










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
Identification of endangered or threatened Costa Rican tree species by wood anatomy and fluorescence activity
Identificación de especies de árboles en peligro o amenazadas de Costa Rica basada en la anatomía de la madera y fluorescencia
Identificación de especies de árboles en peligro o amenazadas de Costa Rica basada en la anatomía de la madera y fluorescencia
*Dirección para correspondencia:
Abstract
A total of 45 native Costa Rican tree species are threatened or in danger of extinction, but the Convention on International Trade Endangered Species (CITES) includes only eight of these in its Appendices. However, the identification of other species based on their wood anatomy is limited. The present study objective was to describe and to compare wood anatomy and fluorescence activity in some endangered or threatened species of Costa Rica. A total of 45 (22 endangered and 23 threatened with extinction) wood samples of these species, from the xylaria of the Instituto Tecnológico de Costa Rica and the Forest Products Laboratory in Madison, Wisconsin, were examined. Surface fluorescence was positive in eight species, water extract fluorescence was positive in six species and ethanol extract fluorescence was positive in 24 species. Almost all species were diffuse porous except for occasional (Cedrela odorata, C. fissilis, Cordia gerascanthus) or regular (C. salvadorensis and C. tonduzii) semi-ring porosity. A dendritic vessel arrangement was found in Sideroxylon capari, and pores were solitary in Guaiacum sanctum and Vantanea barbourii. Vessel element length was shortest in Guaiacum sanctum and longest in Humiriastrum guianensis, Minquartia guianensis and Vantanea barbourii. Finally, anatomical information and fluorescence activity were utilized to construct an identification key of species, in which fluorescence is a feature used in identification.
Key words: fluorescence, Costa Rican woods, tropical woods, wood identification key.
Resumen
Un total de 45 especies de árboles de Costa Rica se catalogaron como amenazadas o en peligro de extinción, de las cuales, CITES (Convention on International Trade Endangered Species) incluye solamente ocho en sus Apéndices. Sin embargo, la identificación de las especies basadas en su anatomía es muy limitada. El presente estudio tiene el objetivo describir y comparar la anatomía y la fluorescencia de las especies amenazadas o en peligro de extinción de Costa Rica. Muestras de madera de las especies en peligro de extinción o amenazadas de la xiloteca del Instituto Tecnológico de Costa Rica y del Laboratorio de Productos Forestales de los Estados Unidos en Wisconsin se examinaron, se describió su anatomía, se evaluó su actividad fluorescente y se midió su densidad. La superficie de la madera fue fluorescente en ocho especies, el extracto en agua fue fluorescente en seis especies y el extracto en etanol fue positivo en 24 especies. Muchas de las especies presentaban porosidad difusa, excepto algunas Cedrela odorata, C. fissilis, Cordia gerascanthus C. salvadorensis y C. tonduzii que presentaban porosidad semi-anular. Vasos con distribución déndrica se encontró en Sideroxylon capari y poros solitarios en Guaiacum sanctum y Vantanea barbourii. Los vasos más cortos se encontraron en Guaiacum sanctum y los vasos más largos en Humiriastrum guianensis, Minquartia guianensis y Vantanea barbourii. Finalmente, la información de la anatomía y de su fluorescencia se utilizó para construir una clave de identificación, donde la actividad de fluorescencia juega un papel importante en la identificación.
Palabras clave: fluorescencia, madera de Costa Rica, maderas tropicales, clave de identificación.
For its size (51 000km2) Costa Rica is one of the most biodiverse countries in the world. Of its 10 000 plant taxa, 1 300 are endemic and more than 25% of these are considered rare (Burger 1980). Nonetheless, intensive deforestation began at the start of the twentieth century, and reached levels in excess of 50 000ha per year in the 1970s (Alvarez 1986). Natural forest decreased to only 24.4% by 1987 (Sánchez et al. 2001).
Tree timber species receive the highest degree of protection, and two actions are used to protect them: the promulgation of laws and government decrees to stop the cutting of 18 commercial species (Costa Rica 1997) protected by international conventions such as CITES, and actions limiting harvesting in certain areas of the country (Costa Rica 1996a, 1996b). A number of important timber-producing species have been identified by some experts as threatened species due to their rarity (Jimenez 1999). Identification of these timber-producing species using floral and tree characteristics has been widely described (Jimenez 1999). In contrast, identification of these species based on their wood anatomy is limited to only a few species: examples are the species included in CITES Appendices I and II. The CITES Identification Guide-Tropical Wood (CITES 2002) has a general identification key with illustrations of the transverse sections of each species. The book is intended for non-experts (Gasson et al. 2010) and its use is limited. More recently, the International Association of Wood Anatomists (IAWA) published several wood descriptions of tree timbers included in CITES (Gasson 2011, Gasson et al. 2011).
Recently, wood anatomical identification has been accompanied by other analyses that together with special methods, can achieve precise identification that are useful in identification of endangered or threatened species. Some of these techniques are relatively easy to implement, for example, wood density, surface fluorescence, or fluorescence of water or ethanol extracts. (Miller & Wiemann 2006, Guzman et al. 2008, Wiemann & Ruffinatto 2012). Studies of anatomical features accompanied with complex statistical procedures, such as the use of multivariate analysis, helps to identify CITES species (Gasson et al. 2010, MacLanchlan & Gasson 2010).
The objective of the present study was to describe and compare wood anatomy, fluorescence, and density of endangered or threatened species of Costa Rica, and to use these characteristics to build an identification key. The key will allow monitoring of the use and commercialization of these tree species (endangered or threatened) in Costa Rica, and perhaps will be also useful to other countries in the region.
Material and Methods
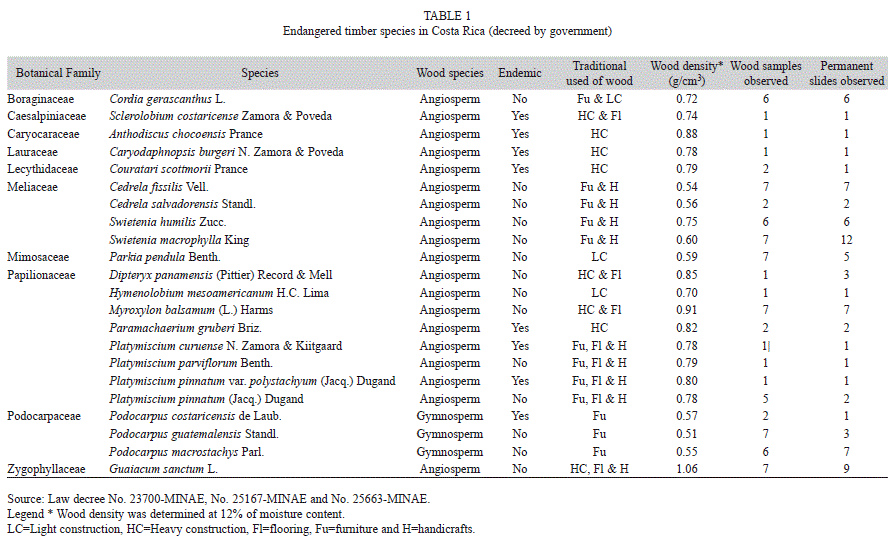
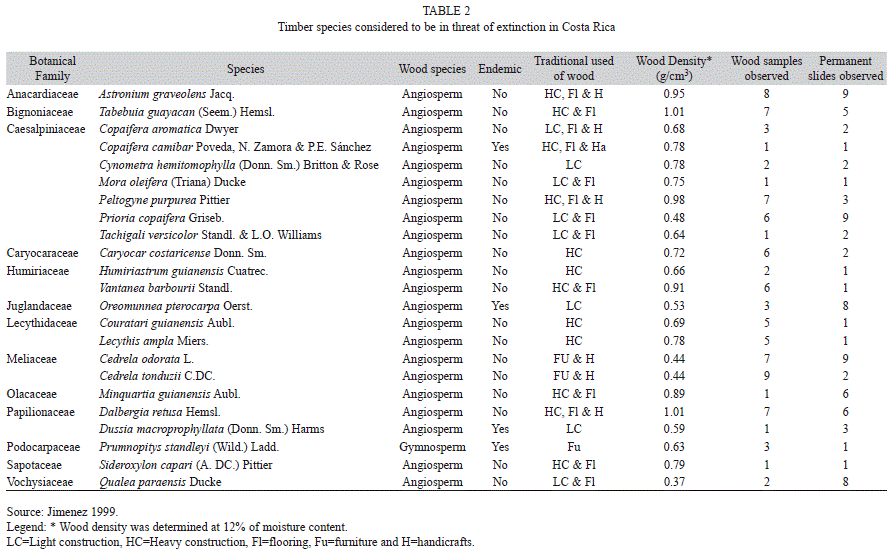
Wood samples and permanent slides: A total 45 timber species growing in Costa Rica were analyzed from March to December of 2010, 22 of which are considered as endangered species by the Costa Rican Government (Costa Rica 1996a and 1996b, Costa Rica 1997) (Table 1). Another 23 timber species studied are considered to be in threat of extinction in this country (Jimenez 1999) (Table 2). Wood samples of all the species were obtained from the Forest Products Laboratory (FPL), Wisconsin-USA collection (MADw and SJRw) and the Instituto Tecnológico de Costa Rica (ITCR) collection (TECw). Their sample numbers are detailed in tables 1 and 2. In some cases only one sample and slide was available, so it was necessary to prepare additional permanent slides of these species. To do this, a block (1cm3) was obtained from a wood sample and was softened in hot water. Tangential, radial and transverse sections were cut (12-15μm thick). These sections were stained with safranin and dehydrated with a series of alcohol (5 minutes each in 50, 70 and 95%); finally, sections were rinsed and mounted on microscope slides. Furthermore, a small piece was cut from each wood block to prepare macerated wood using Franklin’s method (Ruzin 1999).
Wood anatomical description: The IAWA list (IAWA 1989) was used as the basis for choosing identification characteristics, with some modifications to allow for increased accuracy and subsequent species level separation. The quantitative anatomical features that were measured were: length and diameter of the fibers, lumen diameter, cell wall thickness, vessel length, diameter and frequency of pores, solitary pore frequency, diameter of intervessel pits, and height and width of rays. Fiber dimensions and vessel lengths were measured on macerated wood. Permanent slides were used for measurement of the other anatomical characters. Qualitative anatomical features were also determined using the IAWA List as a guide (IAWA 1989).
Additional information: Other important information about endangered or threatened species used was: endemic category if the species grows only in Costa Rica, wood density, traditional uses of species, and fluorescence. Wood density (weight/volume) was measured in the air-dry condition. The four traditional uses considered were light or heavy construction, flooring, furniture, and handicrafts. Heartwood fluorescence was observed directly the surface, in water extract, and in ethanol extract, as described in the IAWA List (IAWA 1989). Surface fluorescence was determined in a darkroom from freshly prepared (planed or scraped) transverse and/or longitudinal surfaces exposed to a low intensity, long wave ultraviolet light (around 365nm). The specimens were recorded as either fluorescente (noting color and intensity) or not fluorescente. Only specimens which exhibited a definite yellow, green, orange or blue fluorescence were recorded as fluorescente. For water and ethanol extracts, color and intensity of fluorescence were scored. Froth tests were conducted and were scored as positive (high intensity) if one minute after shaking vigorously, froth was present and covered the entire surface of the solution, negative if all froth had disappeared, and variable if froth was still present around the edge of the test tube but did not extend over the entire surface of the liquid column.
Results
General aspects: 22% (ten species) were Papilionaceae, 18% (eight species) were Caesalpiniaceae, 13% (six species) were Meliaceae, 9% (four species) were Podocarpaceae, 7% (three species) were Lecythidaceae, 4% (two species) each were Caryocaraceae or Humiriaceae; other families represented the 27% (12 species) of endangered or threatened timber species endemic to Costa Rica. Most of these species are used in heavy or light construction or handicrafts; however, species in the Meliaceae are utilized for furniture manufacturing. High wood density was a characteristic of most of the species, and the main use of the lumber was construction (Table 1 and 2).
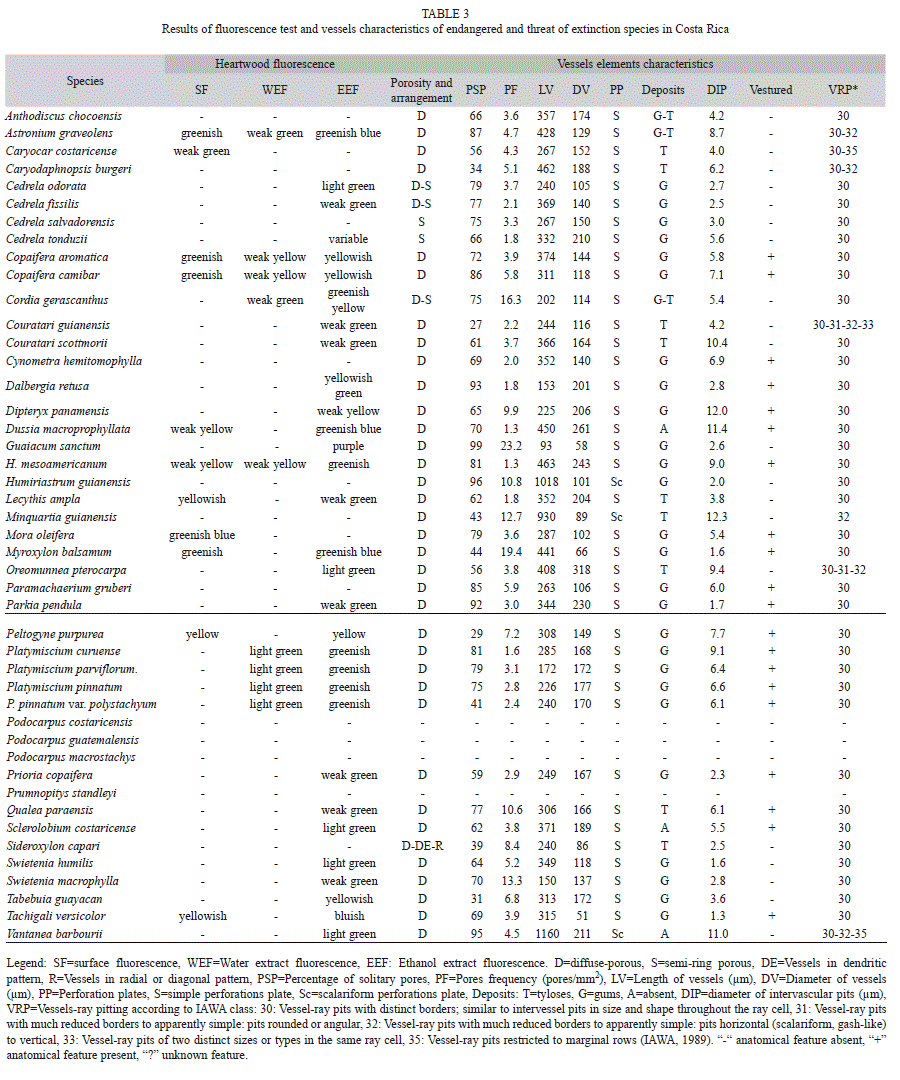
Fluorescence test: Surface fluorescence was positive in 11 species. This represented 24% of the total number of species. Fluorescence in Caryocar costaricense, Copaifera aromatica, Astronium graveolens, Mora oleifera and Myroxylon balsamum was green, greenish, or weak green, whereas in Dussia macroprophyllata, Lecythis ampla, Hymenolobium mesoamericanum, Peltogyne purpurea and Tachigali versicolor it was yellow, yellowish or weak yellow (Table 3).
Water extract fluoresced in only nine species (Table 3). The fluorescence was yellow in Copaifera species, H. mesoamericanum and P. purpurea and green in Cordia gerascanthus, A. graveolens, M. balsamum and Platymiscium species (Table 3). C. gerascanthus and Platymiscium species did not show surface fluorescence, but they did show water extract fluorescence (Table 3).
Ethanol extract fluorescence was found in 31 of the species. The fluorescence was greenish blue or yellow, weak or light green, purple, yellowish or bluish (Table 3). Several species (Cedrela odorata, Cedrela fissilis, Cedrela tonduzii, Couratari guianensis, Couratari scottmorii, Dalbergia retusa, Dipteryx panamensis, Guaiacum sanctum, Oreomunnea pterocarpa, Parkia pendula, Prioria copaifera, Qualea paraensis, Swietenia humilis, Swietenia macrophylla, Tabebuia guayacan and Vantanea barborii) had positive ethanol extract fluorescence, but negative surface or water extract fluorescence (Table 3).
General features of endangered or threatened species: Porosity: Diffuse porosity was common in endangered or threatened species, being found in 39 species. Three species were both diffuse and semi-ring porous (C. odorata, C. fissilis and C. gerascanthus) and two species were semi-ring porous (Cedrela salvadorensis and C. tonduzii). Sideroxylon capari was unique with a radial or diagonal pore pattern (Fig. 1a). Almost all of the species had solitary pores and pore multiples. However, the percentage of solitary pores was higher than 85% in G. sanctum and V. barbourii, so these species can be classified as pores exclusively solitary. Higher frequency of multiple pores (>15 pores/mm2) was measured in three species: C. guianensis, P. purpurea and T. guayacan. Pores frequency was the highest in Guaiacum sanctum, and M. balsamo. Lower pore frequencies (<2 pores/mm2) were observed in C. tonduzii, D. retusa, D. macrophylla, H. mesoamericanum, L. ampla and Platymiscium curuense.
Vessel lengths for these species varied from 93 to 1 160mm (Table 3). Vessel length was shortest in G. sanctum and longest in H. guianensis, M. guianensis and V. barbourii. Small diameter vessels (from 50-100mm) were found G. sanctum, M. guianensis, M. balsamo, S. capari and T. versicolor. Large diameter vessels were found only in O. pterocarpa. Vessel diameters from 100-300mm were found in the other species.
Simple perforation plates were found in almost all species (Table 3). However, H. guianensis (with 10-20 bars), M. guianensis (with <=10 bars) and V. barbourii (with <=20 bars) had scalariform perforations.
Tyloses, deposits or gum were found in the vessels of almost all of the angiosperm species (Table 3). Gum was the most common substance in vessel lumina. Nevertheless, neither substance was observed in three of the angiosperm species: D. macroprophyllata, Sclerolobium costaricense and V. barbourii.
All angiosperm species had alternate polygonal pits. Their diameter was minute (<4μm) in 14 species and large (>=10μm) in five species. Pits in the other species were from 4 to 10μm in diameter (Table 3).
Vestured pits were found in all species of Fabaceae (Caesalpixiaceae, Mimosaceae, Papilionaceae) and in V. barbourii (Table 3). Vessel-ray pits with distinct borders, similar to intervessel pits in size and shape throughout the ray cell, were common. They were found in almost all species. Other pits shapes were found in C. guianensis (Table 3). Vessel-ray pits were restricted to marginal rows in C. costaricense. Vessel-ray pits with much reduced borders to apparently simple, with pits rounded or angular and horizontal (scalariform, gashlike) to vertical (palisade) were found in A. graveolens and Caryodaphnopsis. burgeri (Fig. 1c), C. guianensis, and O. pterocarpa. Vesselray pits were restricted to marginal rows in V. barbourii. L. ampla had vessel-ray pits with much reduced borders to apparently simple: the pits were horizontal (scalariform, gash-like) to vertical (palisade).
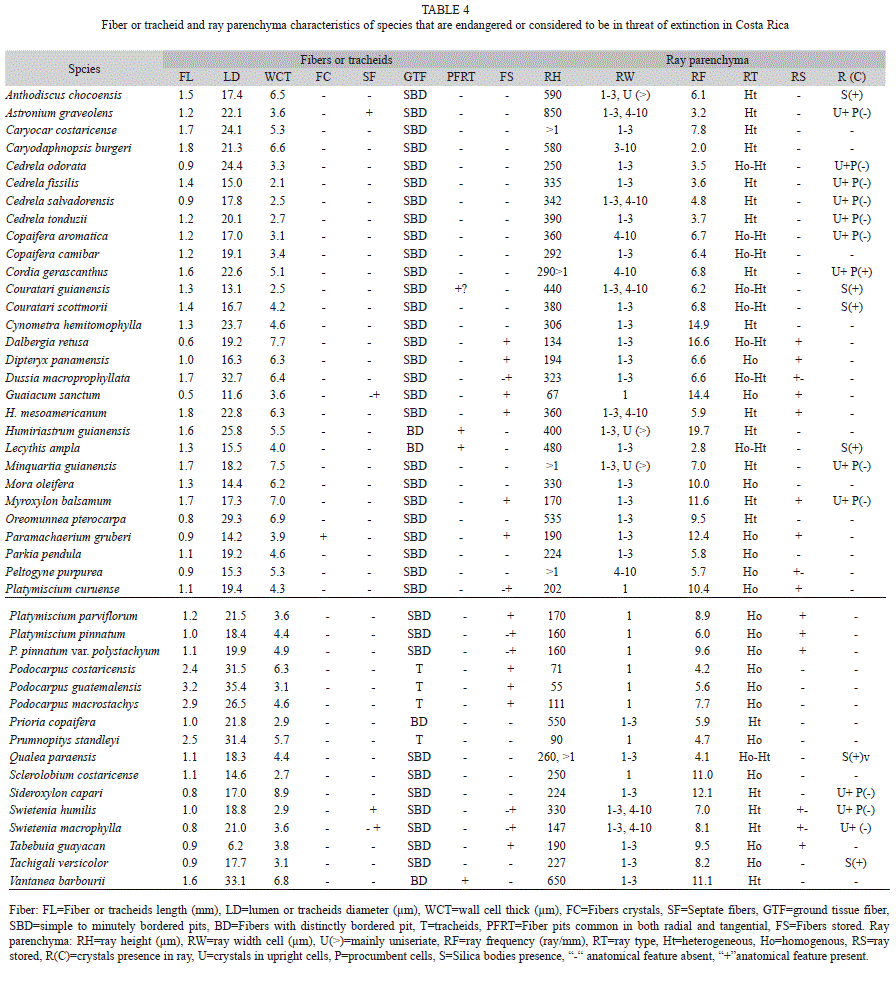
Fibers: as expected, gymnosperm species tracheids were longer than most of the angiosperm fibers, being almost 2mm length. However, the fiber length of V. barbourii was the longest of any species at 2.45mm. Fiber length varied from 1.50 to 1.95mm in C. costaricense, C. burgeri, C. gerascanthus, D. macroprophyllata, H. mesoamericanum, H. guianensis, M. guianensis and M. balsamum. The shortest fiber lengths were found in G. sanctum, although C. odorata, C. salvadorensis, D. retusa, O. pterocarpa and S. capari also had short fibers (Table 4). Fiber lumen diameter was widest in D. macroprophyllata, O. pterocarpa, H. guianensis, P. macrostachys and V. barbourii, and narrowest in T. guayacan. Narrow fibers (<15μm) were also found in C. fissilis, C. guianensis, G. sanctum, Paramachaerium gruberi and S. costaricense. The lumen diameter varied from 15 to 55μm in other species (Table 4). Fibers with very thick-walls (>7μm) were found in D. retusa, H. guianensis, M. guianensis, M. balsamum, S. capari and V. barbourii. Thin-walled fibers were found in species of Cedrela and in C. guianensis, P. copaifera, S. costaricense and S. humilis. Cell wall thickness of other endangered and threatened species varied from 3 to 7μm (Table 4). Septate fibers were observed only in A. graveolens, G. sanctum and Swietenia species (Table 4). In the angiosperms, fibers with simple to minutely bordered pits were most common in H. guianensis, L. ampla and V. barbourii. Distinctly bordered pits were common in both radial and tangential fiber walls. The tracheids of species of the Podocarpaceae had, as expected, bordered pits in radial walls. Many of the angiosperm species had storied fibers (Table 4). Crystals were only found in the fibers of P. gruberi.
Ray parenchyma: Ray height over 1mm was observed in five species (C. costaricense, C. gerascanthus, M. guianensis, P. purpurea and Q. paraensis), representing 11% of the species. Rays were exclusively uniseriate in eight species (Fig. 1b): four Podocarpaceae (Fig. 1d and 1e) and four angiosperms (G. sanctum and Platymiscium species). Rays 1-3 cell in width were the most common, represented by 23 species (51% of total species) (Table 4). Large rays (over 8 cells in width) were observed in eight species (18% of total species) (Table 4). The rays of C. aromatica, C. gerascanthus and P. purpurea were 4-10 seriate. Ray frequency varied from 2 to 20 rays per mm (Table 4). The lowest frequencies were found in C. burgeri and L. ampla and the highest ones (>14 rays/mm) were in Cynometra hemitomophylla, G. sanctum and H. guianensis. Rays composed mostly of procumbent cells were observed in 17 species (38% of total species) and heterogeneous rays were in 19 species (42% of total species). Nine species had both homogeneous and heterogeneous rays (Table 4). Storied rays were found in 14 species (Table 4). Two heights were found in D. macrophylla, P. purpurea and Swietenia species. Crystals were observed in ray cells of 27 species (60% of total species); in 11 species (24%) crystals were found in the marginal ray cells only (Table 4). Silica bodies were present in Anthodiscus chocoensis, Couratari species, L. ampla, Q. paraensis and T. versicolor. Sheath cells were only observed in C. gerascanthus.
Axial parenchyma: Apotraqueal parenchyma was present in 21 species (48% of total species) (Table 5). Parenchyma diffuse and diffuse-in-aggregates was seen in D. retusa, G. sanctum, M. guianensis and S. capari, and it was the only parenchyma type in C. costaricense. Almost all the species of angiosperms had paratracheal parenchyma (Table 5). Scanty paratracheal parenchyma was present in sixteen species, and vasicentric parenchyma was found in A. graveolens, C. odorata, C. fissilis, C. tonduzii, C. gerascanthus and T. versicolor. Three or more different paratracheal types were observed in 17 species (38% of total species). Aliform and confluent parenchyma was very common in several species. Banded parenchyma was observed in 30 species (68 of total species; 73% of angiosperm species). Parenchyma marginal or in wide bands (more than 3 cells wide) were the most common banded parenchyma (Table 5). Axial parenchyma was reticulate in Couratari species, C. hemitomophylla, L. ampla and O. pterocarpa. Axial parenchyma was storied in 13 species (Table 5). Fusiform cells were found in Platymiscium species, Paramachaerium grugeri, M. balsamo, P. copaifera, T. guayacan and T. versicolor. Crystals were present in chambered axial parenchyma in almost all gymnosperm species (Table 5). They were found enlarged in P. gruberi. A. chocoensis, A. graveolens, C. gerascanthus and Podocarpus species, T. guayacan did not have any crystals. Silica bodies were present in Couratari species.
Other anatomical features: some species had other distinctive anatomical features, which are detailed in table 5. They could be used to facilitate wood identification. For example, radial or axial canals are found in A. chocoensis, C. costaricense, C. aromatica and P. copaifera. Traumatic canals were observed in C. camibar, H. guianensis and Platymiscium pinnatum. Pores with two distinct diameters are found in D. retusa and Q. paraensis. C. gerascanthus was a unique species with sheath cell in the rays. Finally, helical thickenings were observed in some vessels elements of S. costaricense.
Specific species: Major differences among species groups are included in the next few paragraphs.
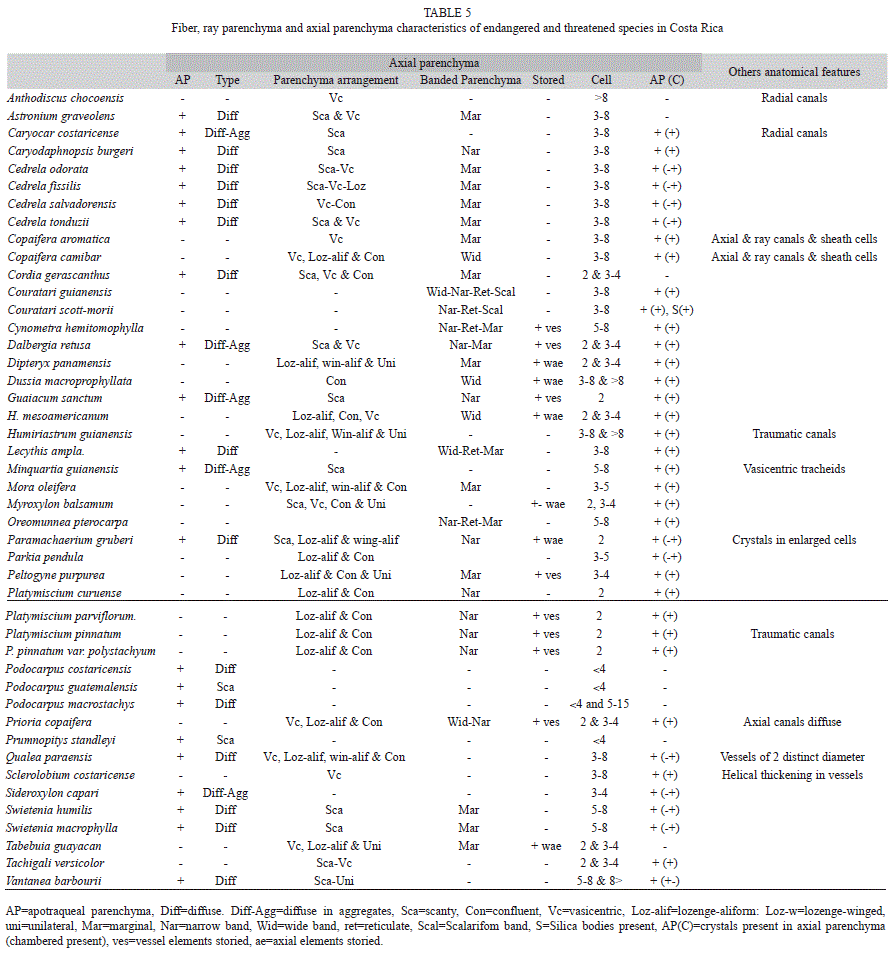
Meliaceae species: Two genera of Meliaceae were analyzed: four species of Cedrela (C. odorata, C. salvadorensis, C. fissilis and C. tonduzii), and two species of Swietenia (S. macrophylla and S. humilis). The wood anatomy of these species, especially C. odorata, C. fissilis and S. macrophylla, has been described by several authors (Panshin 1933, White & Gasson 2008). Four species of Meliaceae (C. salvadorensis, C. fissilis, S. macrophylla and S. humilis) are considered as endangered and their cutting has been prohibited in the natural forest in Costa Rica (Costa Rica 1996a). Felling of the other species, C. odorata and C. tonduzii, is permitted. Therefore, it is important to have a method to separate the species. The species of Swietenia are easily separated from species of Cedrela by anatomical features such as storied rays (Fig. 2g, h, i) and diffuse porosity in Swietenia but not in Cedrela (Fig. 2a-2i).
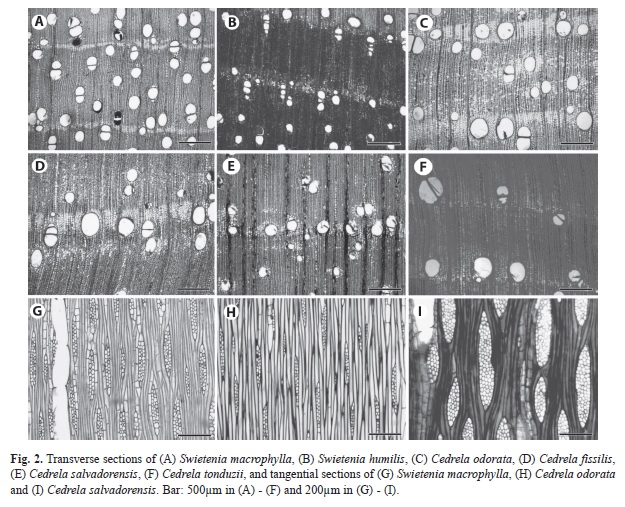
Platymiscium species: some differences were found among species of Platymiscium include presence of traumatic canals in Platymiscium parviflorum (Fig. 3a) but not in the other species (Fig. 3b). Platymiscium pinnatum var. polystachyum had irregularly storied rays (Fig. 4c) whereas in the other species storing was well-defined (Fig. 4a, 4b and 4d). The parenchyma was paratracheal in P. pinnatum var. polystachyum but not in other Platymiscium species. It was scanty, unilateral paratracheal, winged-aliform in P. pinnatum var polystachyum (Fig. 3b), but lozenge-aliform in the other species. We also found marginal parenchyma in P. pinnatum var polystachyum (Fig. 3b).
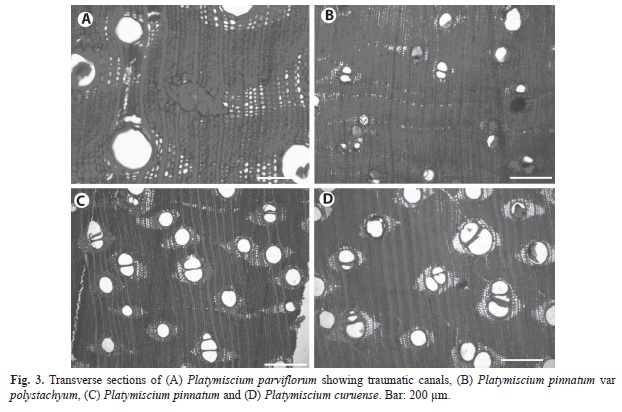
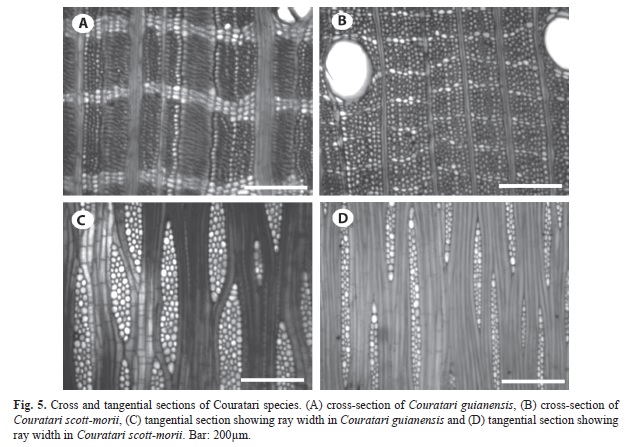
Couratari species: The two Couratari species can be distinguished by several differences. C. scottmorii has narrow reticulate parenchyma bands that are two cells wide (Fig. 5b), whereas reticulate bands in C. guianensis are up to four cells wide (Fig. 5a). Rays are 1-3 seriate in C. scottmorii (Fig. 5d), but up to 5-seriate in C. guianensis (Fig. 5c). The frequency of silica bodies is different among species, with, the highest frequency observed in C. guianensis. The vessels-ray pits in C. scottmorii have much reduced borders and the pits are rounded or angular, unlike those of C. guianensis which are horizontal (scalariform, gash-like) to vertical (palisade).
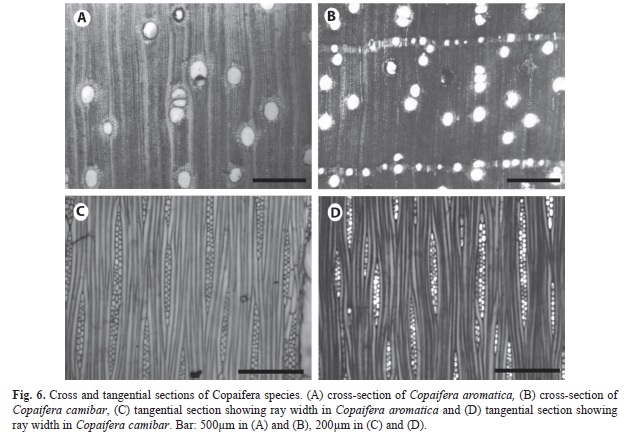
Copaifera species: The two Copaifera species differed mainly by paratracheal parenchyma and ray dimensions. Paratracheal parenchyma is more abundant in C. aromatica than in C. camibar. It is vasicentric 2-3 cells in width in Copaifetra aromatica (Fig. 6a), but scanty paratracheal or vasicentric 1-2 cells in C. camibar (Fig. 6b). The rays are 1- 3 cells wide and high in C. camibar (Fig. 6d), but were commonly 4-10 seriate and low in C. aromatic (Fig. 6c). There is no difference in the resin canals of the species.
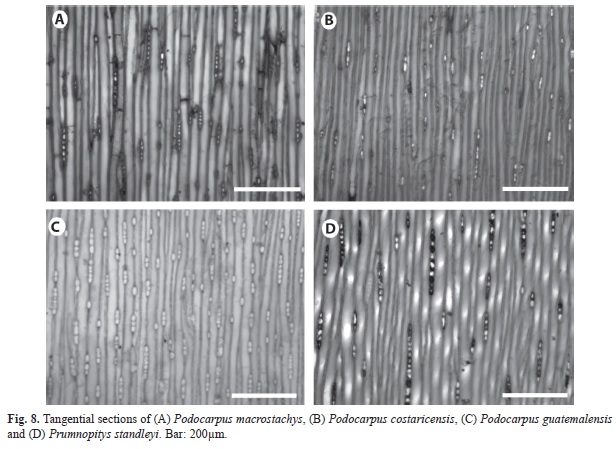
Podocarpaceae species: We looked at two genera: three species of Podocarpus (P. costaricensis, P. guatemalensis and P. macrostachys), and one species of Prumnopitys (P. standleyi). Axial parenchyma is present in P. macrostachys and P. costaricensis (Fig. 7a-7b) and is scanty in P. guatemalensis and Prumnopitys standleyi (Fig. 7c-7d). The highest proportion of axial parenchyma was observed in P. macrostachys (Fig. 7a), it was rare to moderately abundant in P. costaricensis (Fig. 7b) and scanty in P. guatemalensis (Fig. 7d). Another important difference between Podocarpus and Prumnopitys is ray height. Rays were highest in P. macrostachys (5 to 10 cells) (Fig. 8a), but they were only 2-4 cells high in P. costaricensis (Fig. 8b). Ray frequency is highest in P. guatemalensis (Fig. 7c). The rays of Prumnopitys standleyi are similar in shape and frequency to the rays of P. macrostachys. No differences were seen among species in crossfield pit apertures.
Discussion
Forty-five Costa Rican timber species are considered endangered or threatened. The Costa Rican government has decreed that 51% of these species are endangered and 49% of them are considered to be threatened. The CITES Appendices includes only eight of these species (S. humilis, S. macrophylla, D. panamensis, G. sanctum, C. costaricense, O. pterocarpa, C. odorata and D. retusa) (CITES 2002). All gymnosperms growing in the Costa Rican tropics are cataloged as endangered or threatened. However, most of the gymnosperm species in tropical areas around the world are in same situation; they are in decline or are restricted to isolated areas (Farjon et al. 1993).
The development of identification keys, like the one presented in this study, requires knowledge of wood characteristics and structure, as described in IAWA standards (IAWA 1989). Fluorescence provides a quick test for wood identification and it had been utilized by several authors for species separation. For example, Miller & Wiemann (2006) found differences in water and ethanol fluorescence between Dalbergia nigra and D. spruceana. Guzmán et al. (2008) found fluorescence species in the Anacardiaceae, Leguminose and Rubiaceae from Brazil and South Africa. Fluorescence is one of the important distinguishing characteristics of endangered or threatened timber species, although surface and water extract fluorescence can separate only six and eight timber species, respectively. Although, ethanol extract fluorescence was present in many timber species, the color of fluorescence was sometimes the same. However, three species were atypical in ethanol extract fluorescence color, the purple color found in G. sanctum, yellow in P. purpurea and bluish in T. versicolor, make them easy to identify. Guzmán et al. (2008) established that for Mexican timber identification it is necessary to use mixture of fluorescence tests and other characteristics, such as color or anatomical features. Likewise, identification of the endangered or threatened timber species from Costa Rica also requires anatomical studies. Species of Cedrela are easily separated from each other based on pore arrangement and axial parenchyma types. According to White & Gasson (2008), C. odorata is more ring-porous with more aliform parenchyma than C. fissilis. Cedrela tonduzii is similar to C. odorata and C. fissilis, although C. tonduzii is less ring porous. Aliform parenchyma is well defined in C. tonduzii, but not in C. odorata, C. salvadorensis or C. fissilis. C. odorata is considered to be easy to identify by its reddish color and distinct odor. Another important difference among C. tonduzii and other Cedrela species is that C. tonduzii has lower wood density than the other four species. C. salvadorensis has distinctive anatomical features that facilitate its identification; its rays are larger, commonly 4-10 seriate, than the rays of other Cedrela species, which are 1-3 seriate in width. Furthermore, its rays are heterogeneous and homogenous, but the rays of the other Cedrela species are not. However, Bonilla et al. (2004) reported that C. salvadorensis has 1-3 seriate rays, similar to those of other Cedrela. Therefore, we attribute the wider rays that we found in C. salvadorensis in Costa Rica to regional differences.
In species of Platymiscium, traumatic canals were seen in P. parviflorum, but not in other species. Traumatic canals have not been previously reported in Platymiscium (Espinoza & León 2002). Storied rays have been reported for most Platymiscium, for example P. lasiucarpium, P. duckei, P. pinnatum and P. yucatanum (Pérez 1993, Espinoza & León 2002). However, Detienne & Jacquet (1983) reported that rays are irregularly storied in P. ulei, similar to P. pinnatum var polystachyum.
The differences found in Couratari species agreed with previous research, which has also reported differences. For example Leon (2008) separated C. guianensis from C. multiflora by ray width, and Richter (1982) maintained that parenchyma distribution as well as type and configuration of inorganic contents can be employed for separating species of Lecythidaceae.
Canessa (1989) agreed with our results in Copaifera species; he found that the axial parenchyma in C. camibar was different than that of C. officinalis and C. pubiflora. Parenchyma is reported to be vasicentric, aliform to confluent, and aliform of the lozenge type in these species (Melandri & Espinoza de Pernía 2009). The ray dimensions and axial parenchyma of some Copaifera species are characterized by high anatomy variation (Regina et al. 2002), and our results confirm that.
On the other hand, Patel (1967) found similar results in Podocarpaceae species when he evaluated axial parenchyma. They mentioned that numerous species of the genus Podocarpus are characterized by diffuse axial parenchyma and a considerable variation of trached crossfield pit apertures (size, form, number per cross-field). Scanty axial parenchyma in P. guatemalensis can be used for separating this species from other Podocarpaceae in Costa Rica. This is in agreements with P. spicatus growing in New Zealand, in which the lack of axial parenchyma is the main feature to separating it from other Podocarpus species. Abundant axial parenchyma also separates P. macrostachys from P. costaricensis and P. guatemalensis. Patel (1967) agreed with this result: he found that P. dacrydioides can be separated from P. totara, P. hallii and P. acutifolius by the abundance of axial parenchyma. Bauch et al. (2006) mentioned that P. costarricensis has more axial parenchyma than the commercially important species P. salignus growing in Chile and Argentina.
Many of the Costa Rican species included in this study, are also present in other tropical regions, and our results, both as wood descriptions and the identification key, and are applicable to the wider region. Species conservation is a goal for many countries, especially in countries where deforestation of natural forests has increased in the last few years. There is a strong interest to protect timber species which have been over-exploited for many years. This identification key and wood descriptions will assist in the protection of species categorized as endangered or threatened, and will promote reliable conservation plans.
Acknowledgment
The authors wish to thank The Council for International Exchange of Scholars of the Department of Scholar and Professional Programs of USA, Premios Ford de Conservación of Ford Motor Company, and Vicerrectoría de Investigación y Extensión del Instituto Tecnológico de Costa Rica (ITCR) for financial support of this research.
References
Álvarez. D. 1986. Deforestación, causas y soluciones, p. 17-20. In COSTA RICA Ministerio de Gobernación y Policía. La verdadera emergencia nacional. San José, Costa Rica. [ Links ]
Bauch, J., G. Koch, J. Puls, T. Schwarz & S. Voiß. 2006. Wood characteristics of Podocarpus oleifolius var. Macrostachyus (Parl.) Buchholz and Gray native to Costa Rica: their significance for wood utilization. Wood Sci. Technol. 40: 26-38. [ Links ]
Bonilla, L., J. Barajas & P.T. Lezama. 2004. Anatomía de maderas de México. Árboles y Arbustos del Matorral Xerófilo de Tehuacán. Publicaciones Especiales del Instituto de Biología 19. Universidad Nacional Autónoma de México. Ciudad de México, México. [ Links ]
Burger, W.C. 1980. Why are there so many kinds of flowering plants in Costa Rica? Brenesia 17: 371-388. [ Links ]
Canessa, E. 1989. Descripción anatómica de la madera de camíbar (Copaifera camibar Poveda, Zamora & Sánchez). Brenesia 31: 113-115. [ Links ]
CITES, 2002. CITES Identification Guide – Tropical Woods. Guide to identification of tropical woods controlled under conventions of International Trade in Endangered species of wild fauna and flora. Authority of the Ministry of Environment. Ministry of Supply and Services, Canada. [ Links ]
Costa Rica. 1996a. Se declara una restricción para el aprovechamiento maderable de árboles de Almendro Dipteryx panamensis) (25167). MINAE, San José, Costa Rica. [ Links ]
Costa Rica. 1996b. Se mantiene la restricción a la corta o aprovechamiento del árbol conocido como Almendro nombre científico Dipteryx panamensis) (25663). MINAE, San José, Costa Rica. [ Links ]
Costa Rica. 1997. Declara en veda total aprovechamiento de árboles en peligro extinción indicados en el presente decreto (25700). MINAE, San José, Costa Rica. [ Links ]
Detienne, P. & P. Jacque. 1983. Atlas d’identification des bois de l’amazonie et des regions voiseines. Centre Technique Forestier Tropical, Nogent s/Marne, France. [ Links ]
Espinoza, N. & W. León. 2002. Estudio anatómico del leño de 56 especies de la subfamilia Papilionoideae (Leguminosae) en Venezuela. Rev. For. Venezolana 46: 59-71. [ Links ]
Farjon, A., C. Page & N. Schellevis. 1993. A preliminary word list of threatened conifer taxa. Biodivers. Conserv. 2: 304-326. [ Links ]
Gasson, P., R. Miller, D.J. Stekel, F. Whinder & K. Zieminska. 2010. Wood identification of Dalbergia nigra (CITES Appendix I) using quantitative wood anatomy, principal components analysis and naïve Bayes classification. Ann. Bot. 105: 45-56. [ Links ]
Gasson, P. 2011. How precise can wood identification be? Wood anatomy role in support of the legal timber trade, especially CITES. IAWA J. 32: 137-154. [ Links ]
Gasson, P., P. Baas & E. Wheeler. 2011. Wood anatomy of CITES-listed tree species. IAWA J. 32: 155-198. [ Links ]
Guzmán, J.A.S., H.G. Richter, R.R. Anda & F.J. Talavera. 2008 Wood fluorescence of commercial timbers marketed in Mexico. IAWA J. 29: 311-322. [ Links ]
IAWA, 1989. List of microscopic features for hardwood identification. IAWA Bull. 10: 226-332. [ Links ]
Jiménez, Q. 1999. Árboles maderables en peligro de extinción en Costa Rica. Instituto Nacional de Biodiversidad, Heredia, Costa Rica. [ Links ]
León, W. 2008. Estudio anatómico de la madera en 17 especies de la familia Lecythidaceae de Venezuela. Rev. For. Venezolana 52: 213-225. [ Links ]
MacLachlan, I.R. & P. Gasson. 2010. PCA of CITES listed Pterocarpus santalinus (Leguminosae) wood. IAWA J. 31: 121-138. [ Links ]
Melandri, J.L. & N. Espinoza de Pernía. 2009. Wood anatomy of tribe Detarieae and comparison with tribe Caesalpinieae (Leguminosae, Caesalpinioideae) in Venezuela. Rev. Biol. Trop. 57: 303-319. [ Links ]
Miller, R. & M. Wiemann. 2006. Separation of Dalbergia nigra from Dalbergia spruceana. Department of Agriculture, Research Paper FPL-RP-632. Forest Service, Forest Products Laboratory, Madison, Wisconsin, USA. [ Links ]
Panshin, A.J. 1933. Comparative Anatomy of the Woods of the Meliaceae, Sub-Family Swietenioideae. Am. J. Bot. 20: 638-668. [ Links ]
Patel, R.N. 1967. Wood anatomy of Podocarpaceae indigenous to New Zealand. NZ. J. Bot. 5: 307-321. [ Links ]
Pérez, C.P. 1993, Anatomía de la madera de ocho especie con importancia en las artesanías del estado Michoacán. Acta Bot. Mexicana 23: 103-136. [ Links ]
Regina, C., V. Angyalossy-Alfonso & L. Benetati. 2002. Anatomia comparada do lenho de Copaifera langsdorffii Desf. (Leguminosae-Caesalpinoideae) de floresta e cerradão. Rev. Bras. Bot. 24: 311-320. [ Links ]
Richter, 1982. The wood structure of Couratari Aubl and Couropita Aubl. (Lecythidaceae). IAWA J. 3: 45-54. [ Links ]
Ruzin, S.E. 1999. Plant microtechnique and Microscopy. Oxford University, Oxford, England. [ Links ]
Sánchez, G.A., R.C. Harris & D. Skole. 2001. Deforestation in Costa Rica: a quantitative analysis using remote sensing imagery. Biotropica 33: 378-384. [ Links ]
White, L. & P. Gasson. 2008. Mahogany. Kew: RBG Kew. 1-120 p. [ Links ]
Wiemann, M.C. & F. Ruffinatto. 2012. Separation of Dalbergia stevensonii from Dalbergia tucurensis. Research Paper FPL-RP-665. U.S. Department of Agriculture, Forest Service, Forest Products Laboratory, Madison, Wisconsin, USA. [ Links ]
Bauch, J., G. Koch, J. Puls, T. Schwarz & S. Voiß. 2006. Wood characteristics of Podocarpus oleifolius var. Macrostachyus (Parl.) Buchholz and Gray native to Costa Rica: their significance for wood utilization. Wood Sci. Technol. 40: 26-38. [ Links ]
Bonilla, L., J. Barajas & P.T. Lezama. 2004. Anatomía de maderas de México. Árboles y Arbustos del Matorral Xerófilo de Tehuacán. Publicaciones Especiales del Instituto de Biología 19. Universidad Nacional Autónoma de México. Ciudad de México, México. [ Links ]
Burger, W.C. 1980. Why are there so many kinds of flowering plants in Costa Rica? Brenesia 17: 371-388. [ Links ]
Canessa, E. 1989. Descripción anatómica de la madera de camíbar (Copaifera camibar Poveda, Zamora & Sánchez). Brenesia 31: 113-115. [ Links ]
CITES, 2002. CITES Identification Guide – Tropical Woods. Guide to identification of tropical woods controlled under conventions of International Trade in Endangered species of wild fauna and flora. Authority of the Ministry of Environment. Ministry of Supply and Services, Canada. [ Links ]
Costa Rica. 1996a. Se declara una restricción para el aprovechamiento maderable de árboles de Almendro Dipteryx panamensis) (25167). MINAE, San José, Costa Rica. [ Links ]
Costa Rica. 1996b. Se mantiene la restricción a la corta o aprovechamiento del árbol conocido como Almendro nombre científico Dipteryx panamensis) (25663). MINAE, San José, Costa Rica. [ Links ]
Costa Rica. 1997. Declara en veda total aprovechamiento de árboles en peligro extinción indicados en el presente decreto (25700). MINAE, San José, Costa Rica. [ Links ]
Detienne, P. & P. Jacque. 1983. Atlas d’identification des bois de l’amazonie et des regions voiseines. Centre Technique Forestier Tropical, Nogent s/Marne, France. [ Links ]
Espinoza, N. & W. León. 2002. Estudio anatómico del leño de 56 especies de la subfamilia Papilionoideae (Leguminosae) en Venezuela. Rev. For. Venezolana 46: 59-71. [ Links ]
Farjon, A., C. Page & N. Schellevis. 1993. A preliminary word list of threatened conifer taxa. Biodivers. Conserv. 2: 304-326. [ Links ]
Gasson, P., R. Miller, D.J. Stekel, F. Whinder & K. Zieminska. 2010. Wood identification of Dalbergia nigra (CITES Appendix I) using quantitative wood anatomy, principal components analysis and naïve Bayes classification. Ann. Bot. 105: 45-56. [ Links ]
Gasson, P. 2011. How precise can wood identification be? Wood anatomy role in support of the legal timber trade, especially CITES. IAWA J. 32: 137-154. [ Links ]
Gasson, P., P. Baas & E. Wheeler. 2011. Wood anatomy of CITES-listed tree species. IAWA J. 32: 155-198. [ Links ]
Guzmán, J.A.S., H.G. Richter, R.R. Anda & F.J. Talavera. 2008 Wood fluorescence of commercial timbers marketed in Mexico. IAWA J. 29: 311-322. [ Links ]
IAWA, 1989. List of microscopic features for hardwood identification. IAWA Bull. 10: 226-332. [ Links ]
Jiménez, Q. 1999. Árboles maderables en peligro de extinción en Costa Rica. Instituto Nacional de Biodiversidad, Heredia, Costa Rica. [ Links ]
León, W. 2008. Estudio anatómico de la madera en 17 especies de la familia Lecythidaceae de Venezuela. Rev. For. Venezolana 52: 213-225. [ Links ]
MacLachlan, I.R. & P. Gasson. 2010. PCA of CITES listed Pterocarpus santalinus (Leguminosae) wood. IAWA J. 31: 121-138. [ Links ]
Melandri, J.L. & N. Espinoza de Pernía. 2009. Wood anatomy of tribe Detarieae and comparison with tribe Caesalpinieae (Leguminosae, Caesalpinioideae) in Venezuela. Rev. Biol. Trop. 57: 303-319. [ Links ]
Miller, R. & M. Wiemann. 2006. Separation of Dalbergia nigra from Dalbergia spruceana. Department of Agriculture, Research Paper FPL-RP-632. Forest Service, Forest Products Laboratory, Madison, Wisconsin, USA. [ Links ]
Panshin, A.J. 1933. Comparative Anatomy of the Woods of the Meliaceae, Sub-Family Swietenioideae. Am. J. Bot. 20: 638-668. [ Links ]
Patel, R.N. 1967. Wood anatomy of Podocarpaceae indigenous to New Zealand. NZ. J. Bot. 5: 307-321. [ Links ]
Pérez, C.P. 1993, Anatomía de la madera de ocho especie con importancia en las artesanías del estado Michoacán. Acta Bot. Mexicana 23: 103-136. [ Links ]
Regina, C., V. Angyalossy-Alfonso & L. Benetati. 2002. Anatomia comparada do lenho de Copaifera langsdorffii Desf. (Leguminosae-Caesalpinoideae) de floresta e cerradão. Rev. Bras. Bot. 24: 311-320. [ Links ]
Richter, 1982. The wood structure of Couratari Aubl and Couropita Aubl. (Lecythidaceae). IAWA J. 3: 45-54. [ Links ]
Ruzin, S.E. 1999. Plant microtechnique and Microscopy. Oxford University, Oxford, England. [ Links ]
Sánchez, G.A., R.C. Harris & D. Skole. 2001. Deforestation in Costa Rica: a quantitative analysis using remote sensing imagery. Biotropica 33: 378-384. [ Links ]
White, L. & P. Gasson. 2008. Mahogany. Kew: RBG Kew. 1-120 p. [ Links ]
Wiemann, M.C. & F. Ruffinatto. 2012. Separation of Dalbergia stevensonii from Dalbergia tucurensis. Research Paper FPL-RP-665. U.S. Department of Agriculture, Forest Service, Forest Products Laboratory, Madison, Wisconsin, USA. [ Links ]
*Correspondencia a:
Róger Moya. Instituto Tecnológico de Costa Rica, Escuela de Ingeniería Forestal, Apartado 159-7050, Cartago, Costa Rica; rmoya@itcr.ac.cr. Corresponding author
Michael C. Wiemann. Forest Products Laboratory, USDA Forest Service, One Gifford Pinchot Drive Madison, Wisconsin 53726-2398, USA; mwiemann@fs.fed.us
Carlos Olivares. Instituto Tecnológico de Costa Rica, Escuela de Ingeniería Forestal, Apartado 159-7050, Cartago, Costa Rica; colivares@itcr.ac.cr
1. Instituto Tecnológico de Costa Rica, Escuela de Ingeniería Forestal, Apartado 159-7050, Cartago, Costa Rica; rmoya@itcr.ac.cr. Corresponding author
2. Forest Products Laboratory, USDA Forest Service, One Gifford Pinchot Drive Madison, Wisconsin 53726-2398, USA; mwiemann@fs.fed.us
3. Instituto Tecnológico de Costa Rica, Escuela de Ingeniería Forestal, Apartado 159-7050, Cartago, Costa Rica; colivares@itcr.ac.cr
Received 10-I-2012. Corrected 20-X-2012. Accepted 12-XI-2012


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}