










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
Trophic ecology of the exotic Lerma livebearer Poeciliopsis infans (Cyprinodontiformes: Poeciliidae) in the Lago de Pátzcuaro, Central Mexico
Ecología trófica del pez exótico Guatapote del Lerma Poeciliopsis infans (Cyprinodontiformes: Poeciliidae) en el lago de Pátzcuaro, Región Central de México
Ecología trófica del pez exótico Guatapote del Lerma Poeciliopsis infans (Cyprinodontiformes: Poeciliidae) en el lago de Pátzcuaro, Región Central de México
Juan P. Ramírez-Herrejón1*, Lissette S. Castañeda-Sam2*, Rodrigo Moncayo-Estrada3*, Javier Caraveo-Patiño1 & Eduardo F. Balart1
*Dirección para correspondencia:
Abstract
Exotic fish species has caused several impacts on aquatic biodiversity. The Lago de Pátzcuaro has some well-studied exotic species, except the Lerma livebearer Poeciliopsis infans. This fish species was introduced into the Lago de Pátzcuaro before 1997 and the aspects of its biology are still unknown. In this study we assessed aspects of the trophic ecology of this exotic fish, P. infans, using gut content and stable isotope analysis to understand its capacity to tolerate anthropogenic environmental degradation in the Lago de Pátzcuaro. We also determined its trophic guild position (TP) using the TrophLab Program and stable isotope. Niche breadth was calculated by standardized Levins’ Index (Bi). Fish was captured with a seine during wet and dry seasons at six environmentally different sites and gut contents were obtained. We analyzed a total of 239 gut contents of P. infans. The contribution of each food item in the diet was quantified using frequency of occurrence and area percentage. The importance of each prey item was determined according to the index of relative importance (IRI), and the omnivory index (OI) was used to assess the feeding behavior. Fish were categorized by size and the diet was compared between fish sizes and sites. Dorsal muscle tissue and water hyacinth tissue was obtained for nitrogen isotope signature (δ15N) analysis. Additionally, we measured water and habitat quality to evaluate environmental conditions at each site. We concluded that P. infans is an omnivore (OI=0.28) that consumes mainly detritus (44%), epiphytic diatoms (37%), and secondary on terrestrial insects (6%) and zooplankton (10%). The fish can behave as a specialist (Bi=0.39) or generalist (Bi=0.68) and as a primary consumer (TROPH=2.2; TP=2.3) with a feeding strategy that was the same at different sizes, seasons and sites. None of the evaluated sites showed good environmental quality. We argue that P. infans can tolerate changes in water quality and feeding items availability, because it can exploit resources in multiple trophic webs. However, this species could be dependent on habitat complexity, especially in the aquatic vegetation cover.
Key words: shallow lake, introduced species, tolerant species, ecological strategy, trophic ecology.
Resumen
Las especies de peces exóticas han causado numerosos impactos sobre la biodiversidad acuática. El Lago de Pátzcuaro tiene especies exóticas bien estudiadas, excepto el Guatapote del Lerma, Poeciliopsis infans. Esta especie fue introducida en el Lago de Pátzcuaro antes de 1997 y los aspectos de su biología son aún desconocidos. Se estudiaron aspectos de la ecología trófica de este pez exótico, P. infans, mediante el uso de análisis de contenidos del tracto digestivo para entender su capacidad para tolerar la degradación ambiental antrópica en el Lago de Pátzcuaro. Lo que involucró la determinación del gremio y posición trófica (TP) con el programa TrophLab e isotopos estables. La amplitud de dieta fue calculada con el índice estandarizado de Levin (Bi). Un total de 239 tractos digestivos de P.infans fueron analizados. Los peces fueron capturados con una red tipo chinchorro durante la temporada de lluvias y estiaje en seis sitios ambientalmente distintos. La contribución de cada componente de la dieta fue cuantificada con la frecuencia de ocurrencia y el porcentaje de área. La importancia relativa de cada artículo alimentario fue determinada de acuerdo al índice de importancia relativa (IIR) y el índice de omnivoría (OI) fue usado para estimar la conducta en la forma de alimentarse. La dieta fue comparada entre tallas y entre sitios de estudio. El tejido de músculo dorsal y el tejido de lirio fueron obtenidos para determinar valores de δ15N. De manera adicional, la calidad del agua y del hábitat fue medida para evaluar la condición ambiental en cada sitio. P. infans es un omnívoro (OI=0.28) que consume principalmente detritus (44%), diatomeas epifíticas (37%) y de forma secundaria insectos terrestres (6%) y zooplancton (10%); puede comportarse como especialista (Bi=0.39) o generalista (Bi=0.68) y consumidor primario (TROPH=2.2; TP=2.3). La estrategia alimenticia fue similar entre tallas, temporadas y sitios. Ningún sitio mostró buena calidad ambiental. Los resultados del presente estudio permiten argumentar que P. infans puede tolerar cambios en la calidad del agua y en la disponibilidad de artículos alimenticios, porque puede usar recursos provenientes de múltiples redes tróficas. Sin embargo, esta especie podría ser dependiente de la complejidad del hábitat, especialmente de la cobertura de la vegetación acuática.
Palabras clave: lago somero, especies introducidas, especies tolerantes, estrategia ecológica, ecología trófica.
The introduction of exotic fish species has been increasing and is the second most important factor of human impact on aquatic biodiversity (Koehn 2004). The Lerma livebearer Poeciliopsis infans (Woolman 1894) is a fish endemic to the Lerma-Grande de Santiago river basin and tributaries of the Ameca, Armería, Coahuayana and Balsas rivers in Mexico (Miller et al. 2009). This species has the ability to inhabit stagnant and shallow water near the shore in lakes, rivers, streams, springs, ponds, and channels (Miller et al. 2009). P. infans is an introduced species in Lago de Pátzcuaro (Galindo-Villegas & Sosa-Lima 2002) and is considered a tolerant species, because it inhabits freshwater bodies with high levels of environmental degradation (Lyons et al. 2000, Mercado-Silva et al. 2002).
The assessment of the feeding ecology of introduced fish, contributes to understand its survival strategy in novel ecosystems. This omnivorous fish feeds on detritus of plant and animal origin, which can confer resistance, as the base of the food web is altered and trophic levels are simplified (Karr 1981, Vanni et al. 2005). Currently, its trophic ecology has barely been studied. Some authors argue that the species is an herbivore or omnivore (Lyons et al. 1995, Lyons et al. 2000, Mercado-Silva et al. 2002). Studies at Lago de Cuitzeo (Zubieta 1985), Guaracha Reservoir (Escalera-Gallardo 1986) in the state of Michoacán, and at San Miguel Arco Reservoir in the state of México (Navarrete et al. 2008) consider this species an omnivore.
The first published record of P. infans in Lago de Pátzcuaro was in 1997 (Galindo-Villegas & Sosa-Lima 2002), where it is found throughout the lake (Mar-Silva 2011). Lago de Pátzcuaro offers a study model to investigate trophic ecology of this species, particularly when present in exotic environmental conditions. This lake has habitats with different degrees of environmental degradation (Bernal-Brooks et al. 2002, Orbe-Mendoza et al. 2002, Berry et al. 2011); physical and chemical water properties (Alcocer & Bernal-Brooks 2002), and habitat characteristics (Mar-Silva 2011, Vital-Rodríguez 2011).
This study assessed the trophic guild, niche breadth, and trophic level of P. infans to understand its feeding strategy and ability to tolerate variations in the quality of environmental conditions in Lago de Pátzcuaro.
Materials and Methods
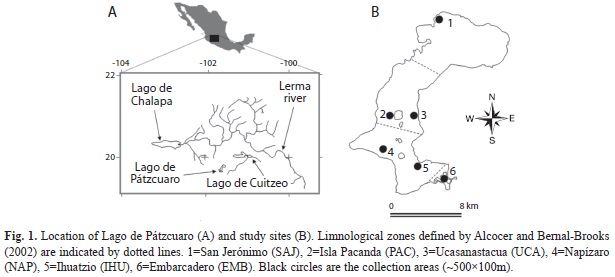
Study area: Lago de Pátzcuaro is in the North-central part of the State of Michoacán, Mexico (19°35’ N - 101°40’ W) at ~2 035m elevation (Fig. 1), with an area of 130km2, including its islands (Bernal-Brooks et al. 2002).
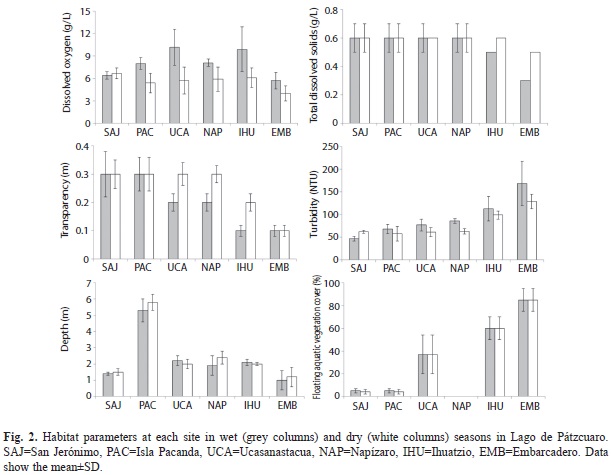
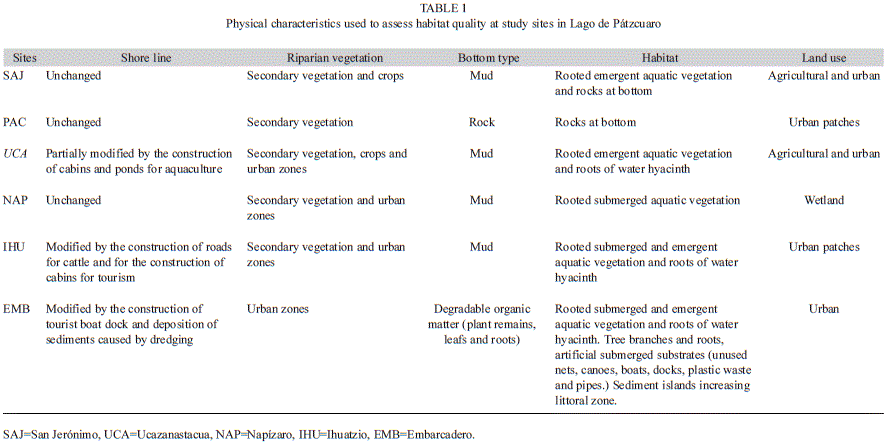
Samples were collected during the wet season (September and November 2009) and dry season (February and June 2010) at six sites (once a month) around the lake: San Jerónimo (SAJ; 19°40’40.4’’ N - 101°36’16.9’’ W), La Pacanda (PAC; 19°36’38.1’’ N - 101°39’2.7’’ W), Ucazanastacua (UCA; 19°35’51.1’’ N - 101°37’58.5’’ W), Napízaro (NAP; 19°35’20.8’’ N - 101°40’12.7’’ W), Ihuatzio (IHU; 19°35’35.5’’ N - 101°40’45.2’’ W), and Embarcadero (EMB; 19°33’0.6’’ N - 101°37’30.7’’ W). The physical and chemical water characteristics and habitat characteristics at these sites are shown in figure 2 and table 1.
Environmental quality: We measured environmental quality at each site; we considered both water and habitat quality, according to qualitative and quantitative criteria proposed by Lyons et al. (1995) in lotic systems, and applied by Medina-Nava (2003) in lakes. The water quality, habitat quality and environmental quality gradient used values from 0 to 100, where high quality sites are close to 100. Water quality variables (pH, dissolved oxygen, total dissolved solids, and turbidity) were measured with a multiparameter equipment (Hydrolab Quanta multimeter, Hach, Loveland, CO; Fig. 2). We look for visual evidence of oils and detergents on the water surface, following Lyons et al. (1995). Habitat quality variables measured were: Secchi disk transparency, depth, bottom type, shoreline condition, aquatic and riparian vegetation, habitat abundance and availability, and land use near the site (Fig. 2, Table 1). Floating aquatic vegetation (water hyacinth Eichornia crassipes) coverage was estimated using a visual technique proposed by Barbour et al. (1999). Sediment samples were collected with an Ekman dredge; the bottom type was described (Bouyoucos 1936).
Fish sampling and trophic analysis: Fish were captured during the day (08:00-13:00) with a seine net (75m long, 5m wide, 1cm mesh size). Specimens were labeled and transported on ice to stop digestion (Caillet et al. 1996). Accumulated prey diversity was measured, using Simpson’s index to determine the minimum number of stomachs to characterize the feeding habits of P. infans (Magurran 2004). The curve became asymptotic at 23 individuals. Standard length (SL), weight, and gastric repletion for each individual were measured. A modified version of the quadrant method (Hynes 1950) for gut content analysis was used.
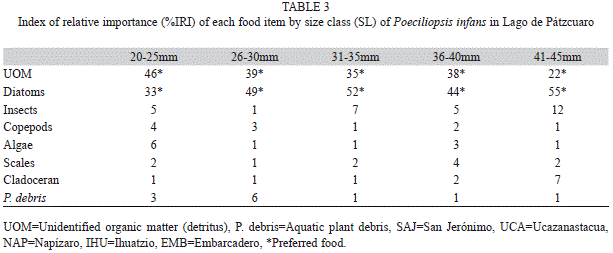
Prey items were identified to equivalent taxa level (Edmondson 1959, Pennak 1978). We mostly found remains of insects and the identification of insects until the taxonomical level of order was not possible. A modified version of the Index of Relative Importance (IRI) proposed by Yañez-Arancibia et al. (1976) was used: IRI=(F×A)/100, where, F is the frequency of occurrence, and A is the area. This method is used when the gut content is constituted by small feeding components (diatoms, copepods, ostracods, rotifers, cladocerans) or when its quantification is not possible (detritus, vegetation debris) (Vega-Cendejas 1990, Canto-Maza & Vega-Cendejas 2008). IRI is expressed as percentages (Cortés 1997) to describe importance of prey. Individuals were categorized in 5mm intervals with respective IRI to determine a probable diet change as growth increases.
The standardized Levins’ Index (Bi) was used to calculate niche breadth, (values from 0 to 1). Values of Bi<0.60 were considered specialists and values of Bi>0.60 were considered generalists (Krebs 1989). Feeding behavior was described by the Omnivory Index (OI); it was calculated as the variance of the trophic levels of a consumer’s preys (Christensen & Pauly 1992). The trophic level of P. infans was estimated using the TrophLab Program (Pauly et al. 2000). It includes the number of groups in the diet, the prey fraction of the diet (IRI, in %), and the trophic level of the prey. Some studies have argued that gut content analysis is biased because it cannot consider assimilated food (Bearhop et al. 2004). δ15N is an accurate indicator of the fish trophic position because it reflects assimilated food and show progressive enrichment (3‰ to 5‰) from feeding components (Jardine et al. 2003). For this reason, the trophic position of P. infans was corroborated with nitrogen stable isotope analysis. Fish and water hyacinth samples were obtained for stable isotope analysis during dry and wet seasons. Approximately 0.5g of dorsal muscle tissue was obtained from fish (>30mm of standard length) and frozen for later isotope analysis. Also, approximately 1g of water hyacinth root tissue was obtained to represent primary productivity in the analysis because this aquatic vegetation represents one of the main components of periphyton.
Stable isotope was analyzed by continuous flow isotope ratio mass spectrometer at the University of California-Davis Stable Isotope Facility. Nitrogen stable isotope ratios (δ15N) are expressed in delta (δ) notation and parts per thousand (‰). Mean standard error is <0.1‰ for δ15N. We estimated the trophic position of P. infans using δ15N (Vander Zanden & Rasmussen 1999): TP=(δ15NS-δ15NPP/ΔTP)+1, where TP is the trophic position; δ15NS is the value of the fish tissue, and δ15NPP is the value of primary productivity. We used 3.4‰ as trophic level enrichment (ΔTP), as reported by Vander Zanden et al. (2003).
The importance of food items along the environmental gradient was explored by a multivariate approach with permutation techniques (Mielke & Iyer 1982, Biondini et al. 1988). IRI values of each combination of sites and seasons were considered as the units of comparison; input data were logarithmically transformed. The hypothesis of no diet differences between sites were tested using the nonmetric MRPP (Multi-response Permutation Procedures) (McCune & Grace 2002). MRPP was performed using PC-ORD 5.07 (McCune & Mefford 2006).
Non-parametric Wilcoxon rank sums-test (Wilcoxon 1945) was used to detect differences of δ15N values and trophic position among sites during the wet and dry seasons. If significant differences were found, multiple comparisons were made using the Tukey-Kramer honestly significant difference (HSD) post hoc test (Zar 1999). Both analyses were performed with JMP 3.1.6.2 software (SAS Institute 1995).
Results
We analyzed 239 specimens from 20-45mm standard length, 100 in the wet season and 139 in the dry season, about 90% of the obtained digestive tracts (216) were at least 50% full, ensuring a good description of the diet.
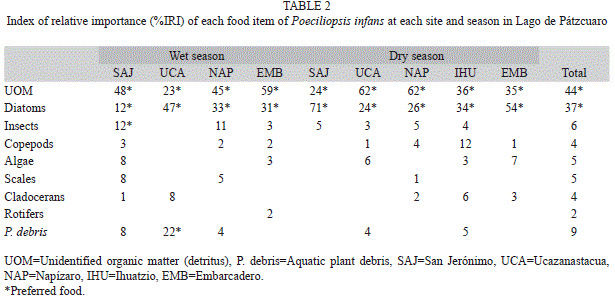
We found nine food items of which the detritus and diatoms recorded were the maximum values considered as food preferred during both seasons, the remaining components were considered as secondary or incidental (Table 2). For the site EMB during wet season and NAP, UCA during dry season, the detritus recorded its maximum value (59% and 62%, respectively). While diatoms recorded its highest value in the UCA site during wet season (47%) and the SAJ site during dry season (71%). Fish length did not show significant differences in diet between seasons or sites (A=0.02, p=0.95). No significant differences in food items were found between seasons (A=0.01, p=1) and sites (A=0.03, p=0.96). However, a pattern of less algae and more insects and macrophytes were found as fish length increased (Table 3).
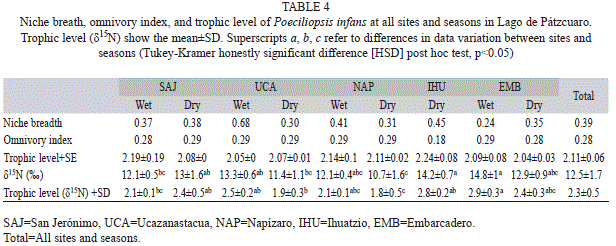
Poeciliopsis infans in Lago de Pátzcuaro was determined to be omnivorous, with a tendency to principally consume benthic and periphytic diatoms and detritus, according to the IRI and the OI (0.28±0.04) (Table 4). Levins’ niche breadth for all sites and both seasons combined was 0.39, which categorizes the species as a specialist (Table 4). However, the values varied according to site and season; at EMB, P. infans behaved as a specialist during the dry season; only at UCA during the wet season, it was a generalist.
The δ15N mean of water hyacinth root tissue was 8.2±0.3‰. The δ15N mean of P. infans for all sites and both seasons and per site are shown in Table 4. We found differences in δ15N values of muscle and trophic positions between sites in wet season (p<0.01, DF=3, χ2=13.09) and dry season (p<0.01, DF=4, χ2=14.14). P. infans was placed between level 2 and 3 at all sites in both seasons, according to the TROPH Program (2.22±0.17) and δ15N analysis (2.3±0.04). This indicates that P. infans is part of the guild of primary consumers (Table 4).
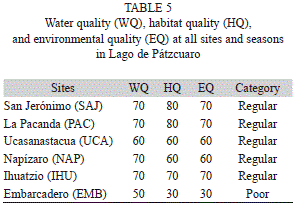
Water quality ranged from 50 to 70; habitat quality ranged between 30 and 80 and environmental quality between 30 and 70. Poor environmental quality (0-40) was found only in the EMB site and the remaining sites had regular environmental quality (50-70). No site had good environmental quality (80-100) (Table 5). At SAJ and PAC, the bottoms were most rock (>90%), at UCA, NAP and IHU, mostly mud (>90%); at EMB, mostly decaying plant remains (>90%).
Discussion
Although P. infans is tolerant of environmental degradation (Lyons et al. 2000, Mercado-Silva et al. 2002), its distribution could be conditioned by habitat quality because at sites that lacked aquatic vegetation, not enough fishes were captured to make accurate trophic analysis. We hypothesized that the distribution of P. infans in Lago de Pátzcuaro is related to cover with floating aquatic vegetation. In part, this habitat offers refuge to avoid predation (Lampert & Sommer 2007). Otherwise, Poeciliopsis infans behaves as a grazer associated with the periphytic and benthic zones, with the capability of feeding on terrestrial insects and aquatic larvae in the benthic zone. We hypothesized that P. infans can tolerate water quality changes in Lago de Pátzcuaro because it feeds on detritus and diatoms from diverse trophicwebs such as benthos and periphyton. These items are abundant resources in freshwater systems at all levels of environmental degradation (Stevenson & Smol 2002).
Omnivory is defined as feeding on more than one trophic level (Pimm 1982, Fagan 1997). Our data showed that Poeciliopsis infans is an omnivore species in Lago de Pátzcuaro. Dietary contributions were similar to those found for populations in other water bodies. In Lago de Cuitzeo (Zubieta 1985) and Guaracha Reservoir (Escalera-Gallardo 1986) in the state of Michoacán, Mexico, detritus and diatoms represented up to 40% of diet volume; in the reservoirs of San Miguel Arco in the state of Mexico, cyanobacteria and filamentous algae were numerically important (76%), while chironomids and cladocerans were volumetrically important (82%) (Navarrete et al. 2008). Such differences could be related to habitat complexity that favored specific feeding resources (Post et al. 2000).
P. infans in Lago de Pátzcuaro feeds on specific prey from different trophic webs periphytic diatoms (littoral trophic web), cladocerans and secondary consumers like copepods (limnetic trophic web) and terrestrial adult insects (terrestrial trophic web).This behavior is described by Vadeboncoeur et al. (2005) as multi-chain omnivores for generalist predators that exploit food chains of different primary-producer functional groups, including detritus. Similar information was reported for Lago de Cuitzeo where stomach contents were 11% terrestrial insects (Duarte 1981) and 67% for San Miguel Arco Reservoir (Armendáriz et al. 2008).
The results of Levins’ index classified P. infans as a specialist because it preferred high abundance of detritus and periphytic diatoms in the lake, which indicates the capability of this species in particular, and poecilids in general, to resist fluctuations in availability of diet (Pollux & Reznick 2011). This fish could be also considered as an opportunist species, however, it is necessary to compare the proportion of prey taxon in the diet and the proportion of prey taxon in the lake (Rachlin et al. 1989).
Compared to another poecilid, such as Gambusia holbrooki, P. infans can be also opportunistic because it consumes insects attached to emergent aquatic vegetation (King & Warburton 2007). Insect predation is reported in introduced poecilids, such as G. holbrooki, which imposes negative competitive effects on native fish species (Schaefer et al. 1994). At EMB, water hyacinth (Eichornia crassipes) covering >90%, could serves as refuge and a habitat with food, because insects and diatoms consumed by fish reside on the roots of this plant; this is consistent with Toft et al. (2003), who found the same function of water hyacinth in the Sacramento River Delta in California. In addition, the large volume of detritus in the gut could indicate the species adaptability, where eutrophication is present. According to Rosenberger et al. (2008), nutrient inputs have significant effects on the nearshore periphyton community composition, which increases productivity.
Although our results describe P. infans as omnivore as well as specialist, the second characteristic of this species can be not related with trophic habitat or diet limitation; on the contrary, this feeding behavior could be associated with the eutrophic condition of the ecosystem. Because, in a eutrophic lake the detritus and algae are ecosystems elements widely abundant (Lampert & Sommer 2007) and P. infans has the capability to eat this resources. This agrees with Trujillo-Jiménez & Toledo-Beto (2007) who mentioned that poeciliids exhibit great plasticity in food habits, they can occupy different trophic levels and guilds, from detritivorous to carnivorous. Ruehl & DeWitt (2005) argued that trophic plasticity of G. affinis can be an approach for resisting environmental variation. Our results also agree with Gkenas et al. (2012) who found that G. holbrooki showed adaptation to prey availability by specialist feeding strategy in a small shallow eutrophic Mediterranean Lake (Lake Pamvotis).
The TROPH and δ15N analyses indicate that the P. infans food is mainly from the primary productivity level, although it feds on other primary consumers and detritus. This differs from Mercado-Silva et al. (2009) who state that P. infans is in the guild of secondary consumers, with values >3‰ of δ15N in river and reservoir ecosystems at Río La Laja basin in the state of Guanajuato, Mexico.
P. infans represents an inconspicuous species that inhabits aquatic vegetation in littoral zones and in the case of Lago de Pátzcuaro, where environmental degradation is evident, P. infans has entered habitats with contrasting environmental conditions. This species, like other introduced poeciliids (P. gracilis, G. affinis, Poecilia latipinna, P. reticulata, Heterandria bimaculata and Xiphophorus hellerii), are tolerant to environmental variability and lower water quality (Mercado-Silva et al. 2002, Varela-Romero et al. 2002, Gómez- Márquez et al. 2008).
Mar-Silva (2011) argued that the density of P. infans is lower at sites with <50% of floating aquatic vegetation cover in Lago de Pátzcuaro. He collected individuals of P. infans in the same six sites of the present study, capturing 7±8 fishes (0.02±0.007 individuals/m2) at SAJ; 4±4 fishes (0.006±0.006 individuals/m2) at PAC; 7±8 fishes (0.01±0.01 individuals/m2) at UCA; 42±54 fishes (0.08±0.07 individuals/ m2) at NAP; 51±144 (0.1±0.1 individuals/m2) at IHU; and 214±206 fishes (0.5±0 individuals/ m2) at EMB. This author argued that the density and biomass of P. infans at Lago de Pátzcuaro is directly related to surfaces covered with floating aquatic vegetation. This suggests that P. infans can be restricted to areas with vegetation cover. According to Mercado-Silva et al. (2009), who collected this species only along the protected (vegetation cover) habitats in reservoirs and river sites in La Laja river basin.
Understanding the strategies and habitat tolerance of fish to environmental changes is essential to build evaluation and monitoring protocols for aquatic ecosystems. We suggest a P. infans continuous monitoring to assess its role and potential impact on this lake (Galindo-Villegas & Sosa-Lima 2002). This will provide elements for management strategies and policies for conservation and rehabilitation.
Acknowledgments
The authors thank members of the Aquatic Biology Laboratory “Javier Alvarado Díaz” at the Universidad Michoacana; M.M. Herrejón Almanza and J.J. Ramírez Becerra for logistical support; R. Quirino, B. Quirino and A. Quirino for field work; and V. Mar Silva, A. Torres Téllez, B. Vital Rodríguez, R. Alvarado, R. Ortega for assistance with lab work. I. Fogel of CIBNOR provided essential editorial services, C. Silva-Bejarano for technical support. This work was funded by Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO grant GN049). J.P.R.H. received support via fellowship (No. 337465/229677) from the Consejo Nacional de Ciencia y Tecnología of Mexico. R.M.E. is a fellow of COFAA-IPN and EDI-IPN.
References
Alcocer, J. & F.W. Bernal-Brooks. 2002. Spatial and temporal heterogeneity of physical and chemical variables for an endorheic, shallow water body: Lake Pátzcuaro, México. Arch. Hydrobiol. 155: 239-253. [ Links ]
Armendáriz, M.A., N.E. Navarrete, E. Fernández, G. Vázquez & E.S. Urrieta. 2008. Relaciones tróficas de los peces del embalse San Miguel Arco, de Soyaniquilpan, Estado de México. Rev. Chapingo Ser. Cienc. 14: 33-38. [ Links ]
Barbour, M.T., J. Gerritsen, B. Zinder & J. Stribling. 1999. Rapid bioassessment protocols for use in streams and wadeable rivers: periphyton, benthic macroinvertebrates and fish. U.S. Environmental Protection Agency, Washington, D.C., USA. [ Links ]
Bearhop, S., C.E. Adams, S. Waldron, R.A. Fuller & H. Macleod. 2004. Determining trophic niche width: a novel approach using stable isotope analysis. J. Anim. Ecol. 73: 1007-1012. [ Links ]
Bernal-Brooks, F., T. Gómez & J. Alcocer. 2002. Lake Pátzcuaro (Mexico): a controversy about the ecosystem water regimen approached by field references, climatic variables, and GIS. Hydrobiologia 467: 187-197. [ Links ]
Berry, J., E. Lee, K. Walton, A. Wilson & F. Bernal-Brooks. 2011. Microcystin production by a persistent cyanobacterial bloom in Lago de Pátzcuaro (Michoacán, Mexico), and apparent bioaccumulation of the toxin in small commercial catches of fish. Environ. Toxicol. Chem. 30: 1621-1628. [ Links ]
Biondini, M., P. Mielke & K. Berry. 1988. Data-dependent permutation techniques for the analysis of ecological data. Vegetatio 75: 161-168. [ Links ]
Bouyoucos, G.J. 1936. Directions for making mechanical analyses of soils by the hydrometer method. Soil Sci. 42: 225-30. [ Links ]
Caillet, G., M. Love & A. Ebeling. 1996. Fishes a Field and Laboratory Manual on Their Structure, Identification and Natural History. Wadsworth, Belmont, CA, USA. [ Links ]
Canto-Maza, W. & M.E. Vega-Cendejas. 2008. Hábitos alimenticios del pez Lagodon rhomboides (Perciformes: Sparidae) en la laguna costera de Chelem, Yucatán, México. Rev. Biol. Trop. 56: 1837-1846. [ Links ]
Christensen, V. & D. Pauly. 1992. ECOPATH II a software for balancing steady-state ecosystem models and calculating network characteristics. Ecol. Model. 61: 169-185. [ Links ]
Cortés, E. 1997. A critical review of methods of studying fish feeding based on analysis of stomach contents: application to elasmobranch fishes. Can. J. Fish. Aquat. Sci. 54: 726-738. [ Links ]
Duarte, m. 1981. Contribución al conocimiento de los hábitos alimentarios de doce especies de peces en el lago de Cuitzeo, Michoacán. Tesis de Licenciatura, Instituto Politécnico Nacional, México, D.F., México. [ Links ]
Edmondson, T. 1959. Freshwater Biology. Wiley & Sons, New Jersey, USA. [ Links ]
Escalera-Gallardo, C. 1986. Alimentación de los peces en la presa Guaracha, Michoacán. Informe TécnicoNo. 22. CIIDIR-IPN-MICHOACÁN, Michoacán, Mexico. [ Links ]
Fagan, F. 1997. Omnivory as a stabilizing feature of natural communities. Am. Nat. 150: 554-567. [ Links ]
Galindo-Villegas, J. & E. Sosa-Lima. 2002. Gonopodial system review and a new fish record of Poeciliopsis infans Cyprinodontiformes: Poeciliidae) for Lake Pátzcuaro, Michoacán, Central Mexico. Rev. Biol. Trop. 50: 1151-1157. [ Links ]
Gkenas, C., A. Oikonomou, A. Economou, F. Kiosse & I. Leonardos. 2012. Life history pattern and feeding habits of the invasive mosquitofish, Gambusia holbrooki, in Lake Pamvotis (NW Greece). J. Biol. Res.- Thessalon. 17: 121-136. [ Links ]
Gómez-Márquez, J., B. Peña-Mendoza, I. Salgado-Ugarte, A. Sánchez-Herrera, L. Sastré-Baez. 2008. Reproduction of the fish Poeciliopsis gracilis (Cyprinodontiformes: Poeciliidae) in Coatetelco, a tropical shallow lake in Mexico. Rev. Biol. Trop. 56: 1801-1812. [ Links ]
Hynes, B. 1950. The food of fresh-water sticklebacks (Gasterosteus aculeatus and Pygosteus pungitius), with a review of methods used in studies of the food of fishes. J. Anim. Ecol. 19: 36-57. [ Links ]
Jardine, T.D., S. McGeachy, C. Paton, M. Savoie & R. Cunjak. 2003. Stable isotopes in aquatic systems: Sample preparation, analysis, and interpretation. Canadian Manuscript Report of Fisheries and Aquatic Sciences No 2656. Fisheries and Oceans Canada, Ontario, Canada. [ Links ]
Karr, J.R. 1981. Assessment of biotic integrity using fish communities. Fisheries 6: 21-27. [ Links ]
King, S. & K. Warburton. 2007. The environmental preferences of three species of Australian freshwater fish in relation to the effects of riparian degradation.Environ. Biol. Fish. 78: 307-316. [ Links ]
Koehn, J.D. 2004. Carp (Cyprinus carpio) as a powerful invader in Australia waterways. Freshwater Biol. 49: 882-894. [ Links ]
Krebs, J. 1989. Ecological Methodology. Harper & Row, New York, USA. [ Links ]
Lampert, W. & U. Sommer. 2007. Limnoecology. Oxford University, Oxford, England. [ Links ]
Lyons, J., S. Navarro-Pérez, A.P. Cochran, C.E. Santana & M. Guzmán-Arroyo. 1995. Index of biotic integrity based on fish assemblages for the conservation of streams and rivers in west-central México. Conserv. Biol. 3: 569-584. [ Links ]
Lyons, J., A. Gutierrez-Hernandez, E. Diaz-Pardo, E. Soto-Galera, M. Medina-Nava & R. Pineda-López. 2000. Development of a preliminary index of biotic integrity (IBI) based on fish assemblages to assess ecosystem condition in the lakes of central Mexico. Hydrobiologia 418: 57-72. [ Links ]
Magurran, A. 2004. Measuring biological diversity. Blackwell, London, England. [ Links ]
Mar-Silva, V. 2011. Variación espacio-temporal de la comunidad de peces en el lago de Pátzcuaro, Michoacán, México. Tesis de Licenciatura, Universidad Michoacana de San Nicolás de Hidalgo, Michoacán, Mexico. [ Links ]
McCune, B. & J. Grace. 2002. MRPP (multi-response permutation procedures) in analysis of ecological communities. MjM Software Design, Oregon, USA. [ Links ]
McCune, B. & J. Mefford. 2006. PC-ORD. Multivariate analysis of ecological data, Version 5.07. MjM Software Design, Oregon, USA. [ Links ]
Medina-Nava, M. 2003. Utilización del Índice de Integridad Biótica (IIB) para determinar áreas de conservación de peces en la cuenca Lerma-Chapala en Michoacán. Tesis de Maestría, Universidad Michoacana de San Nicolás de Hidalgo, Michoacán, México. [ Links ]
Mercado-Silva, N., J. Lyons, G. Salgado-Maldonado & M. Medina-Nava. 2002. Validation of a fish-based index of biotic integrity for streams and rivers of central Mexico. Rev. Fish Biol. Fish. 12: 179-191. [ Links ]
Mercado-Silva, N., M.R. Helmus & M.J. Vander Zanden. 2009. The effects of impoundment and non-native species on a river food web in Mexico’s Central Plateau. River Res. Applic. 25: 1090-1108. [ Links ]
Mielke, P. & H. Iyer. 1982. Permutation techniques for analyzing multi-response data from randomized block experiments. Commun. Stat.-Theor. M. 11: 1427-1437. [ Links ]
Miller, R.R., W.L. Minckley & S.M. Norris. 2009. Peces dulceacuícolas de México. CONABIO, SIMAC, ECOSUR, Consejo de Peces del Desierto, México D.F., México. [ Links ]
Navarrete, N., O. Mauleón & G. Contreras. 2008. Interacciones tróficas de los peces presentes en el embalse San Miguel Arco, Soyaniquilpan, Estado de México (otoño de 2006). Rev. Zool. 19: 7-14. [ Links ]
Orbe-Mendoza, A., J. Acevedo-Mendoza & J. Lyons. 2002. Lake Pátzcuaro fishery management plan. Rev. Fish Biol. Fish. 12: 207-217. [ Links ]
Pauly, D., V. Christensen, R. Froese & M. Palomares. 2000. Fishing down aquatic food webs. Am. Sci. 88: 46-51. [ Links ]
Pennak, W. 1978. Fresh Water Invertebrates of the United States. John Wiley & Sons, New Jersey, USA. [ Links ]
Pimm, S.L. 1982. Food webs. Chapman and Hall, New York, USA. [ Links ]
Pollux, B.J. & D.N. Reznick. 2011. Matrotrophy limits a female’s ability to adaptively adjust offspring size and fecundity in fluctuating environments. Func. Ecol. 25: 747-756. [ Links ]
Post, D., M. Pace & N. Hairston. 2000. Ecosystem size determines food-chain length in lakes. Nature 405: 1047-1049. [ Links ]
Rachlin, J.W., B.E. Warkentine & A. Papantoniou. 1989. The use of niche breadth and proportional similarity in feeding to stipulate resource utilization strategies in fish. J. Freshwater Ecol. 5: 103-112. [ Links ]
Rosenberger, E., S. Hampton, S. Fradkin & B. Kennedy. 2008. Effects ofshoreline development on the nearshore environment in large deep oligotrophic lakes.
Freshwater Biol. 53: 1673-1691. [ Links ]
Ruehl, C.B. & T.J. DeWitt. 2005. Trophic plasticity and fine-grained resource variation in populations of western mosquitofish, Gambusia affinis. Evol. Ecol. Res. 7: 801-19. [ Links ]
SAS Institute. 1995. JMP. Statistic Mode Visual. Version: 3.1.6.2. SAS Institute, Cary, NC, USA. [ Links ]
Schaefer, J., S. Heulett & T. Farrell. 1994. Interactions between two poeciliid fishes (Gambusia holbrooki and Heterandria formosa) and their prey in a Florida marsh. Copeia 2: 516-520. [ Links ]
Stevenson, R. & P. Smol. 2002. Use of algae in environmental assessments, p. 775-804. In J. D. Wehr & R. Sheath (eds.). Freshwater algae of North America: ecology and classification. Academic, New York, USA. [ Links ]
Toft, J., C. Simenstad, J. Crodell & L. Grimaldo. 2003. The effects of introduced water hyacinth on habitat structure, invertebrate assemblages, and fish diets. Estuaries 26: 746-758. [ Links ]
Trujillo-Jiménez, P. & H. Toledo-Beto. 2007. Alimentación de los peces dulceacuícolas tropicales Heterandria bimaculata y Poecilia sphenops (Cyprinidontiformes: Poeciliidae). Rev. Biol. Trop. 55: 603-615. [ Links ]
Vadeboncoeur, Y., K. McCann, J. Vander Zanden & J. Rasmussen. 2005. Effects of multi-chain omnivory on the strength of trophic control in lakes. Ecosystems 8: 682-693. [ Links ]
Vander Zanden, J. & J. Rassmussen. 1999. Primary consumer δ13C and δ15N and the trophic position of aquatic consumers. Ecology 80: 1395-1404. [ Links ]
Vander Zanden, J., S. Chandra, B.C. Allen, J.E. Reuter & C.R. Goldman. 2003. Historical food web structure and restoration of native aquatic communities in the Lake Tahoe (California-Nevada) basin. Ecosystems 6: 274-288. [ Links ]
Vanni, M.J., K.K. Arend, M.T. Bremigan, D.B. Bunnel, J.E. Garvey, M.J. González, W.H. Renwick, P.A. Soranno & R.A. Stein. 2005. Linking landscapes and food webs: effects of omnivorous fish and watersheds on reservoir ecosystems. Bioscience 2: 155-167. [ Links ]
Varela-Romero, A., G. Ruíz-Campos, L.M. Yépiz-Velázquez & J. Alaníz-García. 2002. Distribution, habitat, and conservation status of desert pupfish (Cyprinodon macularius) in the Lower Colorado River Basin, Mexico. Rev. Fish. Biol. Fisher. 12: 157-165. [ Links ]
Vega-Cendejas, M.E. 1990. Interacción trófica entre dos bagres Arius melanopus (Agassiz, 1829) y Arius felis (Linnaeus, 1776) en las costas de Celestún Yucatán, México. An. Inst. Cienc. Del Mar y Limnol. U. Nac. Auton. Mexico 15: 185-194. [ Links ]
Vital-Rodríguez, B. 2011. Hábitos alimentarios del género Chirostoma del lago de Pátzcuaro, Michoacán, México. Tesis de Maestría, Universidad Michoacana de San Nicolás de Hidalgo, Michoacán, México. [ Links ]
Wilcoxon, F. 1945. Individual comparisons by ranking methods. Biometrics Bull. 1: 80-83. [ Links ]
Yáñez-Arancibia, A., J. Curiel & V. Leyton. 1976: Prospección biológica y ecológica del bagre marino Galeichthys caerulescens (Günther) en el sistema lagunar costero de Guerrero, México (Pisces: Ariidae). An. Inst. Cienc. Mar y Limnol. U. Nac. Auton. Mex. 3: 125-180. [ Links ]
Zar, J. 1999. Biostatistical Analysis. Prentice Hall, New Jersey, USA. [ Links ]
Zubieta, T. 1985. Estudio sobre los hábitos alimenticios de seis especies de peces del Lago de Cuitzeo, Mich. Tesis de Licenciatura, Universidad Michoacana de San Nicolás de Hidalgo, Michoacán, México. [ Links ]
Armendáriz, M.A., N.E. Navarrete, E. Fernández, G. Vázquez & E.S. Urrieta. 2008. Relaciones tróficas de los peces del embalse San Miguel Arco, de Soyaniquilpan, Estado de México. Rev. Chapingo Ser. Cienc. 14: 33-38. [ Links ]
Barbour, M.T., J. Gerritsen, B. Zinder & J. Stribling. 1999. Rapid bioassessment protocols for use in streams and wadeable rivers: periphyton, benthic macroinvertebrates and fish. U.S. Environmental Protection Agency, Washington, D.C., USA. [ Links ]
Bearhop, S., C.E. Adams, S. Waldron, R.A. Fuller & H. Macleod. 2004. Determining trophic niche width: a novel approach using stable isotope analysis. J. Anim. Ecol. 73: 1007-1012. [ Links ]
Bernal-Brooks, F., T. Gómez & J. Alcocer. 2002. Lake Pátzcuaro (Mexico): a controversy about the ecosystem water regimen approached by field references, climatic variables, and GIS. Hydrobiologia 467: 187-197. [ Links ]
Berry, J., E. Lee, K. Walton, A. Wilson & F. Bernal-Brooks. 2011. Microcystin production by a persistent cyanobacterial bloom in Lago de Pátzcuaro (Michoacán, Mexico), and apparent bioaccumulation of the toxin in small commercial catches of fish. Environ. Toxicol. Chem. 30: 1621-1628. [ Links ]
Biondini, M., P. Mielke & K. Berry. 1988. Data-dependent permutation techniques for the analysis of ecological data. Vegetatio 75: 161-168. [ Links ]
Bouyoucos, G.J. 1936. Directions for making mechanical analyses of soils by the hydrometer method. Soil Sci. 42: 225-30. [ Links ]
Caillet, G., M. Love & A. Ebeling. 1996. Fishes a Field and Laboratory Manual on Their Structure, Identification and Natural History. Wadsworth, Belmont, CA, USA. [ Links ]
Canto-Maza, W. & M.E. Vega-Cendejas. 2008. Hábitos alimenticios del pez Lagodon rhomboides (Perciformes: Sparidae) en la laguna costera de Chelem, Yucatán, México. Rev. Biol. Trop. 56: 1837-1846. [ Links ]
Christensen, V. & D. Pauly. 1992. ECOPATH II a software for balancing steady-state ecosystem models and calculating network characteristics. Ecol. Model. 61: 169-185. [ Links ]
Cortés, E. 1997. A critical review of methods of studying fish feeding based on analysis of stomach contents: application to elasmobranch fishes. Can. J. Fish. Aquat. Sci. 54: 726-738. [ Links ]
Duarte, m. 1981. Contribución al conocimiento de los hábitos alimentarios de doce especies de peces en el lago de Cuitzeo, Michoacán. Tesis de Licenciatura, Instituto Politécnico Nacional, México, D.F., México. [ Links ]
Edmondson, T. 1959. Freshwater Biology. Wiley & Sons, New Jersey, USA. [ Links ]
Escalera-Gallardo, C. 1986. Alimentación de los peces en la presa Guaracha, Michoacán. Informe TécnicoNo. 22. CIIDIR-IPN-MICHOACÁN, Michoacán, Mexico. [ Links ]
Fagan, F. 1997. Omnivory as a stabilizing feature of natural communities. Am. Nat. 150: 554-567. [ Links ]
Galindo-Villegas, J. & E. Sosa-Lima. 2002. Gonopodial system review and a new fish record of Poeciliopsis infans Cyprinodontiformes: Poeciliidae) for Lake Pátzcuaro, Michoacán, Central Mexico. Rev. Biol. Trop. 50: 1151-1157. [ Links ]
Gkenas, C., A. Oikonomou, A. Economou, F. Kiosse & I. Leonardos. 2012. Life history pattern and feeding habits of the invasive mosquitofish, Gambusia holbrooki, in Lake Pamvotis (NW Greece). J. Biol. Res.- Thessalon. 17: 121-136. [ Links ]
Gómez-Márquez, J., B. Peña-Mendoza, I. Salgado-Ugarte, A. Sánchez-Herrera, L. Sastré-Baez. 2008. Reproduction of the fish Poeciliopsis gracilis (Cyprinodontiformes: Poeciliidae) in Coatetelco, a tropical shallow lake in Mexico. Rev. Biol. Trop. 56: 1801-1812. [ Links ]
Hynes, B. 1950. The food of fresh-water sticklebacks (Gasterosteus aculeatus and Pygosteus pungitius), with a review of methods used in studies of the food of fishes. J. Anim. Ecol. 19: 36-57. [ Links ]
Jardine, T.D., S. McGeachy, C. Paton, M. Savoie & R. Cunjak. 2003. Stable isotopes in aquatic systems: Sample preparation, analysis, and interpretation. Canadian Manuscript Report of Fisheries and Aquatic Sciences No 2656. Fisheries and Oceans Canada, Ontario, Canada. [ Links ]
Karr, J.R. 1981. Assessment of biotic integrity using fish communities. Fisheries 6: 21-27. [ Links ]
King, S. & K. Warburton. 2007. The environmental preferences of three species of Australian freshwater fish in relation to the effects of riparian degradation.Environ. Biol. Fish. 78: 307-316. [ Links ]
Koehn, J.D. 2004. Carp (Cyprinus carpio) as a powerful invader in Australia waterways. Freshwater Biol. 49: 882-894. [ Links ]
Krebs, J. 1989. Ecological Methodology. Harper & Row, New York, USA. [ Links ]
Lampert, W. & U. Sommer. 2007. Limnoecology. Oxford University, Oxford, England. [ Links ]
Lyons, J., S. Navarro-Pérez, A.P. Cochran, C.E. Santana & M. Guzmán-Arroyo. 1995. Index of biotic integrity based on fish assemblages for the conservation of streams and rivers in west-central México. Conserv. Biol. 3: 569-584. [ Links ]
Lyons, J., A. Gutierrez-Hernandez, E. Diaz-Pardo, E. Soto-Galera, M. Medina-Nava & R. Pineda-López. 2000. Development of a preliminary index of biotic integrity (IBI) based on fish assemblages to assess ecosystem condition in the lakes of central Mexico. Hydrobiologia 418: 57-72. [ Links ]
Magurran, A. 2004. Measuring biological diversity. Blackwell, London, England. [ Links ]
Mar-Silva, V. 2011. Variación espacio-temporal de la comunidad de peces en el lago de Pátzcuaro, Michoacán, México. Tesis de Licenciatura, Universidad Michoacana de San Nicolás de Hidalgo, Michoacán, Mexico. [ Links ]
McCune, B. & J. Grace. 2002. MRPP (multi-response permutation procedures) in analysis of ecological communities. MjM Software Design, Oregon, USA. [ Links ]
McCune, B. & J. Mefford. 2006. PC-ORD. Multivariate analysis of ecological data, Version 5.07. MjM Software Design, Oregon, USA. [ Links ]
Medina-Nava, M. 2003. Utilización del Índice de Integridad Biótica (IIB) para determinar áreas de conservación de peces en la cuenca Lerma-Chapala en Michoacán. Tesis de Maestría, Universidad Michoacana de San Nicolás de Hidalgo, Michoacán, México. [ Links ]
Mercado-Silva, N., J. Lyons, G. Salgado-Maldonado & M. Medina-Nava. 2002. Validation of a fish-based index of biotic integrity for streams and rivers of central Mexico. Rev. Fish Biol. Fish. 12: 179-191. [ Links ]
Mercado-Silva, N., M.R. Helmus & M.J. Vander Zanden. 2009. The effects of impoundment and non-native species on a river food web in Mexico’s Central Plateau. River Res. Applic. 25: 1090-1108. [ Links ]
Mielke, P. & H. Iyer. 1982. Permutation techniques for analyzing multi-response data from randomized block experiments. Commun. Stat.-Theor. M. 11: 1427-1437. [ Links ]
Miller, R.R., W.L. Minckley & S.M. Norris. 2009. Peces dulceacuícolas de México. CONABIO, SIMAC, ECOSUR, Consejo de Peces del Desierto, México D.F., México. [ Links ]
Navarrete, N., O. Mauleón & G. Contreras. 2008. Interacciones tróficas de los peces presentes en el embalse San Miguel Arco, Soyaniquilpan, Estado de México (otoño de 2006). Rev. Zool. 19: 7-14. [ Links ]
Orbe-Mendoza, A., J. Acevedo-Mendoza & J. Lyons. 2002. Lake Pátzcuaro fishery management plan. Rev. Fish Biol. Fish. 12: 207-217. [ Links ]
Pauly, D., V. Christensen, R. Froese & M. Palomares. 2000. Fishing down aquatic food webs. Am. Sci. 88: 46-51. [ Links ]
Pennak, W. 1978. Fresh Water Invertebrates of the United States. John Wiley & Sons, New Jersey, USA. [ Links ]
Pimm, S.L. 1982. Food webs. Chapman and Hall, New York, USA. [ Links ]
Pollux, B.J. & D.N. Reznick. 2011. Matrotrophy limits a female’s ability to adaptively adjust offspring size and fecundity in fluctuating environments. Func. Ecol. 25: 747-756. [ Links ]
Post, D., M. Pace & N. Hairston. 2000. Ecosystem size determines food-chain length in lakes. Nature 405: 1047-1049. [ Links ]
Rachlin, J.W., B.E. Warkentine & A. Papantoniou. 1989. The use of niche breadth and proportional similarity in feeding to stipulate resource utilization strategies in fish. J. Freshwater Ecol. 5: 103-112. [ Links ]
Rosenberger, E., S. Hampton, S. Fradkin & B. Kennedy. 2008. Effects ofshoreline development on the nearshore environment in large deep oligotrophic lakes.
Freshwater Biol. 53: 1673-1691. [ Links ]
Ruehl, C.B. & T.J. DeWitt. 2005. Trophic plasticity and fine-grained resource variation in populations of western mosquitofish, Gambusia affinis. Evol. Ecol. Res. 7: 801-19. [ Links ]
SAS Institute. 1995. JMP. Statistic Mode Visual. Version: 3.1.6.2. SAS Institute, Cary, NC, USA. [ Links ]
Schaefer, J., S. Heulett & T. Farrell. 1994. Interactions between two poeciliid fishes (Gambusia holbrooki and Heterandria formosa) and their prey in a Florida marsh. Copeia 2: 516-520. [ Links ]
Stevenson, R. & P. Smol. 2002. Use of algae in environmental assessments, p. 775-804. In J. D. Wehr & R. Sheath (eds.). Freshwater algae of North America: ecology and classification. Academic, New York, USA. [ Links ]
Toft, J., C. Simenstad, J. Crodell & L. Grimaldo. 2003. The effects of introduced water hyacinth on habitat structure, invertebrate assemblages, and fish diets. Estuaries 26: 746-758. [ Links ]
Trujillo-Jiménez, P. & H. Toledo-Beto. 2007. Alimentación de los peces dulceacuícolas tropicales Heterandria bimaculata y Poecilia sphenops (Cyprinidontiformes: Poeciliidae). Rev. Biol. Trop. 55: 603-615. [ Links ]
Vadeboncoeur, Y., K. McCann, J. Vander Zanden & J. Rasmussen. 2005. Effects of multi-chain omnivory on the strength of trophic control in lakes. Ecosystems 8: 682-693. [ Links ]
Vander Zanden, J. & J. Rassmussen. 1999. Primary consumer δ13C and δ15N and the trophic position of aquatic consumers. Ecology 80: 1395-1404. [ Links ]
Vander Zanden, J., S. Chandra, B.C. Allen, J.E. Reuter & C.R. Goldman. 2003. Historical food web structure and restoration of native aquatic communities in the Lake Tahoe (California-Nevada) basin. Ecosystems 6: 274-288. [ Links ]
Vanni, M.J., K.K. Arend, M.T. Bremigan, D.B. Bunnel, J.E. Garvey, M.J. González, W.H. Renwick, P.A. Soranno & R.A. Stein. 2005. Linking landscapes and food webs: effects of omnivorous fish and watersheds on reservoir ecosystems. Bioscience 2: 155-167. [ Links ]
Varela-Romero, A., G. Ruíz-Campos, L.M. Yépiz-Velázquez & J. Alaníz-García. 2002. Distribution, habitat, and conservation status of desert pupfish (Cyprinodon macularius) in the Lower Colorado River Basin, Mexico. Rev. Fish. Biol. Fisher. 12: 157-165. [ Links ]
Vega-Cendejas, M.E. 1990. Interacción trófica entre dos bagres Arius melanopus (Agassiz, 1829) y Arius felis (Linnaeus, 1776) en las costas de Celestún Yucatán, México. An. Inst. Cienc. Del Mar y Limnol. U. Nac. Auton. Mexico 15: 185-194. [ Links ]
Vital-Rodríguez, B. 2011. Hábitos alimentarios del género Chirostoma del lago de Pátzcuaro, Michoacán, México. Tesis de Maestría, Universidad Michoacana de San Nicolás de Hidalgo, Michoacán, México. [ Links ]
Wilcoxon, F. 1945. Individual comparisons by ranking methods. Biometrics Bull. 1: 80-83. [ Links ]
Yáñez-Arancibia, A., J. Curiel & V. Leyton. 1976: Prospección biológica y ecológica del bagre marino Galeichthys caerulescens (Günther) en el sistema lagunar costero de Guerrero, México (Pisces: Ariidae). An. Inst. Cienc. Mar y Limnol. U. Nac. Auton. Mex. 3: 125-180. [ Links ]
Zar, J. 1999. Biostatistical Analysis. Prentice Hall, New Jersey, USA. [ Links ]
Zubieta, T. 1985. Estudio sobre los hábitos alimenticios de seis especies de peces del Lago de Cuitzeo, Mich. Tesis de Licenciatura, Universidad Michoacana de San Nicolás de Hidalgo, Michoacán, México. [ Links ]
*Correspondencia: Juan P. Ramírez-Herrejón: Centro de Investigaciones Biológicas del Noroeste (CIBNOR), Instituto Politécnico Nacional 195, Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23096, México; ramirezherrejon@gmail.com
Lissette S. Castañeda-Sam: Laboratorio de Biología Acuática, Facultad de Biología, Universidad Michoacana de San Nicolás de Hidalgo (UMSNH), Morelia, Michoacán 58080, México; suhying_tataco@hotmail.com
Rodrigo Moncayo-Estrada: Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional, Instituto Politécnico Nacional, Unidad Michoacán (CIIDIR-Michoacán). Justo Sierra 28, Col. Centro, Jiquilpan, Michoacán 59510, México; rmoncayo@hotmail.com
Javier Caraveo-Patiño: Centro de Investigaciones Biológicas del Noroeste (CIBNOR), Instituto Politécnico Nacional 195, Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23096, México; jcaraveo04@cibnor.mx*
Eduardo F. Balart: Centro de Investigaciones Biológicas del Noroeste (CIBNOR), Instituto Politécnico Nacional 195, Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23096, México; ebalart04@cibnor.mx
1. Centro de Investigaciones Biológicas del Noroeste (CIBNOR), Instituto Politécnico Nacional 195, Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23096, México; ramirezherrejon@gmail.com, jcaraveo04@cibnor.mx*, ebalart04@cibnor.mx
2. Laboratorio de Biología Acuática, Facultad de Biología, Universidad Michoacana de San Nicolás de Hidalgo (UMSNH), Morelia, Michoacán 58080, México; suhying_tataco@hotmail.com
3. Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional, Instituto Politécnico Nacional, Unidad Michoacán (CIIDIR-Michoacán). Justo Sierra 28, Col. Centro, Jiquilpan, Michoacán 59510, México; rmoncayo@hotmail.com
Received 02-VII-2012. Corrected 08-XI-2012. Accepted 07-XII-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}