










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.4 San José Oct./Dec. 2013
Diversidad de aves en el Centro Agropecuario Cotové, Santa Fe de Antioquia, Colombia
Diversity of birds in Cotové Agricultural Center, Santa Fe de Antioquia, Colombia
Diversity of birds in Cotové Agricultural Center, Santa Fe de Antioquia, Colombia
*Dirección para correspondencia:
Abstract
Fragmentation of natural habitats is a central concern of biodiversity conservation and is considered a significant factor contributing to species loss and populations decline. We studied this in Cotové Agricultural Center in the municipality of Santa Fe de Antioquia (Colombia),where the conversion of natural forests to different land use systems, has limited the amount of available habitat. With the aim to describe the effect of habitat loss on bird presence in five land uses (fruiting trees, silvopastoral systems, secondary forest, pasture low density of trees and grass cutting) in this area, we studied bird diversity using two methods: fixed point counts and mist nets to analyze the influence of land use in the composition, richness and abundance of birds present were studied. A total of 6 633 individuals of 101 species were observed of which 11 species were migratory. The insectivorous and frugivorous foraging guilds were better represented. The Shannon index indicated the highest values of alpha diversity for the silvopastoral system, and the fruit with the lowest. The grazing and silvopastoral land systems shared more species with low tree density. The importance of the environmental heterogeneity found is highlighted as a positive factor for bird species richness, mostly of low and middle habitat specificity, and preferably from edge habitats and open areas. In order to preserve natural regeneration and connectivity between different land use types, and to promote stability of bird species populations, some management actions should be implemented in the area.
Keywords: bird diversity, species richness, farming systems, land use, tropical dry forest.
Resumen
En el Centro Agropecuario Cotové, en el municipio de Santa Fe de Antioquia (Colombia), se estudió la diversidad de aves utilizando las metodologías de puntos fijos de conteo y redes de niebla en cinco usos del suelo (Frutales, Sistema silvopastoril, Bosque secundario, Pastoreo con baja densidad de árboles y Pastos de corte), para analizar la influencia sobre la composición, riqueza y abundancia de la avifauna. Se registraron 6 633 individuos de 101 especies, de las cuales 11 especies son migratorias. En términos de abundancia, la mayoría de las especies fueron raras y pocas comunes. Los gremios alimenticios mejor representados fueron insectívoros y frugívoros. El índice de Shannon indica que el Sistema silvopastoril presenta los valores de diversidad alfa más altos y los frutales los más bajos. Los usos del suelo que compartieron más especies fueron sistema Silvopastoril y Pastoreo con baja densidad de árboles. La heterogeneidad ambiental del centro actúa de forma diferente para las especies, beneficia en su mayoría aves de baja y media especificidad de hábitat, y con preferencia de hábitats de borde y áreas abiertas, por lo que se deben implementar acciones de manejo dirigidas a la conservación de áreas de regeneración natural y conectividad entre usos del suelo, para que las poblaciones de especies de aves presentes en la zona puedan ser mantenidas en el tiempo.
Palabras clave: diversidad de aves, riqueza de especies, sistemas de cultivo, usos de la tierra, bosque seco tropical.
La pérdida de especies y disminución de poblaciones silvestres está influenciada por diversos factores, entre ellos la fragmentación de ecosistemas (Petit et al., 1999), ya que la conversión de bosques naturales bajo diferentes sistemas de uso del suelo limita la cantidad de hábitat disponible. Por lo tanto, los estudios que incluyen inventarios, análisis de abundancias, comparación de riqueza de especies, análisis de la estructura y la composición de las asociaciones de aves, pueden variar a lo largo del tiempo y permiten tomar decisiones de manejo más acertadas (Remsen, 1994). Las fluctuaciones en los recursos tróficos disponibles (García & Gómez-Laich, 2007), la incorporación de especies migratorias a grupos de especies residentes (Greenberg & Marra, 2005), los factores abióticos, y las comparaciones entre hábitats contrastantes, a su vez, suponen que la lista de especies refleja el valor y carácter ecológico de diferentes lugares, al mostrar la verdadera similitud o disimilitud entre éstos (Balmer, 2001).
Para la zona Andina colombiana se han realizado estudios tendientes a conocer la diversidad y estructura de las comunidades de aves, principalmente en áreas de bosque húmedo, muy húmedo y bosque altoandino, al igual que los efectos que sobre éstas han generado la fragmentación y destrucción de hábitat (Kattan, 1992; Lozano, 1993; Kattan et al., 1994; Stiles, 1998; Stiles & Bohórquez, 2000; Renjifo, 2001; Gallo-Cajiao & Idrobo-Medina, 2004; Duran & Kattan, 2005). A pesar de la importancia en endemismos de los ecosistemas de bosque seco tropical y el fuerte deterioro a que han sido sometidos, el conocimiento de su diversidad de aves es limitado y los estudios realizados se han desarrollado en áreas con alteraciones antrópicas (Herrera, 1974, 1976, Naranjo, 1992; Cárdenas, 2000; Rivera-Gutiérrez, 2006; Castaño et al., 2008). Otros estudios se han ocupado de las preferencias de hábitat para especies particulares en lugar de caracterizar hábitats diferentes en función de su valor ecológico para muchas especies. Ambos enfoques son claramente necesarios en la comprensión de los patrones de uso de hábitat, pero este último enfoque puede ser más útil para establecer prioridades de gestión y conservación del hábitat (Petit et al., 1999).
El ecosistema presente en el Centro Agropecuario Cotové de Santa Fe de Antioquia, se encuentra altamente fragmentado por la expansión de la ganadería y el incremento de fincas de recreo, y los remanentes de bosque que persisten, pueden considerarse el único hábitat de muchas especies de aves, que debido a su capacidad de movilidad cumplen un papel ecológico importante en el control biológico y la dispersión de semillas (Herrera, 1976).
La transformación de los paisajes agropecuarios no permite que ofrezcan servicios ambientales comparables con los ofrecidos por ecosistemas naturales no intervenidos por el hombre. Aunque, es claro que la presencia de remanentes de bosques ofrecen recursos de hábitat y refugio a las aves, por lo que los esfuerzos de conservación deben estar dirigidos a conservar estas áreas por su valor ecológico (Petit et al., 1999), y su potencial como hábitat, corredores biológicos y fuentes de diversidad (Kupfer et al., 2006).
En el Centro Agropecuario Cotové se ha registrado alta diversidad de especies de aves residentes y migratorias que utilizan el cañón del río Cauca en Antioquia (Herrera, 1976), lo que llama la atención por el potencial y el valor ecológico de este agroecosistema. El bosque seco tropical (Holdridge, 1967) es considerado como uno de los ecosistemas prioritarios para la investigación y preservación dentro de las políticas nacionales de biodiversidad (Fandiño & Ferreira, 1998), por estar degradado, fragmentado y ser poco conocido. Algunos estimativos señalan que de bosques secos a subhúmedos en Colombia, sólo existe cerca del 1.5% de su cobertura original de 80 000km2 (Etter, 1993) y menos del 1% de dicha área, se encuentra protegida por el Sistema Nacional de Parques Naturales Nacionales. Adicionalmente, en esta zona de vida se ha establecido gran parte de los sistemas productivos agropecuarios expandiendo la frontera agrícola y pecuaria, y fragmentando las áreas de bosque, y que registra para ecosistemas poco intervenidos gran cantidad de endemismos (IAvH, 1998). En este tipo de bosques, se realizó en 2012 la descripción de la especie Thryophilus sernai, sp. nov., que tiene una distribución restringida al cañón del río Cauca, con algunos individuos o parejas encontrados en los remanentes de bosque y matorrales secos (Lara et al., 2012), destacándose que la nueva especie no es común y está amenazada de extinción debido a la pérdida creciente de los hábitats naturales en el cañón del Cauca y especialmente debido a la inundación planeada por la construcción de la represa hidroeléctrica de Hidroituango.
Para el sitio de estudio, Herrera (1974, 1976) evaluó el impacto de la avifauna sobre los cultivos y aportó un listado parcial de 36 especies para la época, usando una metodología de caza con rifle para estudiar el papel de algunas especies de aves como agentes de control biológico y como dispersores de semillas. Así, resulta importante evaluar la composición y estructura de la avifauna en un área donde la matriz agropecuaria domina el paisaje y los fragmentos de bosque seco pueden desempeñar un papel significativo para la conservación de sus poblaciones. Por lo anterior, este trabajo tuvo como objetivo evaluar la composición, diversidad, abundancia y estructura de la comunidad de aves presentes en cinco diferentes usos de suelo del Centro Agropecuario Cotové, que permita direccionar futuras acciones de manejo para su conservación.
Materiales y Métodos
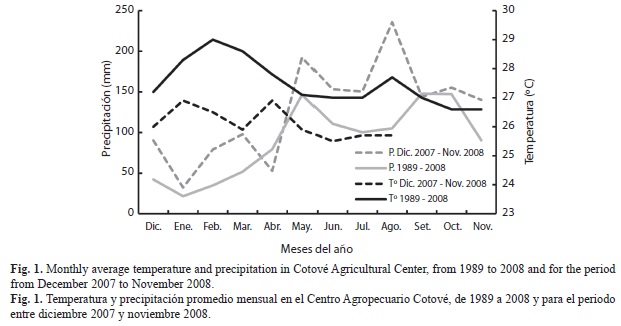
Área de estudio: El Centro Agropecuario Cotové (CAC) se ubica en la vereda el Espinal del municipio de Santa Fe de Antioquia, departamento de Antioquia, Colombia (6º33´32” N - 77º04´51” W; 530msnm) y tiene una extensión de 121 hectáreas en una zona de vida de Bosque seco Tropical (bs-T) (De los Ríos & Gallego, 2003). De acuerdo con los datos de la estación meteorológica 2622503, la temperatura promedio anual entre 1989 y 2008 fue de 27.5ºC, la humedad relativa de 73% y el promedio anual de precipitación de 1 078mm. Para el período de estudio, entre 2007 y 2008, la temperatura promedio mensual fue 26°C y la precipitación1 446.1mm con un pico de lluvias en mayo y otro en agosto (IDEAM, 2008) superiores al promedio de los últimos 20 años (Fig. 1). El estudio se realizó en los siguientes cinco usos del suelo con diferentes tipos de manejo:
Frutales (Fr): Esta área presenta un relieve plano, compuesto en su mayoría por árboles de mango (Mangifera indica) sembrados a principio de la década de los años 90 del siglo pasado, en un arreglo en forma de cuadricula, con una distancia de siembra entre ellos de 5x5m, y en menor proporción por otras especies como el zapote (Matisia cordata), tamarindo (Uribea tamarindoides), pomarrosa (Syzygium malaccense), coco (Cocos nucifera), palma africana (Elais guianenses), ciruelo (Spondias sp.) y caimito (Chrysophyllum cainito), guácimo (Guazuma ulmifolia), piñón de oreja (Enterolobium cyclocarpum) y mamoncillo (Meliccoca bijugatos). El suelo se encuentra cubierto por hojarasca y arvenses en un 57%, que solo en casos excepcionales superó los 50cm, una cobertura de dosel de 68% y 10m de altura.
Pastoreo con baja densidad de árboles (Pb): Caracterizado por un relieve de pendientes muy suaves donde se mantiene cría de ganado Brahman pastando en potreros dominados por guinea (Panicum maximum) y colosoana (Botriochloa pertusa), con uno a tres árboles adultos de piñón de oreja (Enterolobium cyclocarpum) por hectárea, de una altura media de 9m. Presencia de arbustos espinosos y abundante regeneración natural sobre todo arvenses de menos de 50cm de altura, y bordeados de cercas vivas con matarraton (Gliricidia sepium). En las inmediaciones se localiza un área de protección (bosque secundario) perteneciente a otro predio.
Bosque secundario (Bs): Formado por un remanente de bosque seco en un valle con suaves pendientes, abundantes enredaderas y arbustos espinosos. Con predominio de vegetación arbustiva densa y árboles adultos, producto de la regeneración natural. Cobertura de dosel de 84% y a una altura de 17m. Las especies más representativas fueron tachuelo (Xanthoxilum sp.), carbonero (Leucaena leucocephala), piñón de oreja (Enterolobium cyclocarpum), indio desnudo (Bursera simarouba), mamoncillo (Meliccoca bijugatos), diomate (Astronium sp), ceiba verde (Pseudobombax sp.), ciruelo (Spondias sp.). Algunas herbáceas leñosas como santa maría (Euphatorium odoratum), escoba (Sida rhombifolia) y zarza (Mimosa nigra).
Sistema silvopastoril (Ss): Área de pendientes suaves e irregulares, de pastos altos y densos con presencia de hojarasca y arvenses que superan los 50cm. Dedicada a la cría de ganado Brahman en potreros con árboles y arbustos dispersos (entre 50 y 60 árboles adultos por hectárea) producto de la regeneración natural, con una altura media de 17m que proporcionan sombra. Las especies más características fueron: olla de mono (Lecythis minor), matarraton (Gliricidia sepium), piñón de oreja, (Enterolobium cyclocarpum), ciruelo (Spondias sp.), caña fístula (Cassia espectabilis), tachuelo (Xanthoxilum sp.), diomate (Astronium sp.), mamoncillo (Meliccoca bijugatos), indio desnudo (Bursera simarouba), y en menor proporción carbonero (Leucaena leucocephala).
Pastos de corte (Pc): Área plana sin árboles, con pasto angleton (Dishanthium aristatum), cuyo forraje es de aproximadamente de 50-70cm de alto, con arvenses de 20cm de altura y está rodeado de especies como: piñón de oreja (Enterolobium cyclocarpum), indio desnudo (Bursera simarouba), carbonero (Albizia ferruginia), ciruelo (Spondias sp.), diomate (Astronium sp.), mamoncillo (Meliccoca bijugatos), matarraton (Gliricidia sepium), coco (Cocos nucifera), entre otros. Aquí se realizan cortes de pasturas para la elaboración de heno cada 35 días de manera mecanizada.
Métodos de campo: Se realizaron cinco salidas de campo: tres en diciembre de 2007, febrero y abril de 2008 que corresponden al período seco del año, y dos en junio y octubre de 2008 en período de lluvias. Para la recolección de datos se utilizaron dos metodologías complementarias. (1) Puntos fijos de conteo: realizados mediante el uso de binoculares, donde se tomaron registros visuales de las aves presentes en un radio de 25m durante diez minutos para cada uso del suelo. Esto se hizo de acuerdo a la rotación mostrada en el cuadro 1, de tal forma que cada día se iniciaba en un uso distinto, garantizando que se muestreara cada tipo de uso de suelo en los diferentes momentos del día. En cada uso del suelo se tomaron tres estaciones de conteo espaciadas con un mínimo de 150m de distancia y separada más de 25m del borde. Los muestreos fueron realizados en tres intervalos de tiempo: entre las 06:00-09:00 horas, entre las 10:00 y 12:10 horas y entre las 15:00 y 17:10 horas para cada uso del suelo por cinco observadores de forma simultánea (Ralph et al., 1993; Bibby et al., 2000). La intensidad del muestreo por este método fue de 225horas/observador. (2) Captura con redes de niebla: en cada salida se ubicaron ocho redes de 12m de largo y dos metros de ancho a una altura de un metro del piso, en cada sitio de muestreo durante un día, para un total de 320horas/red por cada uso del suelo, y 1 600horas/red para todos los usos del suelo. Adicionalmente, se tomó nota de las especies de aves detectadas durante los recorridos por fuera de los puntos fijos y de las capturas con redes. Para la identificación de las aves en campo se utilizaron las guías de Hilty & Brown (1986), Fundación Los Yalcones (2000) y Restall et al. (2007), y la lista de especies se ordenó taxonómicamente de acuerdo a este último autor.
Análisis de información: Se evaluó el esfuerzo de muestreo y la riqueza de especies a través de curvas de acumulación de especies y el estimador de Chao1, obtenidos a través del programa EstimateS versión 8.0 (Collwell, 2006).
Se calcularon los índices de diversidad alfa (Shannon y Equidad de Shannon) y beta (similitud de Jaccard) a través del programa EstimateS versión 8.0 (Collwell, 2006). Las detecciones realizadas durante los recorridos por fuera de los puntos fijos, no fueron tomadas en cuenta en este análisis y se usaron solamente en el listado de especies, ya que se buscó utilizar una metodología homogénea en todos los usos de suelo para realizar comparaciones entre ellos. Se comprobaron los modelos de abundancia de especies de serie logarítmica, geométrica, Broken Stick y Log normal, descritos por Magurran (1988). El ajuste de una serie de datos a un modelo se determinó mediante la prueba X2(Chi cuadrado) con un nivel de significancia de 0.05.
Las especies detectadas se ordenaron sistemáticamente de acuerdo con la guía de Hilty & Brown (1986). Se analizaron las variaciones de abundancia y riqueza de especies en dos épocas (lluvia y seca) en los cinco usos del suelo, mediante pruebas de rangos de Wilcoxon. La abundancia de especies detectadas en más de tres usos del suelo se examinó a través de un análisis de varianza de dos vías, donde la abundancia de cada una de las especies fue transformada por la raíz cuadrada de la abundancia más 0.5. La comparación de medias se realizó por medio de la prueba de Tukey.
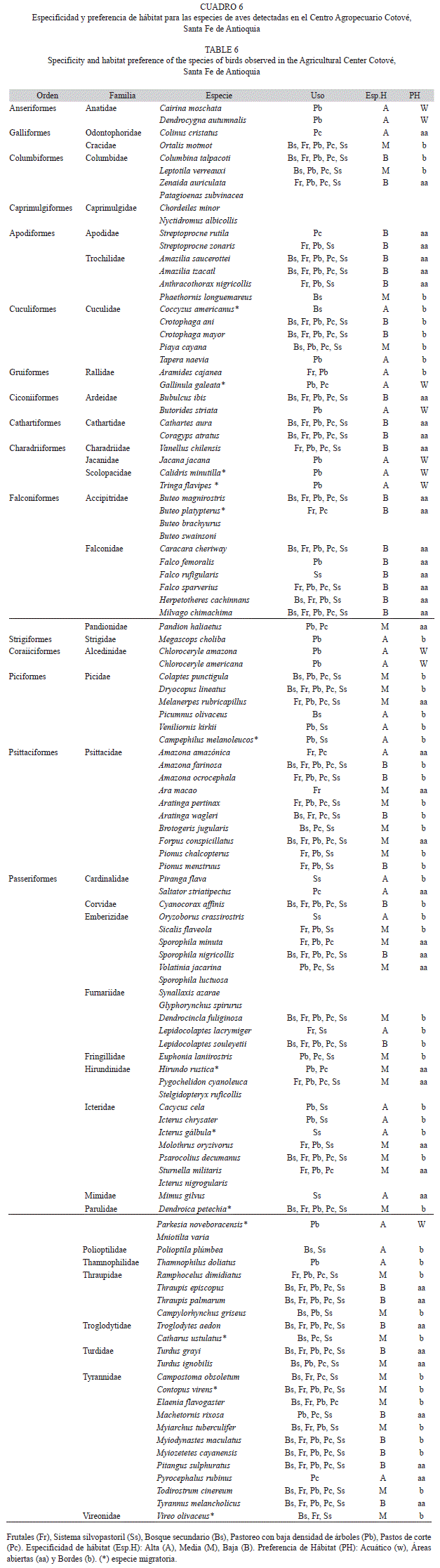
En el análisis de la estructura de la comunidad, la preferencia de hábitat para cada especie fue realizada de acuerdo con Robinson (1999) dentro de cuatro categorías: (1) áreas abiertas, para especies asociadas a áreas de producción agropecuaria y fincas de recreo, y especies que la mayor parte del tiempo permanecían en vuelo por encima del dosel; (2) interior de bosque, para especies detectadas en el interior de bosques secundarios y rastrojos altos; (3) bordes, para especies asociadas a bordes de zonas de rastrojo alto; y (4) acuáticas para aves detectadas exclusivamente haciendo uso de los cuerpos de agua y de naturaleza típicamente acuática.
La especificidad de hábitat se evaluó de acuerdo a Stotz et al. (1996) y Castaño & Patiño (2000), teniendo en cuenta el grado de dependencia de cada especie con respecto a su hábitat: alta, para especies restringidas a un solo tipo de hábitat (boscosas, bordes o abiertas); media, para especies detectadas en al menos dos tipos de hábitat (boscosas-bordes o bordes-abiertas); baja, para especies que no presentan restricciones a un tipo de hábitat.
Además, se determinaron las categorías de residencia de acuerdo a Salaman et al. (2008), Remsen et al. (2011), Naranjo et al. (2012) como residentes o migratorias, y se clasificaron en siete gremios tróficos: frugívoros, nectarívoros, granívoros, insectívoros, omnívoros, carnívoros y carroñeros, siguiendo la propuesta de Hilty & Brown (1986) y Restall et al. (2007). Se empleó un análisis de varianza de doble vía a través del programa SAS (8.0), donde los factores fueron la época de muestreo (lluvia y seca) y los diferentes gremios tróficos, para comparar la riqueza y abundancia de especies.
Resultados
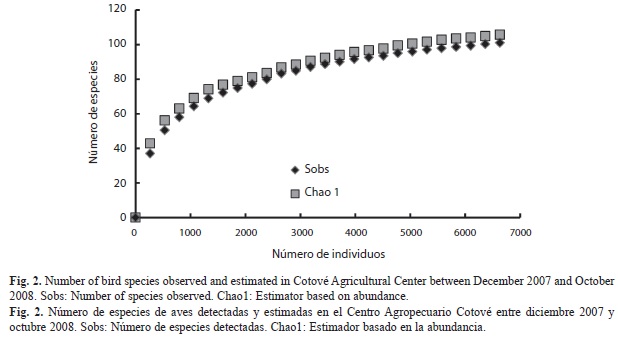
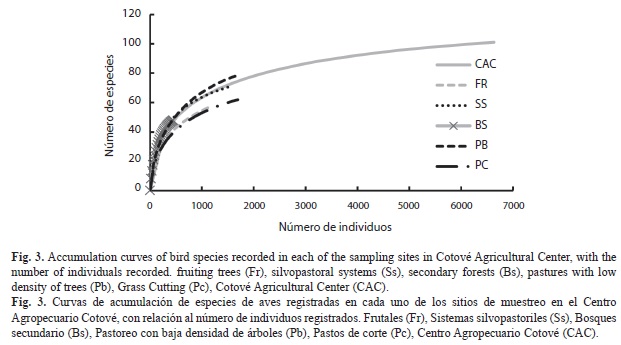
Evaluación de la calidad del censo de especies: El estimador de Chao 1 presenta una tendencia similar a la encontrada para la acumulación de especies en el Centro Agropecuario Cotové, siendo las 101 especies detectadas un 96% de las esperadas (Fig. 2). El número de especies reportado para cada uso del suelo muestra un valor cercano al esperado según el estimador Chao 1 en los usos de suelos de Pastoreo con baja densidad de árboles y Sistema silvopastoril, mientras que para los otros usos de suelo el valor reportado se aleja de éste (Cuadro 2). La curva de acumulación de especies para el Centro Agropecuario presenta una asíntota, mientras que para cada uso del suelo en particular sigue en aumento (Fig. 3).
Composición y riqueza de aves: Se registraron 6 633 individuos pertenecientes a 101 especies. Durante las 1 600horas/red se capturaron 167 ejemplares de 36 especies, y se detectaron visual y/o auditivamente 6 466 individuos de 100 especies. Phaethornis longuemareus fue la única especie que sólo se registró por redes. Fuera de los muestreos sistemáticos se observaron otras once especies: Buteo brachyurus, Buteo swainsoni, Patagioenas subvinacea, Chordeiles minor, Nyctidromus albicollis, Synallaxis azarae, Glyphorynchus spirurus, Stelgidopteryx ruficollis, Sporophila luctuosa, Icterus nigrogularis y Mniotilta varia.
Se detectaron especies exclusivas para cada uso del suelo, para Bosque secundario se encontraron: Phaethornis longuemareus, Polioptila plumbea y Picumnus olivaceus; en frutales: Amazona amazonica y Ara macao; en Sistemas silvopastoriles: Piranga flava y Mimus gilvus; en Pastos de corte: Streptoprocne rutila, Pyrocephalus rubinus, Colinus cristatus y Saltator striatipectus; para Pastoreo con baja densidad de árboles se detectaron trece especies: Cairina moschata, Jacana jacana, Calidris minutilla, Tringa flavipes, Chloroceryle amazona, Chloroceryle americana, Parkesia noveboracensis, Dendrocygna autumnalis, Tapera naevia, Thamnophilus doliatus, Butorides striata, Megascops choliba y Falco femoralis, de estas las nueve primeras fueron detectadas exclusivamente en época de lluvia.
Para el Centro Agropecuario Cotové, en su conjunto, las familias de mayor número de especies fueron Tyrannidae (11), seguido de Psittacidae (10) y las familias Icteridae, Falconidae y Picidae con seis especies cada una.
El análisis de la riqueza de especies a través de la prueba de rangos determinó que no existió diferencias significativas (Tukey, p>0.05), entre el número de especies en la época seca (18.5±5.21) y la época de lluvias (18.7±4.67).
Para la riqueza de especies, no se encontró diferencias significativas (Tukey, p>0.05) entre los diferentes usos del suelo. Se registró mayor número de especies en Pastoreo con baja densidad de árboles (79), seguido de Sistemas silvopastoriles (71), Pastos de corte (62), Frutales (58) y Bosque secundario (48), respectivamente (Cuadro 2).
Diversidad alfa y beta: La diversidad alfa analizada a través del índice de Shannon, indicó que existieron diferencias significativas (Tukey, p<0.05; F=36.06) entre los distintos usos del suelo, siendo Sistema silvopastoril el que presentó mayor diversidad de especies (H´: 3.39), seguido del uso de Pastoreo con baja densidad de árboles (H´: 3.21). Los usos de suelo de Bosque secundario y Pastos de corte, no presentaron diferencia significativa (Tukey, p>0.05) entre sí, pero su diversidad fue significativamente menor a la de los dos primeros, y el uso que presentó menor índice de diversidad fue el de Frutales (H´: 2.65) (Cuadro 2). Para el índice de Equidad de Shannon los usos del suelo Sistema silvopastoril y Bosque secundario no presentaron diferencias significativas (Tukey, p>0.05) entre sí y mostraron una mayor equidad en el número de individuos de las diferentes especies. Para los demás usos del suelo se encontraron diferencias significativas (Tukey, p<0.05) en el índice de equidad que disminuyó en el siguiente orden: Pastoreo con baja densidad de árboles, Pastos de corte y Frutales.
Se observa un bajo valor del índice de similitud de Jaccard entre la mayoría de usos, siendo el valor más alto el de Sistema silvopastoril con los usos Frutales (0.654) y Pastoreo con baja densidad de árboles (0.648). Por su parte, entre el Bosque secundario y los demás usos se presentó baja similitud de especies, en especial con el uso de Pastoreo con baja densidad de árboles (0.443) (Cuadro 3).
Modelos de abundancia de especies: Las series que presentaron mejor ajuste fueron la serie Log normal y la serie logarítmica (Chi-cuadrado, p<0.05) (Cuadro 4).
Análisis de abundancias: De acuerdo con las pruebas de rangos, la abundancia de aves en el Centro Agropecuario Cotové en las diferentes épocas del año (lluvia y seca) no mostró diferencias significativas (Tukey, p=0.68), pero al analizar los datos por usos del suelo de manera separada se encontró que solo Frutales y Bosque secundario tienen diferencias significativas (Tukey, p<0.05).
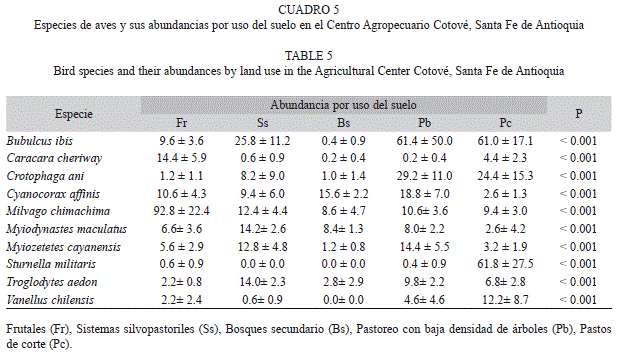
Por su parte, los análisis de varianza para las especies, mostraron que de las 101 especies, solamente 63 especies fueron susceptibles de ser analizadas, por estar presentes en al menos tres usos del suelo, y de estas diez presentaron diferencias estadísticamente significativas (p<0.001; Cuadro 5).
Preferencia y especificidad de hábitat: La preferencia de hábitat, señaló que la mayoría de las especies presentes en el Centro Agropecuario fueron de áreas abiertas (41.58%) y de áreas de borde (48.5%), presentándose sólo diez especies con preferencia de hábitats acuáticos y ninguna de interior de bosque. La especificidad de hábitat indicó que la mayoría de las especies (36.6%) fueron de baja especificidad, es decir, se encontraron en cualquier uso del suelo indistintamente. De especificidad media hubo un 33.6% y de especificidad alta un 29.7%, que sólo se localizaron en uno o dos usos del suelo, principalmente en Pastoreo con baja densidad de árboles, donde se asociaron especies de preferencia de hábitat acuático y en Bosque secundario (Cuadro 6).
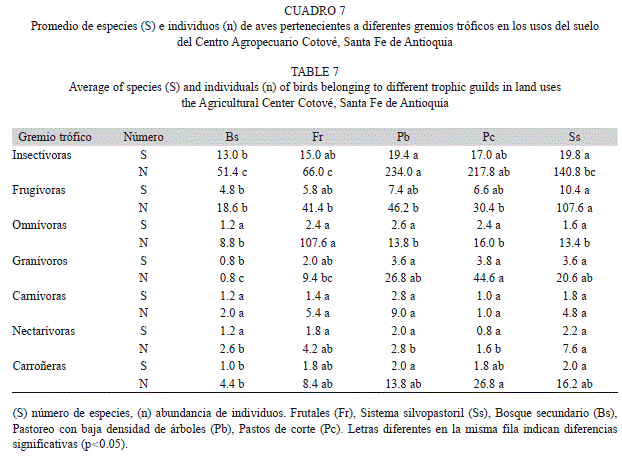
Gremios tróficos: De las 101 especies detectadas en los muestreos sistemáticos se encontraron 47 insectívoras, 21 frugívoras, nueve omnívoras, nueve granívoras, nueve carnívoras, cuatro nectarívoras y dos carroñeras. El número promedio de especies y su abundancia por uso de suelo muestra que Sistema silvopastoril albergó el mayor número de especies insectívoras, frugívoras, nectarívoras y carroñeras. Pastoreo con baja densidad de árboles registró el mayor número de especies omnívoras, carnívoras y carroñeras, y en Pastos de corte se presentó el mayor número de especies granívoras. Se encontró un mayor número de especies insectívoras y frugívoras en los usos de Sistema silvopastoril y Pastoreo con baja densidad de árboles (Cuadro 7). Las especies omnívoras, carnívoras y nectarívoras no presentaron diferencias significativas (Tukey, p>0.05) con relación al número de especies. Sin embargo, con relación a las abundancias, las omnívoras presentaron diferencias significativas (p<0.05) en Frutales (Milvago chimachima). En Bosque secundario se presentaron diferencias significativas (Tukey, p<0.05) presentando este uso de suelo el menor número de especies y de ejemplares de los gremios frugívoros, insectívoros, granívoros y carroñeros.
Las especies granívoras fueron comunes en las áreas típicas de pastos, Sistema silvopastoril, Pastos de corte y Pastoreo con baja densidad de árboles. Las especies carnívoras se encontraron indistintamente en los diferentes usos del suelo, con bajas abundancias. Por su parte, los carroñeros representados por las especies Cathartes aura y Coragyps atratus se avistaron en todos los usos del suelo, con mayor facilidad en usos del suelo que tienen áreas abiertas como Pastos de corte y Pastoreo con baja densidad de árboles y en menor proporción en los usos del suelo con mayor arbolado (Cuadro 7).
Especies migratorias: Entre las especies migratorias se reportaron 11 durante los muestreos sistemáticos: Buteo platypterus, Calidris minutilla, Tringa flavipes, Coccyzus americanus, Contopus virens, Catharus ustulatus, Dendroica petechia, Hirundo rustica, Parkesia noveboracensis, Icterus galbula, Vireo olivaceus y dos por fuera de los muestreos (Buteo swainsoni y Mniotilta varia), para un total de 13 especies migratorias detectadas para el Centro Agropecuario.
Discusión
La convergencia de las curvas de acumulación de especies de aves detectadas para todo el Centro Agropecuario y del estimador Chao1 indica que se logró muestrear cerca de un 96% de las especies esperadas. Sin embargo, los valores observados para cada uso de suelo en las curvas de acumulación de especies se alejan de los estimados, lo que se debe en gran medida a que las áreas de cada uso de suelo dentro del Centro, son reducidas y existe una alta posibilidad de que las especies sean detectadas en cualquiera de ellos, por lo que se espera que con un incremento en el muestreo en cada uso, el número de especies aumente. Por su parte, el análisis de la riqueza de especies determinó que no existieron diferencias significativas entre el número de individuos y especies entre la época seca y la época de lluvias. Sin embargo, la composición de especies si fue diferente entre períodos, observándose una alta tasa de recambio de especies detectadas, que puede deberse a variaciones de las condiciones ambientales y de la disponibilidad de recursos alimenticios, que hace que diferentes especies accedan a recursos variados en diferentes períodos de tiempo.
El número de especies reportadas para el Centro Agropecuario Cotové se incrementó pasando de 36 registradas por Herrera (1974) y Herrera (1976), a 112 para el presente estudio, incluyendo las 11 especies detectadas por fuera de los muestreos sistemáticos. El alto número de especies encontradas en el Centro Agropecuario Cotové, coincide con los valores de riqueza específica encontrada en otros estudios realizados en áreas de Bosque Seco Tropical, como los reportados por Naranjo (1992) con 141 especies y Cárdenas (2000) con 134 especies para zonas ganaderas y centros de producción agropecuaria en el Valle del Cauca, y de 114 especies por Rivera-Gutiérrez (2006) para un parche de bosque secundario cercano a la ciudad de Cali, Colombia.
De acuerdo con Haffer (1967) y Stotz et al. (1996), la avifauna típica de Bosque seco Tropical es una mezcla de elementos de zonas áridas y húmedas, por lo que posee un número reducido de aves especialistas y es considerado para las comunidades de aves, como un ecosistema de transición dentro de un gradiente climático, lo que conlleva a un alto número de especies generalistas en este tipo de bosque. La zona de vida de bosque seco tropical presenta ensamblajes de especies particulares, lo que le confiere gran importancia a cada remanente existente si se busca conservar una muestra representativa de poblaciones de aves viables en los bosques secos de Colombia (Chaves & Arango, 1998). A su vez, el registro de especies exclusivas en cada uno de los diferentes usos de suelo, refleja la capacidad de adaptación de las especies de aves a la fragmentación y deterioro del hábitat, y muestra la importancia que tiene la conservación de áreas espacialmente heterogéneas que ofrecen diversidad de hábitats y coberturas arbóreas, para el mantenimiento de las poblaciones de muchas especies en el sustento de la biodiversidad, tal como lo han encontrado Pearce-Higgins (2000), Blendinger & Álvarez (2002) y Brandolin et al. (2007).
La mayor diversidad de aves encontrada se presentó en el uso de Sistema silvopastoril, lo que coincide con lo reportado por Naranjo (1992), Ramírez-Albores (2006), Codesido & Bialenca (2004). Se encontraron 47 especies en al menos cuatro usos del suelo y el 80% estaban asociadas con los bordes, lo que está relacionado con los movimientos a través de los conectores del paisaje como cañadas arboladas, cercas vivas y árboles dispersos en pasturas. Esto muestra la importancia del mantenimiento de relictos de bosques naturales y de sistemas silvopastoriles en la conectividad de los parches de hábitat para las aves, y el mantenimiento de flujos entre diferentes áreas de alimentación y refugio.
La menor riqueza de especies se presentó en el fragmento de Bosque secundario, lo que puede obedecer al menor tamaño de esta área con relación a los otros usos de suelo. Esto coincide con lo reportado por Borges & López-Mata (2005) para una selva mediana subperennifolia en México, quienes afirman que la riqueza y abundancia de especies disminuye al decrecer el área. Además, se observó que durante el día algunas de las aves salían del área de bosque hacia los usos aledaños, donde encuentran mayor disponibilidad de recursos, y quedan expuestas a aves como Milvago chimachima, Caracara cheriway y Cyanocorax affinis, las cuales fueron reportadas por Ramo & Busto (1984) como depredadoras de nidos de Passeriformes en los llanos de Apure, en Venezuela. El aumento del efecto borde por la alta fragmentación hace que aumente la depredación y el parasitismo de nidos que se ven mayormente expuestos, lo que provoca un decrecimiento poblacional de las poblaciones de aves (Stephens et al., 2003; Gustafson et al., 2002). Esto conlleva a que las poblaciones de aves que persisten en este uso del suelo estén expuestas a una declinación, debido a la reducción de espacios territoriales y áreas de anidación, como posiblemente sucede con Picumnus olivaceus, Phaethornis longuemareus y Coccyzus americanus que rara vez fueron detectadas en este uso del suelo asociadas a áreas de borde, por lo que se considera importante la conservación y restauración de estos fragmentos para el mantenimiento de las poblaciones de estas especies.
El Bosque secundario posee una vegetación producto de la sucesión natural que favorece a especies con hábitos rastreros poco conspicuas tales como Leptotila verreauxi y sigilosas como Piaya cayana, Elaenia flavogaster, Coccyzus americanus, Campostoma obsoletum, Campylorinchus griceus, Polyoptila plumbea, Tapera naevia y Aramides cajanea. Igualmente, se observó que otras especies pasan de largo, por encima o por debajo del dosel camino hacia otros lugares, como: Aratinga wagleri, Brotogeris juguralis, Bubulcus ibis, Buteo magnirostris, Cathartes aura, Crotophaga ani, Cyanocorax affinis, Dendrocincla fuliginosa y Coragyps atratus. A pesar de la menor riqueza de especies observada en el Bosque secundario, éste ofrece un sitio de resguardo para muchas especies de aves residentes en la zona, y eventualmente, como zona de abastecimiento de insectos como alimento para las especies migratorias Catharus ustulatus y Coccyzus americanus que llegan a la zona (Duran & Kattan, 2005). Estas especies pueden ser capaces de tolerar la pérdida de bosques naturales ocupando ambientes modificados (Norton et al., 2000).
Los usos de riqueza intermedia de especies fueron, Pastos de corte, Frutales y Sistema silvopastoril. Estas zonas permiten el acceso de gran variedad de aves de hábitos generalistas y de amplia distribución, ya que los árboles dispersos pueden proveer valiosos recursos y refugio para las aves. Naranjo (1992) y Cárdenas (2000) han reportado una alta riqueza de aves en diferentes sistemas de producción con predominio de áreas ganaderas, quienes al igual que en este trabajo mencionan la presencia de pocas especies típicas de ambientes poco alterados con poblaciones muy reducidas y un alto número de especies generalistas características de ambientes intervenidos. En frutales, la diferencia en la abundancia entre las épocas (lluvia y seca), puede estar relacionada con la disponibilidad de alimento. Los sistemas silvopastoriles, pueden soportar un número significativamente más grande de especies de aves que las pasturas abiertas con densidades muy bajas de árboles y arbustos (Naranjo, 2001; Múnera et al., 2008; Cárdenas, 2000). Además, adicionan condiciones favorables para la comunidad de aves, permitiendo que se establezcan fácilmente algunas especies que no podrían mantenerse en áreas descubiertas de vegetación arbórea y proporcionan hábitat que puede facilitar la persistencia de algunas especies de plantas y animales dentro de paisajes fragmentados, mitigando parcialmente los impactos negativos de la deforestación y la fragmentación sobre la biodiversidad (Naranjo,1992; Greenberg et al., 1997; Pezo & Ibrahim, 1998; Harvey et al., 2000; Waltert et al., 2004).
Otros estudios indican que una significante porción de la biodiversidad original puede ser mantenida dentro de sistemas de producción agropecuarios apropiadamente diseñados y manejados (Greenberg et al., 1997; Harvey et al., 2000), donde se favorecen principalmente especies de áreas abiertas o de bordes (Primack, 2002), que evaden las áreas de bosque que son más oscuras y altas con un microclima diferente al de los usos agropecuarios circundantes que están más abiertos, luminosos y con diferentes estratos vegetales que ofrecen variedad de recursos (Ricketts, 2001).
La riqueza de especies resultó mayor en el área de Pastoreo con baja densidad de árboles, con 79 especies, debido a que en temporada de lluvias, en octubre 2008, se formó un encharcamiento temporal al que llegaron las especies Cairina moschata, Jacana jacana, Calidris minutilla, Tringa flavipes, Chloroceryle amazona, Chloroceryle americana, Parkesia noveboracensis, Dendrocygna autumnalis, Butorides striata y Tapera naevia. Adicionalmente, la configuración espacial encontrada en este uso del suelo la hace un lugar de paso de gran importancia para aves de todos los gremios alimenticios encontrados, y el bajo porcentaje de cobertura de dosel favorece el avistamiento de aves adaptadas a la intervención antrópica y los árboles en pie ofrecen sitios de percha, de alimentación y anidación (Lynch, 1989). El uso del suelo de Pastos de corte presenta gran cantidad de especies de aves detectadas considerando la relativa homogeneidad de su fisionomía, y aves como Bubulcus ibis, Vanellus chilensis, Zenaida auriculata, Crotophaga mayor y Crotophaga ani, aprovechan la abundancia súbita de invertebrados y pequeños vertebrados que se presentan disponibles al momento del corte de los pastos (Cárdenas, 2000).
El índice de Equidad de Shannon en los usos de suelo de Bosque secundario (0.83) y Sistema silvopastoril (0.85) presentan los mayores valores, lo que permite inferir que las abundancias presentes en estos usos son más similares, sin que se presente una dominancia marcada de algunas especies. Esto se relaciona con una mejor distribución de recursos, porque las características del hábitat (número de especies arbóreas, densidad de árboles y arbustos y cobertura de dosel) permiten una mejor distribución vertical de las especies de diferentes estratos que son aprovechados por los diferentes grupos tróficos (Verea et al., 2000). A su vez índice de equidad de Shannon para los usos de suelo Pastoreo con baja densidad de árboles, Pastos de corte y Frutales presentan los menores valores. El valor más bajo de equidad en Frutales (0.71), es producto de la alta dominancia que presentaron las especies Milvago chimachima y Caracara cheriway, consideradas en este estudio como especies omnívoras, las cuales se ven favorecidas por el aumento de áreas abiertas para el pastoreo de ganado (Duran & Kattan, 2005), y de lugares de anidación y alimentación. El uso de Pastoreo con baja densidad de árboles presentó una dominancia de especies como Bubulcus ibis, Cyanocorax affinis y Crotophaga ani, especies adaptadas a áreas abiertas, y en Pastos de corte de Sturnella militaris capaz de anidar en los pastos, Vanellus chilensis y Bubulcus ibis, aves típicas de pastos y que se alimentan de insectos presentes en éstos.
Las diferencias en la composición de especies entre usos del suelo, expresado a través del índice de Jaccard, mostró valores que oscilaron entre 0.44 y 0.65, lo que implica tasas importantes de recambio de especies entre los diferentes usos de suelo, mostrando que los usos que presentan mayor intervención antrópica tienen valores de similitud mayor entre sí, como es el caso de Sistema silvopastoril que presenta los valores más altos de similitud con los usos de Frutales y Pastoreo con baja densidad de árboles. Esto se debe principalmente a que se presentan especies de aves con preferencias de hábitats abiertos y de borde, y con menor especificidad de hábitat que utilizan indistintamente estos usos; pero cabe destacar que entre los usos Pastoreo con baja densidad de árboles y Bosque secundario se presentaron los valores más bajos del índice de Jaccard (0.44), compartiendo menor número de especies y presentando un mayor recambio de ellas. Esto puede deberse a las marcadas diferencias en las condiciones ambientales como densidad de arbustos, cobertura y altura de dosel, que generan condiciones ambientales diferentes (Koleff, 2005).
El ajuste de la comunidad de aves a un modelo de abundancia de serie logarítmica se debe a que la zona de estudio es un área sometida a perturbaciones propias de los sistemas de manejo agropecuarios con la dominancia de muy pocas especies, y el ajuste a la serie log-normal, a que se ha mantenido el disturbio por mucho tiempo, lo que hace que se muestre un equilibrio en cuanto al uso del recurso, pero aun así la comunidad de aves se mantiene disturbada (Shmida & Wilson, 1985).
Por lo tanto, el Centro Agropecuario Cotové es un área dinámica e interactuante, donde las aves de los diferentes gremios tróficos se mueven entre los usos del suelo en búsqueda de alimento, con mayor representación de insectívoras, seguido de frugívoras, y en menor proporción nectarívoras, granívoras y carnívoras, siendo ésta distribución, típica de los bosques en todo el Neotrópico (Karr et al., 1990). Así, los insectos representan un alimento abundante para las aves que no varía significativamente en el tiempo (Idrobo-Medina & Gallo-Cajiao, 2004), y las aves poseen una gran variedad de técnicas para consumirlos, convirtiéndose en un recurso primordial para el mantenimiento de la comunidad y de su estructura trófica. Además, las especies insectívoras se mueven regularmente a través de muchas especies de árboles y usos del suelo (Cárdenas, 2000). Según Duran & Kattan (2005), las especies frugívoras aprovechan la heterogeneidad ambiental para desplazarse en búsqueda de alimento. Esto puede explicar la presencia de este gremio trófico en todos los usos del suelo del Centro Agropecuario Cotové, donde se encontraron árboles frutales dispersos en pasturas. Por otra parte, las especies nectarívoras suelen aprovechar de manera general áreas de borde para su alimentación, pues para los colibríes son más importantes los recursos alimenticios, que el grado de antropogenización del paisaje donde se encuentren (Kattan & Murcia, 2003).
Las aves migratorias se mueven a través del paisaje rural en búsqueda de alimento y refugio. Su presencia en Centro Agropecuario Cotové puede deberse a la gran pérdida y fragmentación de ecosistemas boscosos que ha causado que muchas especies de aves migratorias deban ocupar temporalmente el paisaje agropecuario que posee una menor calidad ambiental para continuar con la migración (Kurosawa & Askins, 2003; Johnson et al., 2005; Duran & Kattan, 2005), pero que para algunas especies como Dendroica petechia pueden aportar recursos alimenticios necesarios para que sigan su migración, funcionando muchas veces como oasis dentro de las matrices productivas (DeClerck et al., 2011). Sin duda, una finca cubierta de árboles, puede funcionar como un corredor biológico espacialmente heterogéneo que contribuye a la conservación de la biodiversidad, suministrando hábitats y recursos para las especies animales y vegetales (Naranjo, 1992; Harvey et al., 2008). Al respecto, Johnson et al. (2005) afirman que los hábitats agrícolas arbóreos en los trópicos pueden ser útiles para la conservación de aves insectívoras generalistas, incluyendo muchas paserinas migratorias como Hirundo rustica, Icterus galbula y Dendroica petechia.
La pérdida de conectividad de los diferentes fragmentos de bosque y la cada vez más acentuada transformación de los predios en la región, han podido conducir a la reducción y pérdida de poblaciones, por lo menos a nivel local, de especies que fueron reportadas por Herrera (1974, 1976) para el Centro Agropecuario Cotové, y que no fueron detectadas en el presente estudio como Piranga rubra, Ramphastos swainsonii, Elanoides forficatus, Theristicus caudatus, y Setophaga castanea. Sin embargo, se debe realizar un monitoreo permanente que permita hacer una evaluación del estado de sus poblaciones. Al respecto, Bierregaard et al., (1992) aseguran que muchas especies de aves tropicales residentes permanentes son más sensibles que las migrantes a la degradación y pérdida del hábitat, tal vez debido a un mayor nivel de especialización de nichos y requisitos estrechos de microhábitat o microclima. Kattan (1992) asegura que especies frugívoras y de interior de bosque con regularidad son más propensas a la extinción, debido a que no se desplazan entre fragmentos a través de los claros.
Por otra parte, la presencia de las especies Dendrocincla fuliginosa, Lepidocolaptes lacrymiger y Lepidocolaptes souleyetii, que se detectaron en varios usos del suelo, y que son propias de áreas de bosque bien conservados, evidencian la importancia del Centro Agropecuario Cotové para la protección de estas especies con susceptibilidad a la extinción (Stiles & Bohórquez, 2000; Castaño & Patiño, 2000).
En general, las especies de aves del Centro Agropecuario Cotové se han visto afectadas por las perturbaciones ambientales propias de agroecosistemas, pero al mismo tiempo algunas se adaptan a estos cambios de acuerdo a su plasticidad ecológica y otras se ven altamente afectadas con la reducción drástica de sus poblaciones (Barrett et al., 1994; McIntyre & Hobbs, 1999). Para la comunidad de aves en el Cañón del río Cauca, el aumento de las fincas de recreo, procesos urbanísticos y la ampliación de la frontera agropecuaria, ha reducido drásticamente la disponibilidad de recursos y ha modificado los hábitats originales de bosque seco tropical, viéndose favorecidas las especies de aves con preferencia a áreas abiertas o de borde y con baja especificidad de hábitat. A corto y mediano plazo se puede generar el decline de las poblaciones de diferentes especies de aves que se pueden ver sometidas a procesos marcados de estocasticidad demográfica y que deben realizar desplazamientos entre los fragmentos de bosques o áreas que presentan aún coberturas arbóreas y que obedecen a procesos de partición de nichos que permiten aprovechar los recursos de manera diferencial en el tiempo y el espacio. Kattan (1992) afirma que en una escala espacial mayor, las especies que se mueven entre fragmentos son menos probables que estén localmente extintas, porque sus poblaciones pueden ser aumentadas a través de inmigración y recolonización.
El Centro Agropecuario al mantener una cobertura vegetal permite que especies de aves que no requieren estrictamente el hábitat de bosque o condiciones de baja perturbación como los Tyranidos y Cuculiformes, y que tienen especificidad de hábitat baja o media, puedan encontrar recursos para mantener sus poblaciones, indicando el potencial para la conservación de las especies en áreas pequeñas con adecuada cobertura vegetal, (representada en cercas vivas, sistemas silvopastoriles, árboles dispersos y áreas de regeneración natural), y con un adecuado manejo agropecuario que permitan la conectividad de los fragmentos de bosques secundarios presentes en la región.
Agradecimientos
Los autores agradecen a la Dirección de Investigación Medellín -DIME y a la Vicerrectoría de investigación de la Universidad Nacional de Colombia por la financiación del proyecto. Al programa Jóvenes Investigadores de Colciencias por el respaldo económico a C.M. Martinez-Bravo, y el apoyo de los estudiantes auxiliares que participaron en la fase de campo de la investigación. Igualmente, a los revisores anónimos por sus pertinentes comentarios para mejorar el manuscrito.
Referencias
Balmer, O. (2001). Species lists in ecology and conservation: Abundances matter. Conservation Biology, 16, 1160-1161. [ Links ]
Barrett, G., Ford, H. A., & Recher, H. F. (1994). Conservation of wood- land birds in a fragmented rural landscape. Pacific Conservation Biology, 1, 245-256. [ Links ]
Bibby, C. J., Jones, M., & Marsden, S. (2000). Expedition Field Techniques Bird Surveys. Expedition Advisory Centre, London. [ Links ]
Bierregaard, R. O. Jr., Lovejoy, T. E., Kapos, V., Dos Santos, A. A., & Hutchings, R. W. (1992). The biological dynamics of tropical rainforest fragments. BioScience, 42, 859-866. [ Links ]
Blendinger, P., & Álvarez, M. E. (2002). Ensambles de aves de los bañados de Carilauquen (Laguna Llancanelo, Mendoza, Argentina): consideraciones para su conservación. El Hornero, 17, 71-83. [ Links ]
Borges, J. C., & López-Mata, L. (2005). Riqueza y diversidad de especies de aves en una selva mediana subperennifolia en el Centro de Veracruz, México. Acta Zoológica Mexicana, 21(1), 1-20. [ Links ]
Brandolin, P., Martori, R., & Avalos, M. (2007). Variaciones temporales de los ensambles de aves de la reserva natural de fauna laguna La Felipa (Córdoba, Argentina). El Hornero, 22(1), 1-8. [ Links ]
Cárdenas, G. (2000). Comparación de la composición y estructura de la avifauna en diferentes sistemas de producción (Tesis de grado). Universidad del Valle, Cali, Colombia. [ Links ]
Castaño, G. J., & Patiño, J. C. (2000). Cambios en la composición de la avifauna en Santa Helena durante el siglo XX. Crónica forestal y del medio ambiente, 15, 137-161. [ Links ]
Castaño, G., Morales, J., & Bedoya-Álvarez, M. (2008).Aportes de una plantación forestal mixta a la conservación de la avifauna en el cañón del río Cauca, Colombia. Revista Facultad Nacional de Agronomía Medellín, 61(1), 4358-4365. [ Links ]
Chaves, M. E., & Arango, N. (1998). Informe Nacional sobre el Estado de la Biodiversidad 1997 Colombia. Santa Fe de Bogotá: Instituto Alexander vonHumboldt, PNUMA, Ministerio de Medio Ambiente. Tomo 1. [ Links ]
Codesido, M., & Bilenca, D. (2004). Variación estacional de un ensamble de aves en un bosque subtropical semiárido del Chaco Argentina. Biotropica, 36(4), 544-554. [ Links ]
Collwell, R. (2006). Statistical Estimation of Species Richness and Shared Species from Samples.Version 8.0 EstimateS. University of Connecticut. USA. [ Links ]
DeClerck, F., Martínez, A., & DeClerck, R. (2011). Aves en cercas vivas. Agroforestería en las Américas, 48, 21-25. [ Links ]
De Los Ríos, J., & Gallego, A. (2003). Caracterización y evaluación de los agroecosistemas de los centros agropecuarios Cotové y Paysandú (Trabajo de grado). Universidad Nacional de Colombia, sede Medellín. Colombia. [ Links ]
Duran, S., & Kattan, G. (2005). A Test of the utility of exotic tree plantation for undestory birds and food resources in the Colombian Andes. Biotropica, 37(1), 129-135. [ Links ]
Etter, A. (1993). Diversidad ecosistémica en Colombia hoy. In: Nuestra diversidad biótica. CEREC y Fundación Alejandro Ángel Escobar. [ Links ] 43-61.
Fandiño, M. C., & Ferreira, P. (1998). Colombia, Biodiversidad Siglo XXI: Propuesta técnica para la formulación de un plan de acción nacional en biodiversidad. Instituto Alexander von Humboldt, Ministerio del Medio Ambiente, DNP, Santa Fe de Bogotá [ Links ].
Fundación Los Yalcones. (2000). Guía de Campo: Aves del Corredor Biológico PNN Puracé PNN Cueva de los Guacharos. San Agustín Colombia: FUNYA, MINAMBIENTE, UAESPNN, CAM. [ Links ]
Gallo-Cajiao, E., & Idrobo-Medina, C. (2004). Fragmentos de bosque y conservación de aves: un estudio de caso en los Andes de Colombia. Memorias: Manejo de Fauna silvestre en Amazonia y Latinoamérica. 178-185. [ Links ]
García, G. O., & Gómez-Laich, A. (2007). Abundancia y Riqueza específica de un ensamble de aves marinas y costeras del sudeste de la provincia de Buenos Aires, Argentina. El Hornero, 22 (1), 9-16. [ Links ]
Greenberg, R., Bichier, P., & Sterling, J. (1997). Acacia, cattle and migratory birds in southeastern Mexico. Biological Conservation, 80, 235-247. [ Links ]
Greenberrg, R., & Marra, P. P. (2005). Birds of two worlds: The ecology and evolution of migration. London: Johns Hopkins University Press. [ Links ]
Gustafson, E. J., Knutson, M. G., Niemi, G. J., & Friberg, M. (2002). Evaluation of spatial models to predict vulnerability of forest birds to brood parasitism by cowbirds. Ecological Applications, 12, 412-426. [ Links ]
Haffer, J. (1967). Zoogeographical notes on the “nonforest” lowland bird faunas of northwestern South America. El Hornero, 10, 315-333. [ Links ]
Harvey, C. A., Guindon C. F., Haber W. A., DeRosier, H., & Murray, K. G. (2000). The importance of forest patches, isolated trees and agricultural windbreaks for local and regional biodiversity: the case of Monte Verde, Costa Rica. XXI IUFRO World Congress, Kuala Lumpur, Malaysia, International Union of Forestry Research Organizations, Sub plenary sessions. 1: 787-798. [ Links ]
Harvey, C. A., Komar, O., Chazdon, R., Ferguson, B. G., Finegan, B., Griffith, D. M., Martínez-Ramos, M., Morales, H., Nigh, R., Soto-Pinto, L., Van Breugel, M., & Wishnie, M. (2008). Integrating Agricultural Landscapes with Biodiversity Conservation in the Mesoamerican Hotspot. Conservation Biology, 22 (1), 8-15. [ Links ]
Herrera, D. (1974). Estudio Preliminar de la Avifauna en Cotové. Seminario Facultad de Agronomía. Universidad Nacional de Colombia, sede Medellín. Colombia. [ Links ]
Herrera, D. (1976). Estudio Preliminar de las Aves en los Alrededores de Santa Fe de Antioquia (Trabajo de grado). Universidad Nacional de Colombia, sede Medellín, Colombia. [ Links ]
Hilty, S. L., & Brown, W. L. (1986). A guide to the birds of Colombia. Princeton: Princeton University Press. [ Links ]
Holdridge, L. R. (1967). Life zone ecology. Costa Rica: Centro Científico Tropical. [ Links ]
IDEAM. (2008). Sistema de información nacional ambiental, Estación 2622503 Cotove Hda. Disponible en http://intranet.ideam.gov.co/Vinfmeteo/Mventaig.asp [ Links ]
Instituto Alexander von Humboldt, IAvH. (1998). El bosque seco tropical (Bs-T) en Colombia. Programa de inventario de la biodiversidad, grupo de exploraciones y monitoreo ambiental GEMA, IAVH, Villa de Leyva. 24 p. [ Links ]
Idrobo-Medina, C. J., & Gallo-Cajiao, E. (2004). Movilidad de aves de sotobosque entre fragmentos de bosque subandino en los Andes colombianos. Memorias de VI Congreso Internacional sobre Manejo de Fauna Silvestre en la Amazonia y Latinoamérica, Iquitos, Perú. Disponible en http://www.revistafauna.com.pe/memo.htm [ Links ]
Johnson, M. D., Sherry, T. W., Holmes, R. T., & Marras, P. P. (2005). Assessing Habitat Qualily for a Migratory Songbird Wintering in Natural and Agricultural Habitats. Conservation Biology, 20(59), 1433-1444. [ Links ]
Karr, J. R., Robinson S. K., Blake J. G., & Bierregaard, R. O. (1990). Birds of four Neotropical forests (pp. 237-269). In A. H. Gentry (Ed.), Four Neotropical rainforests. Yale University Press. [ Links ]
Kattan, G. H. (1992). Rarity and vulnerability: the birds of the Cordillera Central Colombia. Conservation Biology, 6(1), 64-70. [ Links ]
Kattan, G. H., Álvarez-López, H., & Giraldo, M. (1994) .Forest Fragmentation and Bird Extinctions: San Antonio Eighty Years Later. Conservation Biology, 8(1), 138-146. [ Links ]
Kattan, G., & Murcia, C. (2003) .A review and synthesis of conceptual frameworks for the study of forest fragmentation (pp. 183-200). In G. A. Bradshaw & P. A. Marquet (Eds.), How landscapes change. Human disturbance and ecosystem fragmentation in the Americas. New York: Springer-Verlag. [ Links ]
Koleff, P. (2005). Conceptos y medidas de la diversidad beta (pp. 19-40). In G. Halffter, Soberón, J., Koleff, P. & Melic, A. Sobre Diversidad Biológica: El significado de las Diversidades áâϒ. CONABIO, SEA, DIVERSITAS y CONACyT. Volumen 4. Zaragoza: Editorial Monografías Tercer Milenio. [ Links ]
Kupfer, J. A., Malanson, G. P., & Franklin, S. B. (2006). Not seeing the ocean for the islands: the mediating influence of matrix-based processes on forest fragmentation effects. Global Ecology and Biogeography, 15, 8-20. [ Links ]
Kurosawa, R., & Askins, R. A. (2003). Effects of habitat fragmentation on birds in deciduous forests in Japan. Conservation Biology, 17(3), 695-707. [ Links ]
Lara, C. E., Cuervo, A. M., Valderrama, S. V., Calderón-F. D., & Cadena, C. D. (2012). A new species of wren (Troglodytidae: Thryophilus) from the dry Cauca River Canyon, northwestern Colombia. The Auk, 129 (3), 537-550. [ Links ]
Lozano, I. E. (1993). Diversidad y organización en gremios de la comunidad de aves del sotobosque de bosque primario y vegetación secundaria (pp. 141-154). In G. I. Andrade (Ed.), Carpanta, selva nublada y páramo. Colombia: Fundación Natura. [ Links ]
Lynch, J. F. (1989). Distribution of overwintering neartic migrants in the Yucatan Peninsula, II: use of relative and human-modified vegetation (pp. 78-196). In J. M. Hagan, & D. W. Johnston (Eds.), Ecology and Conservation of Neotropical Migrant Land birds. Washington, D. C.: Smithsonian Institution Press. [ Links ]
Magurran, A. E. (1988). Ecological diversity and its measurement. Princeton University Press. [ Links ]
McIntyre, S., & Hobbs, R. (1999). A framework for conceptualizing human effects on landscapes and its relevance to management and research model. Conservation Biology, 13, 1282-1292. [ Links ]
Múnera, E., Carl, B. B., Bolívar, D. M., & Botero, J. A. (2008). Composición y estructura de la avifauna en Diferentes Hábitats en el Departamento de Córdoba, Colombia (pp. 205-226). In E. Murgueitio, C. Cuartas, & J. Naranjo (Eds.), Ganadería del futuro: Investigación para el desarrollo. Cali, Colombia: Fundación CIPAV. [ Links ]
Naranjo, L. G., Amaya, J. D., Eusse-González, D., & Cifuentes-Sarmiento, Y. (Ed.). (2012). Guía de las Especies Migratorias de la Biodiversidad en Colombia. Aves. Vol. 1. Bogotá, D.C. Colombia: Ministerio de Ambiente y Desarrollo Sostenible/WWF Colombia. [ Links ]
Naranjo, L. G. (2001). Sistemas agroforestales para la producción pecuaria y la conservación de la biodiversidad (pp. 13-25). In Agroforestería para la producción animal en América Latina. Memorias de la segunda conferencia electrónica. Estudio FAO. Producción y sanidad animal 155. [ Links ]
Naranjo, L. G. (1992). Estructura de la avifauna en un área ganadera en el Valle del Cauca, Colombia. Caldasia, 17(1), 55-68. [ Links ]
Norton, M. R., Hannon, S. J., & Schmiegelow, F. K. A. (2000). Fragments are not islands: patch vs. landscape perspectives on songbird presence and abundance in a harvested boreal forest. Ecography, 23, 209-223. [ Links ]
Pearce-Higgins, J. W. (2000). The avian community structure of a Bolivian savanna on the edge of the Cerrado system. El Hornero, 15, 77-84. [ Links ]
Petit, L. J., Petit, D. R., Christian, D. G., & Powell, H. D. W. (1999). Bird communities of natural and modified habitats in Panama. Ecography, 22, 292-304. [ Links ]
Pezo, D., & Ibrahim, M. (1998). Sistemas silvopastoriles. CATIE, Proyecto Agroforestal CATIE/GTZ, Turrialba, Costa Rica. [ Links ]
Primack, R. B. (2002). Essentials of Conservation Biology. 3 ed. Sinauer Assoc., Inc., Sunderland, USA. [ Links ]
Ralph, C. J., Geupe, G. R., Pyle, P., Martín, T. E., & DeSante, D. F. (1993). Handbook of field methods for monitoring land birds. U.S. Department of Agriculture, Forest Service General Technical Report. PSW-GTR-144. USDA. Albany, CA. [ Links ]
Ramírez-Albores, J. E. (2006).Variación en la composición de comunidades de aves en la Reserva de la Biosfera Montes Azules y áreas adyacentes, Chiapas, México. Biota Neotropical, 6(2), 1-19. [ Links ]
Ramo, C., & Busto, B. (1984). Nidificación de los Passeriformes en los llanos de Apure, Venezuela. Biotropica, 16(1), 59-68. [ Links ]
Remsen, J. V. Jr., Cadena, C. D., Jaramillo, A., Nores, M., Pacheco, J. F., Robbins, M. B., Schulenberg, T. S., Stiles, F. G., Stotz, D. F., & Zimmer, K. J. (2011). A classification of the bird species of South America. American Ornithologists’ Union. Disponible en: http://www.museum.lsu.edu/~Remsen/SACCBaseline.html. [ Links ]
Remsen, J. V. (1994). Use and misuse of bird lists in community ecology and conservatio0n. The Auk, 111(1), 225-227. [ Links ]
Renjifo, L. (2001). Effect of natural and anthropogenic landscape matrices on the abundant of subandean birds species. Ecological Applications, 11(1), 14-31. [ Links ]
Restall, R., Rodner, C., & Lentino, M. (2007). Birds of Northern South America: An Identification Guide, Volume 1 y 2. U.S.: Yale University Press. [ Links ]
Ricketts, T. H. (2001). The matrix matters: effective isolation in fragmented landscapes. The American naturalists, 158(1), 87-99. [ Links ]
Rivera-Gutiérrez, F. (2006). Composición y estructura de una comunidad de aves en un área suburbana en el suroccidente Colombiano. Ornitología Colombiana, 4, 28-38. [ Links ]
Robinson, W. (1999). Long-term changes in the avifauna of Barro Colorado island, Panamá, a tropical forest isolate. Conservation Biology, 13(1), 85-97. [ Links ]
Salaman, P., Donegan, T., & Caro, D. (2008). Listado de avifauna colombiana 2007. Conservación Colombiana 2 Suplemento. 85. [ Links ]
SAS/STAT. (2001). User´s Guide (Relase 8.0), Cary, NC, USA: SAS inst. Inc. [ Links ]
Shmida, A. V. I., & Wilson, M. V. (1985). Biological determinants of species diversity. Journal of Biogeography, 12, 1-20. [ Links ]
Stephens, S. E., Koons, D. N., Rotella, J. J., & Willey, D. W. (2003). Effects of habitat fragmentation on avian nesting success: a review of the evidence at multiple spatial scales. Biological Conservation, 115, 101-110. [ Links ]
Stiles, F. G. (1998). Inventario de las aves de un bosque altoandino: Comparación de dos métodos. Caldasia, 20, 29-43. [ Links ]
Stiles, F. G., & Bohórquez, C. I. (2000). Evaluando el estado de la biodiversidad: el caso de la avifauna de la Serranía de las Quichas, Boyacá, Colombia. Caldasia, 22(1), 61-92. [ Links ]
Stotz, D. F., Parker, T. A., Fitzpatrick, J. W., & Moskovits, D. K. (1996). Neotropical birds: Ecology and Conservation. Chicago: University Press. [ Links ]
Verea, C., Fernández-Badillo, A., & Solorzano, A. (2000). Variación en la composición de las comunidades de aves de sotobosque de dos bosques en el norte de Venezuela. Ornitología Neotropical, 11, 65-79. [ Links ]
Waltert, M., Mardiastuti, Ani., & Muhlengerg, M. (2004). Efectos del uso del suelo sobre la riqueza de especies de aves en Sulawesi, Indonesia. Conservation Biology, 18 (5), 1339-1346. [ Links ]
*Correspondencia a:
1Caty Milena Martínez-Bravo. Postgrado en Bosques y Conservación Ambiental, Universidad Nacional de Colombia, Sede Medellín, Colombia; catymilenam@yahoo.es
2 Néstor Javier Mancera-Rodríguez. Universidad Nacional de Colombia, Departamento de Ciencias Forestales, Sede Medellín, Calle 59A No. 63-20, Bloque 20, oficina 21, Medellín, Colombia; njmancer@unal.edu.co
3Germán Buitrago-Franco. Grupo Ecología y Conservación de Fauna Silvestre. Departamento de Ciencias Forestales, Sede Medellín, Calle 59A No. 63-20; gabuitra@gmail.com
Recibido 28-XI-2012. Corregido 20-VI-2013. Aceptado 22-VII-2013


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}