & Ardizzone, G. D. 2002. Biology and population dynamics of Donax trunculus L. (Bivalvia: Donacidae) in the South Adriatic coast (Italy). Estuarine, Coastal and Shelf Science, 54, 971-982. [ Links ]
*Correspondencia a:
1 Frank A. Ocaña. Centro de Investigaciones y Servicios Ambientales y Tecnológicos, Calle 18 s/n esq. a Maceo, Rpto. “El Llano”, Holguín, Cuba.
2 Yanet C. Apín. Centro de Estudios Multidisciplinarios de Zonas Costeras, Universidad de Oriente, Ave. Las Américas, Santiago de Cuba, Cuba; yanet@cemzoc.uo.edu.cu
3 Programa de doctorado en ecología y desarrollo sustentable. El Colegio de la Frontera Sur, Unidad Chetumal, Q. Roo, México; frankocisat@gmail.com
3Yuself R. Cala. Programa de doctorado en ecología y desarrollo sustentable. El Colegio de la Frontera Sur, Unidad Chetumal, Q. Roo, México; sgigas21824@gmail.com
Recibido 27-III-2012. Corregido 10-II-2013. Aceptado 13-III-2013


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}