










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
Establishment of cell suspension cultures of two Costa Rican Jatropha species (Euphorbiaceae)
Establecimiento de suspenciones celulares de dos especies Jatropha species (Euphorbiaceae) de Costa Rica
Establecimiento de suspenciones celulares de dos especies Jatropha species (Euphorbiaceae) de Costa Rica
*Dirección para correspondencia:
Abstract
J. curcas has been studied in different countries and some interesting agronomic, pharmacological and industrial properties have been reported. More recently, it has been considered an important alternative source for biofuel production. The objective of this study was to establish a long-term method for the maintenance of calli and cell suspension cultures of the local species J. curcas and J. gossypifolia, in order to allow future studies for novel compounds with pharmaceutical or industrial applications. For this, friable calli were successfully induced from hypocotyl segments of J. curcas and J. gossypifolia that were cultured in semisolid MS media supplemented with 1.5mg/L, and 0.5mg/L of 2,4-D, respectively. Cell suspension cultures of J. curcas were established using 1g of 35 and 60-day calli, in 50mL of liquid MS media supplied with 1.5mg/L of 2,4-D; sucrose and maltose were additionally evaluated as carbon sources. After 35 days, cell suspension cultures initiated with 35-day calli, showed greater cell growth with a maximum biomass of 194.9g/L fresh weight, 6.59g/L dry weight and 17.3% packed volume. The exponential phase ended at day 35 for cultures initiated with 35-day calli, and at day 21 for cultures initiated with 60-day calli. Higher biomass production was obtained with sucrose. Cell cultures were established with 35-day calli in MS media with the same 2,4-D concentration used for calli induction and 30g/L sucrose. This medium was considered optimum for the maintenance and growth of cell suspensions for both species, with sub-cultures every 20 days. The biotechnological potential for the production of bioactive compounds in these species for pharmacological, agricultural and industrial applications is being evaluated.
Key words: Jatropha curcas, Jatropha gossypifolia, calli, cell suspension culture, growth kinetics.
Resumen
J. curcas es un importante recurso alternativo de biocombustible. Por otro lado, propiedades de interés agronómico, farmacológico e industrial han sido reportadas para esta especie. El objetivo de este estudio fue el establecimiento y mantenimiento a largo plazo de callos y cultivos celulares en suspensión de J. curcas y J. gossypifolia, con el objetivo de permitir futuros estudios para nuevos compuestos con aplicaciones farmaceúticas e industriales. Los callos friables fueron exitosamente inducidos a partir de segmentos de hipocótilos J. curcas and J. gossypifolia cultivados en medio MS semisólido suplementado con 1.5mg/L y 0.5mg/L of 2,4-D, respectivamente. Los cultivos celulares en suspensión de J. curcas fueron establecidos utilizando 1g de callos de 35 y 60 días de edad en 50mL de medio MS líquido adicionado con 1.5mg/L de 2,4-D. Después de 35 días, los cultivos en suspensión celular iniciados con callos de 35 días, mostraron mayor crecimiento celular con una biomasa máxima de 194.9g/L de peso fresco y 6.59g/L de peso seco y 17.3% de volumen empacado. La fase exponencial finalizó al día 35 en los cultivos iniciados con callos de 35 días, y al día 21 en los cultivos iniciados con callos de 60 días. Dos fuentes de carbono fueron evaluadas: sacarosa y maltosa. La producción de mayor biomasa fue obtenida con sacarosa. Los cultivos celulares se establecieron con callos de 35 días cultivados en medio MS con la misma concentración de 2,4-D utilizada para la inducción de callos y 30g/L de sacarosa. Este medio fue considerado el óptimo para el mantenimiento y crecimiento de suspensiones celulares en ambas especies con subcultivos cada 20 días. El potencial biotecnológico para la producción de compuestos bioactivos en estas especies, para aplicaciones farmacológicas, agrícolas e industriales está siendo evaluado.
Palabras clave: Jatropha curcas, Jatropha gossypifolia, callos, cultivos de suspensión celular, cinética de crecimiento.
Jatropha curcas L. (physic nut, purging nut), a tropical plant belonging to the Euphorbiaceae family, is cultivated mainly as a hedge in many Latin American, Asian and African countries (Haas & Mitterbach 2000, Aiyelaagbe et al. 2007). It is a multipurpose tree of significant economic importance because of its industrial and medicinal uses. Fresh and decoction preparations from seeds, leaves, and bark are used in traditional medicine and for veterinary purposes (Devappa et al. 2010, Nayak & Patel 2010). The oil obtained from the seeds is used in the soap industry and as a substitute for diesel after transesterification; while the press cake is used as an animal feedstock or for biogas production (Martin & Mayeux 1984, Staubmann et al. 1997, Gübitz et al. 1997, Gübitz et al. 1999). During the past years, many large-scale cultivation projects have been developed to produce Jatropha seed oil as feedstock. More recently, Kalimuthu et al. (2010) reported antifungal activity on the methanol extract of leaf-derived callus of Jatropha curcas.
The biological significance of Jatropha proteins and peptides, along with its nutritional and therapeutic applications, will open avenues for new applications of proteins present in Jatropha (Devappa et al. 2010). Different pharmacological activities have been reported for this plant, including antibacterial (Aiyelaagbe et al. 1998, Aiyelaagbe et al. 2007), antitumor (Morris et al. 1970, Lin et al. 2003, Luo et al. 2007), antimicrobial (Aiyelaagbe et al. 2000, Kalimuthu et al. 2010), anti-inflammatory, antifungal (Wei et al. 2005, Jin-xia et al. 2005, Kalimuthu et al. 2010), antiprotozoal (Schmeda-Hirschmann et al. 1996), procoagulant and anticoagulant in latex (Cano & Plumbley 1989, Osoniyi & Onajobi 2003).
Cell suspension cultures offer an in vitro system that can be used as a tool for various studies in J. curcas and J. gossypifolia. They can be used in experiments involving mutant selection, mass propagation, protoplast isolation, gene transfer, cell biology to study cell wall traits (Mazarei et al. 2011), and for the large scale culturing of plant cells from which secondary metabolites can be extracted (Mulabagal & Tsay 2004). Cell suspension cultures provide an unlimited supply of uniform cells, which are grown in liquid culture. In comparison with whole plant systems, which have a relatively longer growth cycle as well as complex tissue specific proteomes, cells in suspension have a relatively shorter life cycle and remain undifferentiated. The shorter life cycle provides a continuous supply of experimental units, which are grown under tightly controlled environmental conditions, thus increasing reproducibility within and between experiments (Ngara et al. 2008). Additionally, a two stage culture regime is often used for the production of secondary metabolites from plant cells. The first stage involves growing and maintaining the cells at a high density on a standard maintenance medium, and the second stage involves the transfer of these cells into a production medium to obtain secondary metabolites (WaiLeng & Lai-Keng 2004).
A limited number of in vitro culture studies have been reported on the formation of calli (Kalimuthu et al. 2010, Kumar et al. 2008, Attaya et al. 2012), and cell suspension cultures (Soomoro & Memon 2007, Elfahmi & Ruslan 2011) of Jatropha curcas, but the accessions used for developing propagation protocols mainly originated from Asia, where the genetic diversity of the J. curcas is limited (Soomro & Memon 2007, Attaya et al. 2012). Further optimization of in vitro culture conditions, and the implementation of American accessions will be required to develop commercially viable in vitro propagation methods for local J. curcas (Attaya et al. 2012) and other Jatropha species. The origin of a plant that is used for in vitro cultivation is a critical factor that will determine the cultivation requirements and the protocol needed for a successful in vitro culturing (Attaya et al. 2012). Regeneration in J. curcas is also reported to be highly genotype dependent (da Camara et al. 1997, Sharma et al. 2011, Siang et al. 2012).
The objective of this study was to complete the establishment and long-term maintenance of calli and cell suspension cultures of local J. curcas and J. gossypifolia, in order to allow future studies on novel compounds for pharmaceutical and/or industrial applications.
Materials and Methods
The experiments were developed from January 2009 to December 2011.
Plant material: Calli induction experiments were conducted with two types of explants taken from in vitro or greenhouse germinated plants. Hypocotyl segments were used for J. curcas and J. gossypifolia, leaf tissue was used for J. curcas calli induction, but not for J. gossypifolia (due to previous results with J. curcas).
Leaves were washed with distilled water for five minutes, followed by a Tween 20 solution (three drops/100mL). They were then placed in a carbendazine solution (0.5%) for five minutes, and washed three times with sterile distilled water. Afterwards, leaves were disinfected with a sodium hypochlorite solution (0.1%) for five minutes, and finally washed three times with sterile distilled water.
Seeds were placed in a water and commercial soap solution and shaken for 24h. Next, they were washed in a Tween 20 solution for 10 minutes, followed by a carbendazine solution (1%) for 30 minutes with agitation. In a laminar flow hood, the seeds were placed in 80% ethanol solution for five minutes with agitation; finally, they were transferred to a sodium hypochlorite solution (35%) for 15 min. All seeds were washed three times with sterile distilled water in between all steps.
Germination of zygotic embryos: Zygotic embryos were extracted from the disinfected seeds and placed on basal media (BMS) supplied with 2mg/L gibberellic acid. Embryos were kept in darkness for one week, then transferred to BMS and maintained an extra week with a photoperiod of 16/8h (light/dark).
Calli induction: Composition of the basal media (BMS): Explants were placed in contact with semisolid MS media (Murashige & Skoog 1962) supplemented with 30g/L (w/v) sucrose and 0.7% agar. All medium pH was adjusted to 5.8 before autoclaving (120°C for 20min).
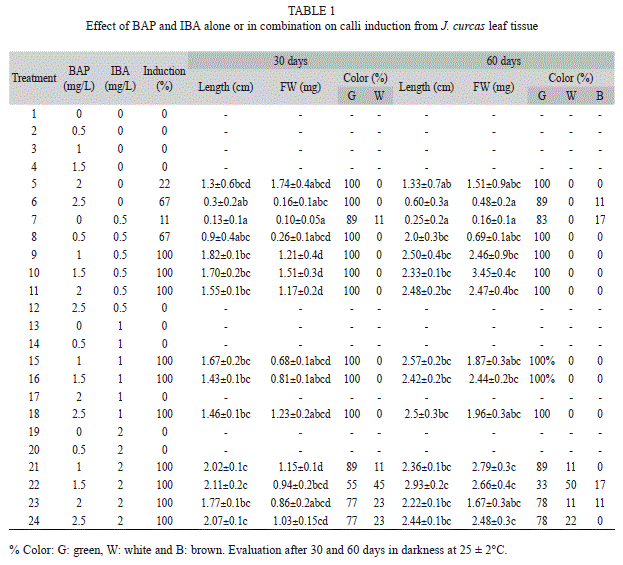
Calli induction from leaf tissue: We used leaves of J. curcas germinated in greenhouse. Leaf disks were cut with a 10mm diameter hole punch and placed with the abaxial side in contact with BMS supplemented with IBA (indole-3-butyric acid) at 0.5 up to 2.0mg/L, and BAP (6-Benzylaminopurine) at 0.5 up to 2.5mg/L. Growth regulators were used separately and in combination. A total of three explants per jar and five repetitions per treatment were used; all treatments were incubated in darkness for four weeks.
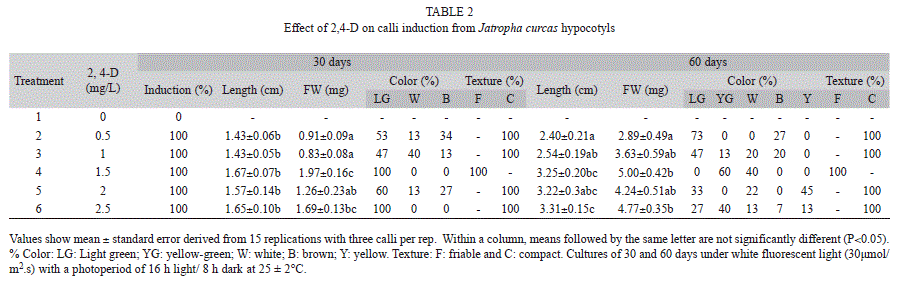
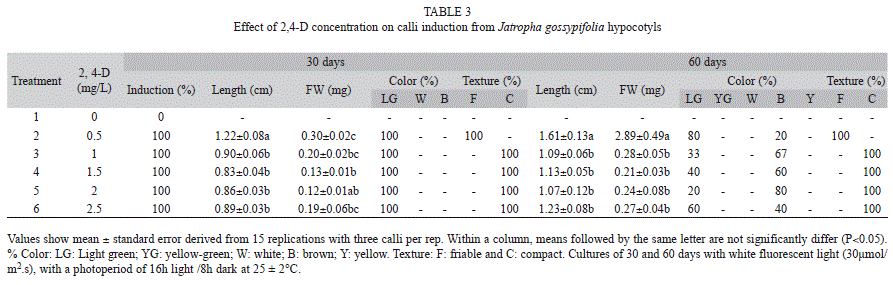
Calli induction from hypocotyl segments: Hypocotyls from 15-day-old in vitro germinated J. curcas and J. gossypifolia plants were used for calli induction. Five millimeter segments were placed in BMS supplied with 2,4-D (dichlorophenoxyacetic acid) at 0, 0.5, 1, 1.5, 2 or 2.5mg/L. Explants were incubated at 25±2°C with a photoperiod of 16h with white fluorescent light (30μmol/m2.s) and 8h darkness. A total of three explants were used per jar with five repetitions per treatment.
Calli growth kinetics: Calli fresh weight (g), length (cm), color and texture, and calli growth kinetics were evaluated during the induction phase, and the best treatment for the establishment of cell suspensions was selected.
Cell suspension culture media (CSM): Liquid MS media (Murashige & Skoog 1962) with sucrose (30g/L-w/v), myo-inositol (100mg/L) and thiamine-HCL (10mg/L), was supplemented with 2,4-D (1.5mg/L for J. curcas and 0.5mg/L for J. gossypifolia). The pH was adjusted to 5.8 prior to autoclave sterilization (120°C for 20 min).
Establishment of J. curcas cell suspensions: Effect of two calli ages: Two calli ages were evaluated for J. curcas (35 and 60 DAI, days after induction), and one for J. gossypifolia (35 DAI). For this, 50mL CSM was inoculated with 1g of fresh weight (FW) of friable calli and incubated in 250mL Erlenmeyer flasks. Suspensions were incubated at 25±2°C with a photoperiod of 12h light and 12h darkness in an orbital rotary shaker (160rpm). Three replications per treatment were used.
Cell suspension growth kinetics: Fresh weight (FW), dry weight (DW) and packed cell volume (PCV) were measured every seven days until day 49. Fresh weight was obtained by removing the suspension, and dry weight was obtained after drying the sample in the oven for 24h at 80°C. To determine PCV, the cell suspension culture sample was homogenized and 10mL were placed in a 50mL graduate cylinder and were allowed to settle for 15min. The volume occupied by the cells was measured. Three measurements per replicate were done.
The exponential growth of the cells in suspension was calculated using the equation: ln X/X0=m(t-t0), where X0 is the cell concentration (g/L) at time t0, when exponential growth begins, and X is the cell concentration at time t. The doubling time was calculated using the equation: td=(ln2/m), and the growth index according to: IC=(X-X0)/X0 (Rodríguez-Monroy 2007). The m, specific growth rate (over time-1), rate at which growth occurs; IC, growth index; td: doubling time.
Effect of initial concentration of sucrose and maltose on cell cultures: The effect of sucrose or maltose was compared in cell cultures established from calli and cultures started from the established suspensions. The carbon source of the previously described CSM was evaluated: sucrose and maltose at 20, 30, 40 and 50 (g/L-w/v). Three replications per treatment were done. Liquid MS media (Murashige & Skoog 1962) supplied with 100mg/L myoinositol, 10mg/L thiamine-HCL and 1.5mg/L 2, 4-D was inoculated with fresh cells of each cell culture type (one gram fresh weight/25mL media) in 250mL Erlenmeyer flasks. Fresh weight (FW) and dry weight (DW) were evaluated after 60 days.
Determination of sugar content and yield: Sugar content of the media was determined according to the method proposed by Dubois et al. (1956) with some modifications. Briefly, cell suspension were filtered and diluted (1:100); 100μL of the dilution was placed in a 20mL tube, then 1mL of phenol (5%) and 5mL of concentrated H2SO4 were added and mixed. Samples were analyzed with a spectrophotometer at 490nm. Zero was adjusted using 100μL of distilled water instead of sample. Total sugar (g/L) was calculated using the calibration curve described by Rodríguez-Monroy (2007). Cell yield was calculated with the equation Yx/s=(X-X0)/(S0-S), where S0 is the concentration of the initial carbon source (g/L) and S is the concentration at time t.
Establishment of J. gossypifolia cell suspensions: Cell suspensions of J. gossypifolia were established by inoculating 1g of friable 35-day-old calli into 50mL of CSM media with 0.5mg/L 2,4-D.
Descriptive analysis and analysis of variance (ANOVA) were used to determine significant differences between treatments, using the statistical package STATISTICA (1998). A completely random design was used for callus induction studies, and a factorial design with a completely random arrangement was used for cell suspension studies. In the callus induction studies, percent induction, morphology, texture, length (cm), fresh weight (g), dry weight (g) and color were evaluated and photographs were made. For the cell suspensions, the following variables were evaluated: percent callus disaggregation, suspension color, number of days required for disaggregation and growth kinetics.
Results
Calli induction from leaf tissue: No calli from J. curcas were formed in the absence of growth regulators. Calli formed in BMS supplied with BAP at 2.0-2.5mg/L, but no calli were formed at lower concentrations (0.5-1mg/L). Best results for calli induction were obtained with both regulators: 100% of the explants produced calli when the media was supplied with IBA at 0.5 to 2.0mg/L and BAP at 0.5 to 2.5mg/L. However, calli formed were always compact (Table 1), and did not disaggregate in the cell suspension (data not shown), and as such they were not suitable for the establishment of cell suspensions. Given these results, J. gossypifolia was not evaluated with this treatment.
Calli induction from hypocotyl segments: Calli induction was 100% in explants of both species 30 days after application of treatments with concentrations from 0.5-2.5mg/L of 2,4-D. There were no calli formed in the absence of the regulator. In all treatments with 2,4-D, J. curcas calli were formed, but only in presence of 1.5mg/L of 2,4-D, yellowish-green calli classified as friable were observed (Table 2, Fig. 1). The texture of J. gossypifolia calli treated with 1 to 2.5mg/L of 2,4-D were not friable, but calli treated with 0.5mg/L were classified as friable (Table 3). After 60 days, the calli treated with 0.5mg/L of 2,4-D which remained friable.
The growth kinetics of J. curcas calli cultured in 1.5mg/L 2,4-D showed continuous growth for DW and FW during the evaluation period. J. gossypifolia cultured in 0.5mg/L 2,4-D showed greater length and calli weight at 60 days of culture, with significant differences between treatments (Table 3). However, within a single callus there were both friable and compact areas (Fig. 1). Subcultures were therefore made every 20 days in order to obtain greater growth of friable calli (data not shown).
Establishment of J. curcas cell suspensions: Calli formed in 1.5mg/L 2,4-D were used to initiate cell suspensions of J. curcas. The texture of calli formed at this concentration were friable, although calli length were not significantly different from other treatments (Table 2). The J. curcas cells grew well in liquid MS media with the same concentration of growth regulator used for calli induction (1.5mg/L 2,4-D).
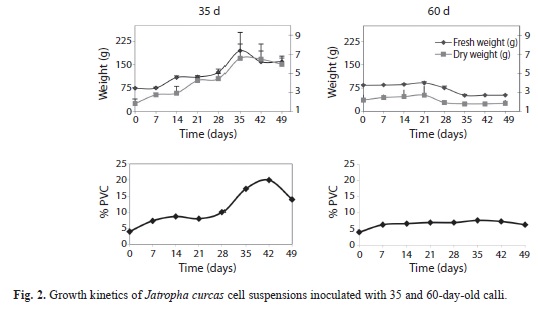
Effect of two calli ages: Growth kinetics were similar for FW, DW (g/L) and PCV of suspensions established with 35-day-old calli. The three variables showed a lag phase from 0-7 days, an exponential growth phase from 7-35 days and a stationary phase from 35-49 days. FW (194.90g/L) and DW (6.59g/L) were greatest at 35 days (Fig. 2).
Growth kinetics for FW and DW of suspensions initiated with 60-day-old calli did not show a lag phase, but showed an exponential phase after 21 days and a stationary phase afterwards from 21-49 days. Maximum FW (91.58g/L) and DW (2.71g/L) were observed on day 21 (Fig. 2).
In the cell culture inoculated with 35-day- old calli, total callus disaggregation was observed on the first day. The appearance of the suspension was translucent from day 0-7, whitish from day 14-28 and cream-colored from day 35-49. Thick cellular aggregates were observed after 35 days of culture. In contrast, the growth curve for suspensions established with the 60-day-old calli, did not show distinct growth phases and the PCV were lower (Fig. 2). In the case of the suspensions established with 35-day-old calli, the exponential growth phase was observed until day 35, and the specific growth rate (measured as divisions per day) was µ=0.22; and the suspensions established with 60-day-old calli, showed the exponential phase until day 21 with a specific growth rate (divisions per day) of µ=0.02 (Fig. 2).
Time required to double the biomass of the initial inoculum (td) was 3.2 and 28.4 days for cultures initiated with 35 and 60 day-old calli, respectively. The increase in cell concentration (IC) was 2.62 for cultures started with 35-day- old calli, and 0.24 for cultures started with 60-day-old calli.
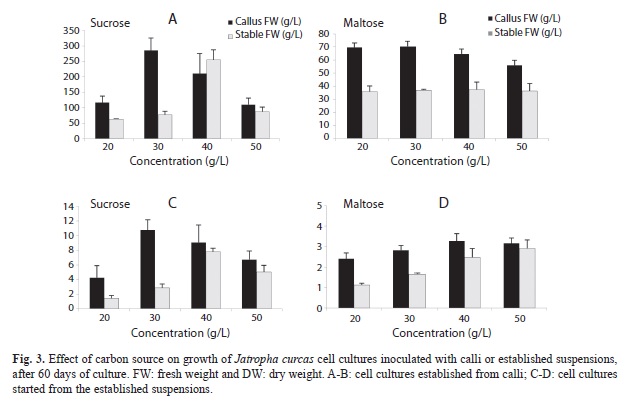
Effect of initial concentration of sucrose and maltose on cell cultures: The fresh biomasses of calli-inoculated cell cultures grown at different sucrose concentrations were between 110.59 and 285.33g/L (FW), no differences were found between treatments (Fig. 3). In contrast, the biomasses of cultures inoculated with suspensions were between 63.48 and 254.94g/L (FW). The highest fresh weight was obtained with the 40% sucrose treatment (254.9g/L) (Fig. 3). Dry weight in calli-inoculated cultures was between 4.26-10.76g/L (DW), with no significant differences between treatments. Dry weight of cultures inoculated with established suspensions was between 1.41 and 7.78g/L (DW), with significant differences between treatments (Fig. 3). The PCV was 43.33% in the 30g/L sucrose treatment, and 28% with 40g/L sucrose. Calli-inoculated cultures grown with different maltose concentrations showed no significant differences for FW (55.99-70.37g/L) or DW (2.41-3.27g/L) (Fig. 3). PCV was greater (8.33%) with 30g/L maltose, but this was not greater than the biomass produced with 30g/L of sucrose. Differences between DW of established cultures grown at different maltose concentrations were observed (between 1.12 and 2.92g/L), no differences were detected when FW parameter was used (between 35.82 and 37.36g/L) (Fig. 3). In general terms, differences between cultures inoculated with calli or established suspensions grown with sucrose were observed, higher fresh and dry biomass values were observed in calli-inoculated cultures.
Cultures with maltose, inoculated with calli or suspensions, showed significant differences for fresh and dry biomass. Calli-inoculated cultures showed greater values (FW 65.17 and DW 2.04g/L). This was expected, as thick cell aggregates were present in calli-inoculated cultures. In established cultures (FW 36.60 and DW 2.92g/L), only fine cell aggregates were present (Fig. 3).
Cultures inoculated with calli and supplied with sucrose had higher fresh and dry biomass values than those supplied with maltose (sucrose=FW 180.94 and DW 7.71g/L, maltose=FW 65.17 and DW 2.92g/L). These results were also obtained with cultures inoculated with cell suspensions (sucrose=FW 117.62 and DW 4.35g/L, maltose=FW 36.60 and DW 2.04g/L). This suggests that sucrose can be used as a carbon source in cell cultures of Jatropha.
Determination of sugar content and yield: Cell yield in the J. curcas established from cell suspensions was 0.08g/L with 20% sucrose, 0.95g/L with 30%, 1.13g/L with 40% and 1.01g/L for cultures with 50% sucrose. This means that more sucrose was consumed than produced in the 20% sucrose treatment; slightly more biomass was produced than the amount of sucrose consumed within the 40% treatment. Finally, production and consumption was nearly equal in the 30 and 50% treatments. Cell yield in the cultures treated with maltose was 0.012, 0.108, 0.113 and 0.239 at 20, 30, 40 and 50% of carbon source. These results showed that maltose consumption was greater than biomass production.
Establishment of J. curcas cell line: J. curcas cell suspensions were established from 35-day-old calli inoculated in CSM media with 30g/L sucrose in flasks with agitation. This cell line has been maintained by continuous subculturing every 20 days in 250mL Erlenmeyer flasks with 50mL CSM media. Flasks were maintained on an orbital rotary shaker at 160rpm at 25 ± 2°C with a photoperiod of 12h light and 12h darkness.
Establishment of J. gossypifolia cell line: Calli obtained in the 0.5mg/L 2,4-D treatment were used to initiate cell suspensions, because calli texture were 100% friable and their length and weight were significantly greater than in other treatments (Table 3). Cells of J. gossypifolia grew well in liquid MS media with the same concentration of growth regulator (1.5mg/L 2,4-D). A total calli disaggregation was observed the first day.
Discussion
In this research, J. curcas calli were obtained from leaf tissue grown in BAP (1.0-2.5mg/L) and IBA (0.5-2.0mg/L); however, calli were compact and not adequate for the establishment of cell suspensions. The combination of BAP and IBA has been reported in the direct induction of adventitious shoots from leaf segments of J. integerrima (Sujatha & Mukta 1993, Sujatha et al. 2005, Deore & Johnson 2008), and indirect organogenesis of J. curcas (Sujatha & Mukta 1996, Weida et al. 2003, Ajay & Sudhakar 2008, Misra et al. 2010).
Using hypocotyl segments from J. curcas and J. gossypifolia plants germinated in vitro, friable calli were induced in all explants in the presence of 1.5mg/L 2,4-D. This coincides with results obtained by Soomro & Memon (2007), who reported 100% induction of friable calli from J. curcas hypocotyl with 0.5mg/L, and 40% induction of compact calli using leaf tissue. In our research, no calli development were observed from leaf tissue at 2,4-D concentrations from 0.5 to 2.5mg/L (data not shown). In other research, 2,4-D has been used for the induction of friable calli for the establishment of cell suspensions. Varisai et al. (2004) used 2.0mg/L of 2,4-D in leaf tissue segments of Macrotyloma uniflorum (Lam.) Verdc. and obtained 48.6% of calli induction. Khafagi (2007) obtained calli for cell suspensions establishment of Peganum harmala using 0.5mg/L of 2,4-D; while, Deo et al. (2009) obtained 20.4% calli formation for Colocasia esculenta var. esculenta using 0.5-1mg/L of 2,4-D.
In J. curcas, yellow-green calli with friable texture were formed from hypocotyl segments cultured in 1.5mg/L 2,4-D. This treatment was therefore used to initiate cell cultures. Although calli showed continuous growth over time, at 60 days, the morphology and color were characteristic of non-friable calli. Conditions necessary for the establishment of cell suspensions were maintained by subculturing every 20 days, with selection of sections of friable calli.
Initially, J. curcas cell suspensions were established in liquid MS media with the same concentration used for calli induction (1.5mg/L of 2,4-D). One gram of calli were used for inoculation since frequently, if larger calli (2g) were used, the cell suspension became necrotic and growth decreased dramatically, and if less than 1g was used, cell division and proliferation did not begin (Ben Amar et al. 2007).
The effect of inoculum age on the establishment of J. curcas cell suspensions was evaluated using 35 and 60-day-old calli. Soomro & Memon (2007) established Jatropha curcas suspensions from 28-day-old calli inoculated to media supplied with 0.5mg/L of 2,4-D. Other authors have also established cell suspensions of other species using media with 2,4-D (Doelling & Pikaard 1993, Mythili et al. 1999, Lee & Chan 2004). Results of this study showed that growth of cell suspensions was influenced by inoculum age, with growth kinetics showing higher values for suspensions initiated with 35-day-old calli. Use of 60-dayold calli had a negative effect on growth, as lower values were observed for cell biomass, as determined by FW, DW and PCV. In a study of the influence of calli age on the establishment of cell suspensions of coffee, González et al. (2010) observed greater cell growth with 28 and 35-day-old calli, and a decrease in growth with increased calli age. This may be because younger tissues have higher growth and proliferation rates than older tissues.
Growth comparison of cell suspensions showed that inoculation with 35-day-old calli resulted in greater growth when compared to cultures inoculated with 60-day-old calli, with a difference of 103.32g/L in FW and 3.88g/L in DW, and at day 35 a specific growth rate of 0.22 divisions per day. Soomro & Memon (2007) also reported greater growth in Jatropha curcas suspensions at day 21.
Results of this study indicated that sucrose should be used as a carbon source in Jatropha cell cultures inoculated with calli or for established cell suspensions. Sucrose is the main carbohydrate synthesized by plants and has been found to increase production of fresh biomass. Sugars function both as a carbon source and as an osmotic regulator in culture media. Cell growth depends on the utilization of carbon for the formation of principal cell components and as an energy source. The use of 30g/L sucrose as a carbon source is indicated, although treatments with 40% sucrose showed the highest FW and DW values. An increase in sugars in the culture media increases the production of secondary metabolites in many plant cell cultures (Zhang et al. 1996), and may also be associated with plasmolysis of plant cells, and a resulting decrease in the biosynthesis of secondary metabolites (Chattopadhyay et al. 2003).
Based on results obtained with J. curcas cultures, cell cultures were established for both J. curcas and J. gossypifolia. These cultures were inoculated with 35-day-old calli in MS media supplemented with the same concentration of 2,4-D used for calli induction. To date, these cell lines have been maintained by subculturing every 20 days, using a 1mm x 1mm filter to separate thick aggregates and obtain cell suspensions with only fine aggregates. The fine aggregates were subcultured in 250mL flasks with 50mL CSM media, using 10% PCV as initial inoculum. Suspensions were incubated with a photoperiod of 12h light, 12h darkness in an orbital rotary shaker (160rpm) at 25 ± 2°C. The thick aggregates were stored at -4°C for future studies, as their use as inoculum did not allow growth of fine cellular aggregates in cell suspensions (data not shown).
Cell suspensions are used for the generation of large quantities of cells for quantitative and qualitative analyses of growth and metabolism of new compounds, as well as for studies of the cell cycle and systems of regeneration of plants under standardized conditions (Torabi et al. 2008). Cell suspensions are also suitable for protoplast isolation in studies of transitory genetic expression and transformation by Agrobacterium (Torabi et al. 2008). In our case, cell lines of Jatropha curcas and Jatropha gossypifolia, both native to Costa Rica, will be used for future bioprospection of secondary metabolites of agronomic, pharmacological and industrial interest.
Acknowledgement
The authors wish to thank to the Vicerrectoría de Investigación de la Universidad de Costa Rica for funding Project 111-A9-097; Inversiones J.S.S.A. de Costa Rica for providing Jatropha plant material; Secretaría de Relaciones Exteriores de México and CENIBiot, Costa Rica for financial assistance with Project 301CR152; Mario Rodríguez Monroy, CEPROBI, for training Solís in establishment of suspension cultures and determination of growth variables, and for collaboration during the development of this research; Centro de Investigación en Biología Celular y Molecular (CIBCM, UCR) for the use of the rotary orbital shaker; Centro de Investigación en Ciencias del Mar y Limnología (CIMAR-UCR) for the use of the spectrophotometer; Luis Salazar Figueroa, Laboratorio de Nematología, Escuela de Agronomía, UCR for greenhouse space; Elmer G. García Díaz, for providing Jatropha gossypifolia plant material; and Escuela de Biología, Sección de Genética for partial funding for a student assistant ship. The authors wish to acknowledge Marc Berthouly, CIRAD, Montpellier, France, for manuscript revision.
References
Aiyelaagbe, O.O., K. Adesogan, O. Ekundayo & A. Hassanali. 1998. Antifeedant activity of Jatropha podagrica roots. Fitoterapia 69: 175-176. [ Links ]
Aiyelaagbe, O.O., K. Adesogan, O. Ekundayo & B.A. Adeniyi. 2000. The antimicrobial activity of roots of Jatropha podagrica (Hook). Phytother. Res. 14: 60-62. [ Links ]
Aiyelaagbe, O.O., K. Adesogan, O. Ekundayo & J.B. Gloer. 2007. Antibacterial diterpenoids from Jatropha podagrica Hook. Phytochemistry 68: 2420-2425. [ Links ]
Ajay, D. & T. Sudhakar. 2008. High-frequency plant regeneration from leaf-disc cultures of Jatropha curcas L.: an important biodiesel plant. Plant Biotechnol. Rep. 2: 7-11. [ Links ]
Attaya, A.S., D. Geelen & A.E.H. Belal. 2012. Progress in Jatropha curcas tissue culture. American-Eurasian J. Sustain. Agric. 6: 6-13. [ Links ]
Ben, A., P. Cobanov, K. Boonrod, G. Krczal, S. Bouzid, A. Ghorbel & G. Reustle. 2007. Efficient procedure for grapevine embryogenic suspension establishment and plant regeneration: role of conditioned medium for cell proliferation. Plant Cell Rep. 26: 1439-1447. [ Links ]
Cano, L.M. & R.A. Plumbley. 1989. Purification and partial characterization of a hemagglutinin from seeds of Jatropha curcas. J. Food Biochem. 13: 1-20. [ Links ]
Chattopadhyay, S., R.S. Mehra, A.K. Srivastava, S.S. Bhojwani & V.S. Bisaria. 2003. Effect of major nutrients on podophyllotoxin production in Podophyllum hexandrum suspension cultures. Appl. Micro. Biotechnol. 60: 541-546. [ Links ]
da Camara, M.A., N. Frick, R. Kremen, H. Katinger & M. da Camara. 1997. Biotechnological Approaches to the Improvement of Jatropha curcas. Proceedings of the International Symposium on Jatropha, Managua, Nicaragua, 15. [ Links ]
Deo, P., R. Harding, M. Taylor, A. Tyagi & D. Becker. 2009. Somatic embryogenesis, organogenesis and plant regeneration in taro (Colocasia esculenta var. esculenta). Plant Cell Tiss. Org. 99: 61-71. [ Links ]
Deore, A.C. & T.S. Johnson. 2008. High-frequency plant regeneration from leaf-disc cultures of Jatropha curcas L.: an important biodiesel plant. Plant Biotechnol. Rep. 2: 10-15. [ Links ]
Devappa, R.K., H.P.S. Makkar & K. Becker. 2010. Nutritional, Biochemical, and Pharmaceutical Potential of Proteins and Peptides from Jatropha: Review. J. Agric. Food Chem. 58: 6543-6555. [ Links ]
Doelling, J. & C. Pikaard. 1993. Transient expression in Arabidopsis thaliana protoplasts derived from rapidly established cell suspension cultures. Plant Cell Rep. 12: 241-244. [ Links ]
Dubois, M., K. A. Gilles, J.K. Hamilton, P.A. Rebers & F. Smith. 1956. Colorimetric method for determination of sugars and related substances. Analytical Chem. 28: 350-356. [ Links ]
Elfahmi, A. & Ruslan, K. 2011. Phytochemical study of Jatropha curcas cell culture. Biotropia 18: 42-49. [ Links ]
González, M., M. Hernández & A. Hernández. 2010. Influencia de la edad del callo en la inducción de suspensiones celulares de cafeto. Agr Mesoamericana 21: 299-306. [ Links ]
Gübitz, G.M., M. Mittelbach & M. Trabi. 1997. Biofuels and Industrial Products from Jatropha curcas. Developed from the Symposium “Jatropha 97”. Managua, Nicaragua, February 23-27. Sponsored by the Austrian Ministry of Foreign Affairs and by Sucher & Holzer, Alberstraße 4, A-8010 Graz, Austria, 190-205. [ Links ]
Gübitz, G., M. Mittelbach & M. Trabi. 1999. Exploitation of the tropical oil seed plant Jatropha curcas L. Bior. Technol 67: 73-82. [ Links ]
Haas, W. & M. Mittelbach. 2000. Detoxification experiments with the seed oil from Jatropha curcas L. Ind. Crops Prod. 12: 111-118. [ Links ]
Jin-xia, Z., W. Qin, Z. Li-jun, L. Guo-jin, C. Yu, T. Lin & C. Fang. 2005. The extraction of β-1, 3-glucanase and analysis of partial characteristics in Jatropha curcas. South West China J. Agric. Sci. 18: 328-333. [ Links ]
Kalimuthu, K., S. Vijayakumar & R. Senthilkumar. 2010. Antimicrobial activity of the biodiesel plant, Jatropha curcas L. Int. J. Pharma. Bio. Sci. 1: 1-6. [ Links ]
Khafagi, I. 2007. Generation of Alkaloid-containing Idioblasts During Cellular Morphogenesis of Peganum harmala L. Cell Suspension Cultures. Am. J. Plant Physiol. 2: 17-26. [ Links ]
Kumar, N., S.D.V.N. Pamidimarri, M. Kaur, G. Boricha & M.P. Reddy. 2008. Effects of NaCl on growth, ion accumulation, protein, proline contents and antioxidant enzymes activity in callus cultures of Jatropha curcas. Biologia 63: 378-382. [ Links ]
Lee, W. & L. Chan. 2004. Establishment of Orthosiphon stamineus Cell Suspension Culture for Cell Growth. Plant Cell Tiss. Organ 78: 101-106. [ Links ]
Lin, J., F. Yan, L. Tang & F. Chen. 2003. Antitumor effects of curcin from seeds of Jatropha curcas. Acta Pharmacol. Sin. 24: 241-246. [ Links ]
Luo, M.J., W.X. Liu, X.Y. Yang, Y. Xu, F. Yan, P. Huang & F. Chen. 2007. Cloning, Expression, and Antitumor Activity of Recombinant Protein of Curcin. Russ. J. Plant Physiol. 54: 202-206. [ Links ]
Martin, G. & A. Mayeux. 1984. Réfléxions sur les cultures olagineuses, energétiques II.-Le Pourghere (Jatropha curcas L.): un carburant possible. Olagineux 39: 283-287. [ Links ]
Mazarei, M., H. Al-Ahmada, M.R. Rudis, B.L. Joyce & C.N. Jr. Stewart. 2011. Switchgrass (Panicum virgatum L.) cell suspension cultures: Establishment, characterization, and application. Article in press. Plant Science. [ Links ]
Misra, P., N. Gupta, D. Toppo, Pande, V., Mishra, M. & Tuli, R. 2010. Establishment of long-term proliferating shoot cultures of elite Jatropha curcas L by controlling endophytic bacterial contamination. Plant Cell Tiss. Organ 100: 189-197. [ Links ]
Morris, S., C.W. Sigel, M.J. Matz, J.A. Saenz, R.C. Haltiwanger & R.F. Bryan. 1970. Jatrophone, a Novel Macrocyclic Diterpenoid Tumor Inhibitor from Jatropha gossypiifolia. J. Am. Chem. Soc. 92: 14. [ Links ]
Mulabagal, V. &. H. S. Tsay. 2004. Plant Cell Cultures An Alternative and Efficient Source for the Production of Biologically Important Secondary Metabolites. Int. J. Appl. Sci. Eng. 2: 29-48. [ Links ]
Murashige, T. & F. Skoog. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 15: 473-479. [ Links ]
Mythili, P., N. Seetharama & V. Reddy. 1999. Plant regeneration from embryogenic cell suspension cultures in wild species of sorghum (Sorghum dimidiatum Stapf). Plant Cell Rep. 18: 424-428. [ Links ]
Nayak, B.S. & K.N. Patel. 2010. Pharmacognostic studies of the Jatropha curcas leaves. Int. J. Pharm.Tech. Res. 2: 140-143. [ Links ]
Ngara, R., J. Rees & B.K. Ndimba. 2008. Establishment of sorghum cell suspension culture system for proteomics studies. Afr. J. Biotechnol. 7: 744-749. [ Links ]
Osoniyi, O. & F. Onajobi. 2003. Coagulant and anticoagulant activities in Jatropha curcas latex. J. Ethnopharmacol. 89: 101-105. [ Links ]
Rodríguez-Monroy, M. 2007. Técnicas para el análisis de muestras y determinación de variables de crecimiento y cálculo de parámetros cinéticos. Manual de prácticas del curso Aspectos Teórico Prácticos del cultivo de células vegetales en biorreactores. Instituto Politécnico Nacional, CEPROBI y la Sociedad Mexicana de Biotecnología y Bioingeniería A. C., Morelos, México, 14-28. [ Links ]
Schmeda-Hirschmann, G., I. Razmilic, M. Sauvain, C. Moretti, V. Muñoz, E. Ruiz, E. Balanza & A. Fournet. 1996. Antiprotozoal activity of Jatrogrossidione from Jatropha grossidentata and Jatrophone from Jatropha isabellii. Phytother. Res. 10: 375-378. [ Links ]
Sharma, S., N. Kumar & M. Reddy. 2011. Regeneration in Jatropha curcas: Factors affecting the efficiency of in vitro regeneration. Ind. Crop. Prod. 34: 943-951. [ Links ]
Siang, T., S. Soong & A. Yien. 2012. Plant regeneration studies of Jatropha curcas using induced embryogenic callus from cotyledon explants. Afr. J. Biotechnol. 11: 8022-8031. [ Links ]
Soomro, R. & R.A. Memon. 2007. Establishment of callus and suspension culture in Jatropha curcas. Pakistan J. Bot. 39: 2431-2441. [ Links ]
Staubmann, R., G. Foidl, N. Foidl, G. Gübitz, R. Lafferty, V. Valencia & W. Steiner. 1997. Biogas Production from Jatropha curcas Press-Cake. Appl. Biochem. Biotechnol. 63: 457-467. [ Links ]
Sujatha, M. & D. Mukta. 1993. Rapid plant regeneration from various explants of Jatropha integerrima. Plant Cell Tiss. Organ 35: 293-296. [ Links ]
Sujatha, M., H. Makkar & K. Becker. 2005. Shoot bud proliferation from axilary nodes and leaf sections of non-toxic Jatropha curcas L. Plant Growth Regul. 47: 83-90. [ Links ]
Sujatha, M. & N. Mukta. 1996. Morphogenesis and plant regeneration from tissue cultures of Jatropha curcas. Plant Cell Tiss. Organ 44: 135-141. [ Links ]
Torabi, F., A. Majad & A. Ehsanpour. 2008. Plant regeneration from cell suspension culture of potato (Solanum tuberosum L.). Pakistan J. Biol. Sci. 11: 778-782. [ Links ]
Varisai, S., C. Wang, M. Thiruvengadam & N. Jayabalan. 2004. In vitro plant regeneration via somatic embryogenesis through cell suspension cultures of horsegram [Macrotyloma uniflorum (lam.) verdc.]. In Vitro Cellular & Developmental Biology – Plant 40: 284-289. [ Links ]
Wai-Leng, L. & Ch. Lai-Keng. 2004. Establishment of Orthosiphon stamineus cell suspensión culture for cell growth. Plant Cell Tiss. Organ 78: 101-106. [ Links ]
Wei, Q., Y. Liao, Y. Chen, S.H. Wang, Y. Xu, L. Tang & F. Chen. 2005. Isolation, characterisation and antifungal activity of β-1, 3-glucanase from seeds of Jatropha curcas. South Afr. J. Botany 71: 95-9. [ Links ]
Weida, L., W. Qim, T. Lin, Y. Fang & C. Fang. 2003. Induction of callus from Jatropha curcas and its rapid propagation. Chin. J. Appl. Environ. Biol. 9: 127-130. [ Links ]
Zhang, Y.H., J.J. Zhong & J.T. Yu. 1996. Enhancement of ginseng saponin production in suspension cultures of Panax notoginseng: manipulation of medium sucrose. J. Biotechnol. 51: 49-56. [ Links ]
*Correspondencia a: Aiyelaagbe, O.O., K. Adesogan, O. Ekundayo & B.A. Adeniyi. 2000. The antimicrobial activity of roots of Jatropha podagrica (Hook). Phytother. Res. 14: 60-62. [ Links ]
Aiyelaagbe, O.O., K. Adesogan, O. Ekundayo & J.B. Gloer. 2007. Antibacterial diterpenoids from Jatropha podagrica Hook. Phytochemistry 68: 2420-2425. [ Links ]
Ajay, D. & T. Sudhakar. 2008. High-frequency plant regeneration from leaf-disc cultures of Jatropha curcas L.: an important biodiesel plant. Plant Biotechnol. Rep. 2: 7-11. [ Links ]
Attaya, A.S., D. Geelen & A.E.H. Belal. 2012. Progress in Jatropha curcas tissue culture. American-Eurasian J. Sustain. Agric. 6: 6-13. [ Links ]
Ben, A., P. Cobanov, K. Boonrod, G. Krczal, S. Bouzid, A. Ghorbel & G. Reustle. 2007. Efficient procedure for grapevine embryogenic suspension establishment and plant regeneration: role of conditioned medium for cell proliferation. Plant Cell Rep. 26: 1439-1447. [ Links ]
Cano, L.M. & R.A. Plumbley. 1989. Purification and partial characterization of a hemagglutinin from seeds of Jatropha curcas. J. Food Biochem. 13: 1-20. [ Links ]
Chattopadhyay, S., R.S. Mehra, A.K. Srivastava, S.S. Bhojwani & V.S. Bisaria. 2003. Effect of major nutrients on podophyllotoxin production in Podophyllum hexandrum suspension cultures. Appl. Micro. Biotechnol. 60: 541-546. [ Links ]
da Camara, M.A., N. Frick, R. Kremen, H. Katinger & M. da Camara. 1997. Biotechnological Approaches to the Improvement of Jatropha curcas. Proceedings of the International Symposium on Jatropha, Managua, Nicaragua, 15. [ Links ]
Deo, P., R. Harding, M. Taylor, A. Tyagi & D. Becker. 2009. Somatic embryogenesis, organogenesis and plant regeneration in taro (Colocasia esculenta var. esculenta). Plant Cell Tiss. Org. 99: 61-71. [ Links ]
Deore, A.C. & T.S. Johnson. 2008. High-frequency plant regeneration from leaf-disc cultures of Jatropha curcas L.: an important biodiesel plant. Plant Biotechnol. Rep. 2: 10-15. [ Links ]
Devappa, R.K., H.P.S. Makkar & K. Becker. 2010. Nutritional, Biochemical, and Pharmaceutical Potential of Proteins and Peptides from Jatropha: Review. J. Agric. Food Chem. 58: 6543-6555. [ Links ]
Doelling, J. & C. Pikaard. 1993. Transient expression in Arabidopsis thaliana protoplasts derived from rapidly established cell suspension cultures. Plant Cell Rep. 12: 241-244. [ Links ]
Dubois, M., K. A. Gilles, J.K. Hamilton, P.A. Rebers & F. Smith. 1956. Colorimetric method for determination of sugars and related substances. Analytical Chem. 28: 350-356. [ Links ]
Elfahmi, A. & Ruslan, K. 2011. Phytochemical study of Jatropha curcas cell culture. Biotropia 18: 42-49. [ Links ]
González, M., M. Hernández & A. Hernández. 2010. Influencia de la edad del callo en la inducción de suspensiones celulares de cafeto. Agr Mesoamericana 21: 299-306. [ Links ]
Gübitz, G.M., M. Mittelbach & M. Trabi. 1997. Biofuels and Industrial Products from Jatropha curcas. Developed from the Symposium “Jatropha 97”. Managua, Nicaragua, February 23-27. Sponsored by the Austrian Ministry of Foreign Affairs and by Sucher & Holzer, Alberstraße 4, A-8010 Graz, Austria, 190-205. [ Links ]
Gübitz, G., M. Mittelbach & M. Trabi. 1999. Exploitation of the tropical oil seed plant Jatropha curcas L. Bior. Technol 67: 73-82. [ Links ]
Haas, W. & M. Mittelbach. 2000. Detoxification experiments with the seed oil from Jatropha curcas L. Ind. Crops Prod. 12: 111-118. [ Links ]
Jin-xia, Z., W. Qin, Z. Li-jun, L. Guo-jin, C. Yu, T. Lin & C. Fang. 2005. The extraction of β-1, 3-glucanase and analysis of partial characteristics in Jatropha curcas. South West China J. Agric. Sci. 18: 328-333. [ Links ]
Kalimuthu, K., S. Vijayakumar & R. Senthilkumar. 2010. Antimicrobial activity of the biodiesel plant, Jatropha curcas L. Int. J. Pharma. Bio. Sci. 1: 1-6. [ Links ]
Khafagi, I. 2007. Generation of Alkaloid-containing Idioblasts During Cellular Morphogenesis of Peganum harmala L. Cell Suspension Cultures. Am. J. Plant Physiol. 2: 17-26. [ Links ]
Kumar, N., S.D.V.N. Pamidimarri, M. Kaur, G. Boricha & M.P. Reddy. 2008. Effects of NaCl on growth, ion accumulation, protein, proline contents and antioxidant enzymes activity in callus cultures of Jatropha curcas. Biologia 63: 378-382. [ Links ]
Lee, W. & L. Chan. 2004. Establishment of Orthosiphon stamineus Cell Suspension Culture for Cell Growth. Plant Cell Tiss. Organ 78: 101-106. [ Links ]
Lin, J., F. Yan, L. Tang & F. Chen. 2003. Antitumor effects of curcin from seeds of Jatropha curcas. Acta Pharmacol. Sin. 24: 241-246. [ Links ]
Luo, M.J., W.X. Liu, X.Y. Yang, Y. Xu, F. Yan, P. Huang & F. Chen. 2007. Cloning, Expression, and Antitumor Activity of Recombinant Protein of Curcin. Russ. J. Plant Physiol. 54: 202-206. [ Links ]
Martin, G. & A. Mayeux. 1984. Réfléxions sur les cultures olagineuses, energétiques II.-Le Pourghere (Jatropha curcas L.): un carburant possible. Olagineux 39: 283-287. [ Links ]
Mazarei, M., H. Al-Ahmada, M.R. Rudis, B.L. Joyce & C.N. Jr. Stewart. 2011. Switchgrass (Panicum virgatum L.) cell suspension cultures: Establishment, characterization, and application. Article in press. Plant Science. [ Links ]
Misra, P., N. Gupta, D. Toppo, Pande, V., Mishra, M. & Tuli, R. 2010. Establishment of long-term proliferating shoot cultures of elite Jatropha curcas L by controlling endophytic bacterial contamination. Plant Cell Tiss. Organ 100: 189-197. [ Links ]
Morris, S., C.W. Sigel, M.J. Matz, J.A. Saenz, R.C. Haltiwanger & R.F. Bryan. 1970. Jatrophone, a Novel Macrocyclic Diterpenoid Tumor Inhibitor from Jatropha gossypiifolia. J. Am. Chem. Soc. 92: 14. [ Links ]
Mulabagal, V. &. H. S. Tsay. 2004. Plant Cell Cultures An Alternative and Efficient Source for the Production of Biologically Important Secondary Metabolites. Int. J. Appl. Sci. Eng. 2: 29-48. [ Links ]
Murashige, T. & F. Skoog. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 15: 473-479. [ Links ]
Mythili, P., N. Seetharama & V. Reddy. 1999. Plant regeneration from embryogenic cell suspension cultures in wild species of sorghum (Sorghum dimidiatum Stapf). Plant Cell Rep. 18: 424-428. [ Links ]
Nayak, B.S. & K.N. Patel. 2010. Pharmacognostic studies of the Jatropha curcas leaves. Int. J. Pharm.Tech. Res. 2: 140-143. [ Links ]
Ngara, R., J. Rees & B.K. Ndimba. 2008. Establishment of sorghum cell suspension culture system for proteomics studies. Afr. J. Biotechnol. 7: 744-749. [ Links ]
Osoniyi, O. & F. Onajobi. 2003. Coagulant and anticoagulant activities in Jatropha curcas latex. J. Ethnopharmacol. 89: 101-105. [ Links ]
Rodríguez-Monroy, M. 2007. Técnicas para el análisis de muestras y determinación de variables de crecimiento y cálculo de parámetros cinéticos. Manual de prácticas del curso Aspectos Teórico Prácticos del cultivo de células vegetales en biorreactores. Instituto Politécnico Nacional, CEPROBI y la Sociedad Mexicana de Biotecnología y Bioingeniería A. C., Morelos, México, 14-28. [ Links ]
Schmeda-Hirschmann, G., I. Razmilic, M. Sauvain, C. Moretti, V. Muñoz, E. Ruiz, E. Balanza & A. Fournet. 1996. Antiprotozoal activity of Jatrogrossidione from Jatropha grossidentata and Jatrophone from Jatropha isabellii. Phytother. Res. 10: 375-378. [ Links ]
Sharma, S., N. Kumar & M. Reddy. 2011. Regeneration in Jatropha curcas: Factors affecting the efficiency of in vitro regeneration. Ind. Crop. Prod. 34: 943-951. [ Links ]
Siang, T., S. Soong & A. Yien. 2012. Plant regeneration studies of Jatropha curcas using induced embryogenic callus from cotyledon explants. Afr. J. Biotechnol. 11: 8022-8031. [ Links ]
Soomro, R. & R.A. Memon. 2007. Establishment of callus and suspension culture in Jatropha curcas. Pakistan J. Bot. 39: 2431-2441. [ Links ]
Staubmann, R., G. Foidl, N. Foidl, G. Gübitz, R. Lafferty, V. Valencia & W. Steiner. 1997. Biogas Production from Jatropha curcas Press-Cake. Appl. Biochem. Biotechnol. 63: 457-467. [ Links ]
Sujatha, M. & D. Mukta. 1993. Rapid plant regeneration from various explants of Jatropha integerrima. Plant Cell Tiss. Organ 35: 293-296. [ Links ]
Sujatha, M., H. Makkar & K. Becker. 2005. Shoot bud proliferation from axilary nodes and leaf sections of non-toxic Jatropha curcas L. Plant Growth Regul. 47: 83-90. [ Links ]
Sujatha, M. & N. Mukta. 1996. Morphogenesis and plant regeneration from tissue cultures of Jatropha curcas. Plant Cell Tiss. Organ 44: 135-141. [ Links ]
Torabi, F., A. Majad & A. Ehsanpour. 2008. Plant regeneration from cell suspension culture of potato (Solanum tuberosum L.). Pakistan J. Biol. Sci. 11: 778-782. [ Links ]
Varisai, S., C. Wang, M. Thiruvengadam & N. Jayabalan. 2004. In vitro plant regeneration via somatic embryogenesis through cell suspension cultures of horsegram [Macrotyloma uniflorum (lam.) verdc.]. In Vitro Cellular & Developmental Biology – Plant 40: 284-289. [ Links ]
Wai-Leng, L. & Ch. Lai-Keng. 2004. Establishment of Orthosiphon stamineus cell suspensión culture for cell growth. Plant Cell Tiss. Organ 78: 101-106. [ Links ]
Wei, Q., Y. Liao, Y. Chen, S.H. Wang, Y. Xu, L. Tang & F. Chen. 2005. Isolation, characterisation and antifungal activity of β-1, 3-glucanase from seeds of Jatropha curcas. South Afr. J. Botany 71: 95-9. [ Links ]
Weida, L., W. Qim, T. Lin, Y. Fang & C. Fang. 2003. Induction of callus from Jatropha curcas and its rapid propagation. Chin. J. Appl. Environ. Biol. 9: 127-130. [ Links ]
Zhang, Y.H., J.J. Zhong & J.T. Yu. 1996. Enhancement of ginseng saponin production in suspension cultures of Panax notoginseng: manipulation of medium sucrose. J. Biotechnol. 51: 49-56. [ Links ]
Laura Yesenia Solís-Ramos. Laboratorio de Biotecnología y Transformación Genética de Plantas, Escuela de Biología, Universidad de Costa Rica; laura.solisramos@ucr.ac.cr.
Laura Miranda Carballo. Laboratorio de Biotecnología y Transformación Genética de Plantas, Escuela de Biología, Universidad de Costa Rica. Actual address: Centro de Investigaciones Agronómicas (CIA), Escuela de Agronomía, Universidad de Costa Rica; lmirandacar@gmail.com.
Marta Valdez-Melara. Laboratorio de Biotecnología y Transformación Genética de Plantas, Escuela de Biología, Universidad de Costa Rica; marta.valdez@ucr.ac.cr.
1. Laboratorio de Biotecnología y Transformación Genética de Plantas, Escuela de Biología, Universidad de Costa Rica; laura.solisramos@ucr.ac.cr, marta.valdez@ucr.ac.cr. Corresponding author
2. Actual address: Centro de Investigaciones Agronómicas (CIA), Escuela de Agronomía, Universidad de Costa Rica; lmirandacar@gmail.com
Received 28-VIII-2012. Corrected 10-I-2013. Accepted 13-II-2013


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}